
Sustainable Agronomic Valorization of Unsulfured Molasses and Defatted Soybean Meal as an Optimized Formulation of Bio-Organic Fertilizer Enriched with High Cell Density P-Solubilizing Bacteria

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Characterization of Molasses and DSM
2.2. Test Bacteria and Fermentation Condition
2.3. C and N Sources Supplementation
2.4. Cell Density Enumeration
2.5. Two-Level Factorial Design (TFLD)
2.6. Central Composite Design (CCD) Using Response Surface Methodology (RSM)
2.7. Post-Analysis of the Optimized Model
2.8. P Solubilization Activity of E. hormaechei 40a in Different Media
2.9. Shelf-Life Determination
2.10. Statistical Analysis
3. Results and Discussion
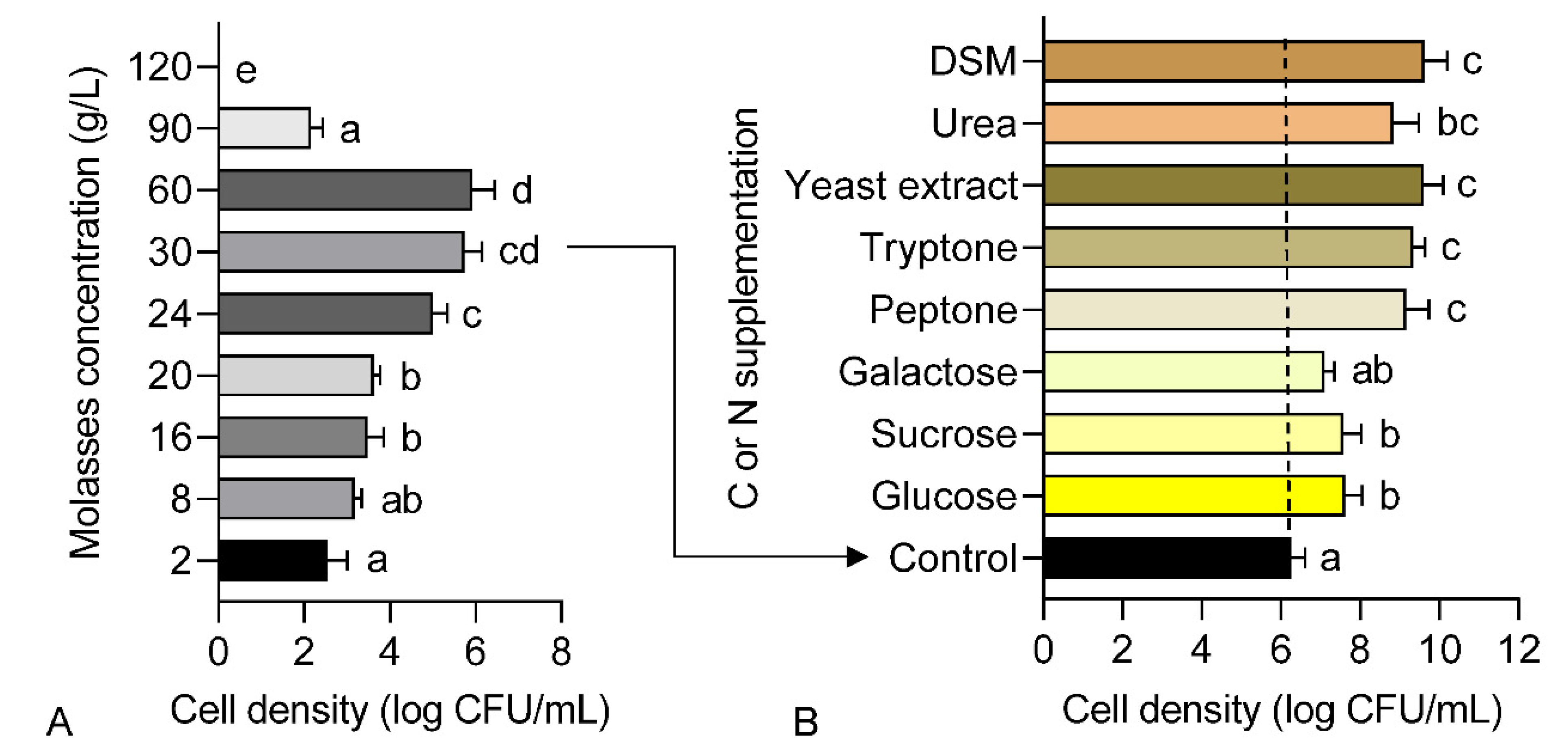
3.1. Effects of C and N Substrates Supplementation
3.2. Screening for Significant Fermentation Factors
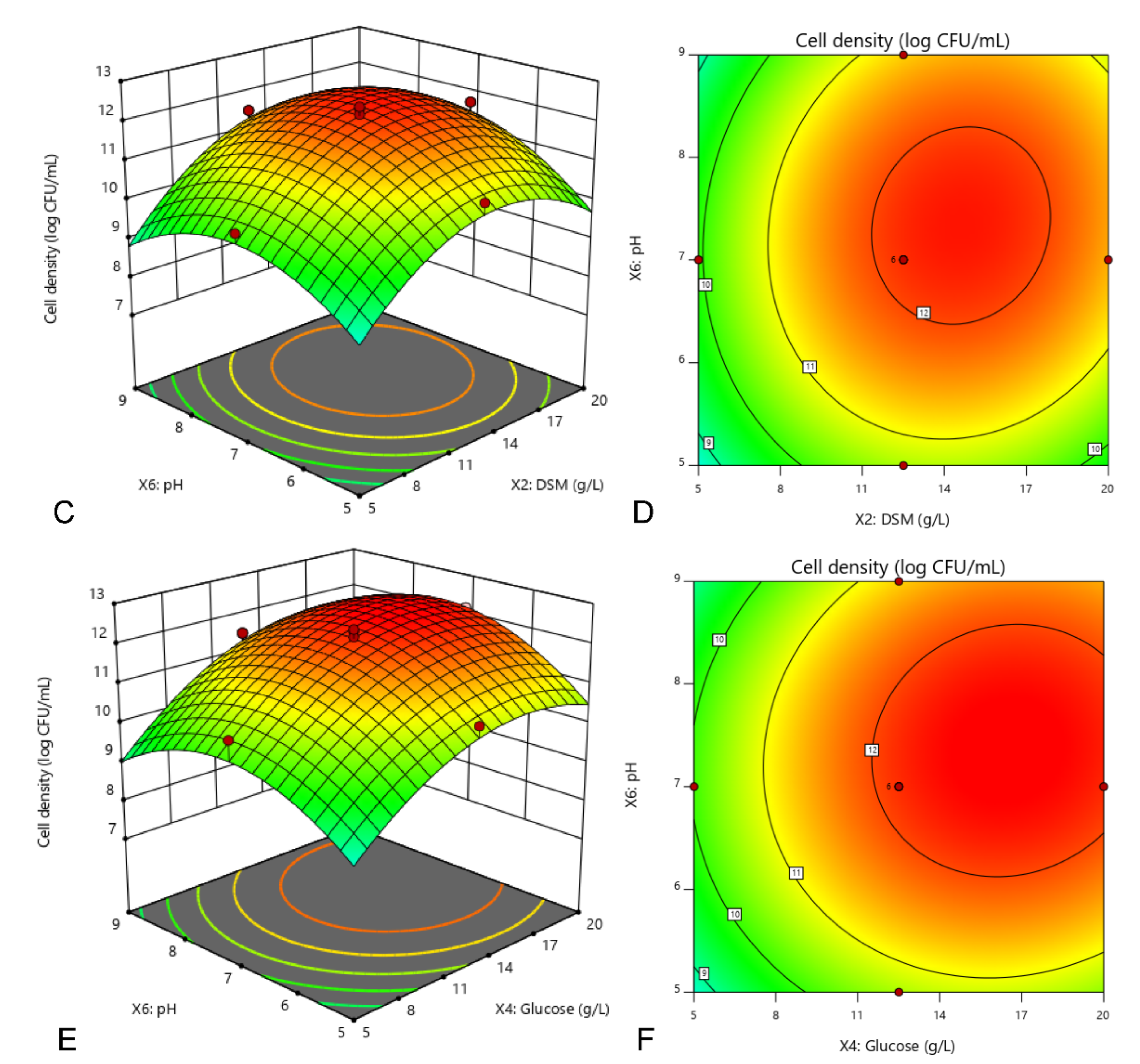
3.3. Optimization of Culture Condition
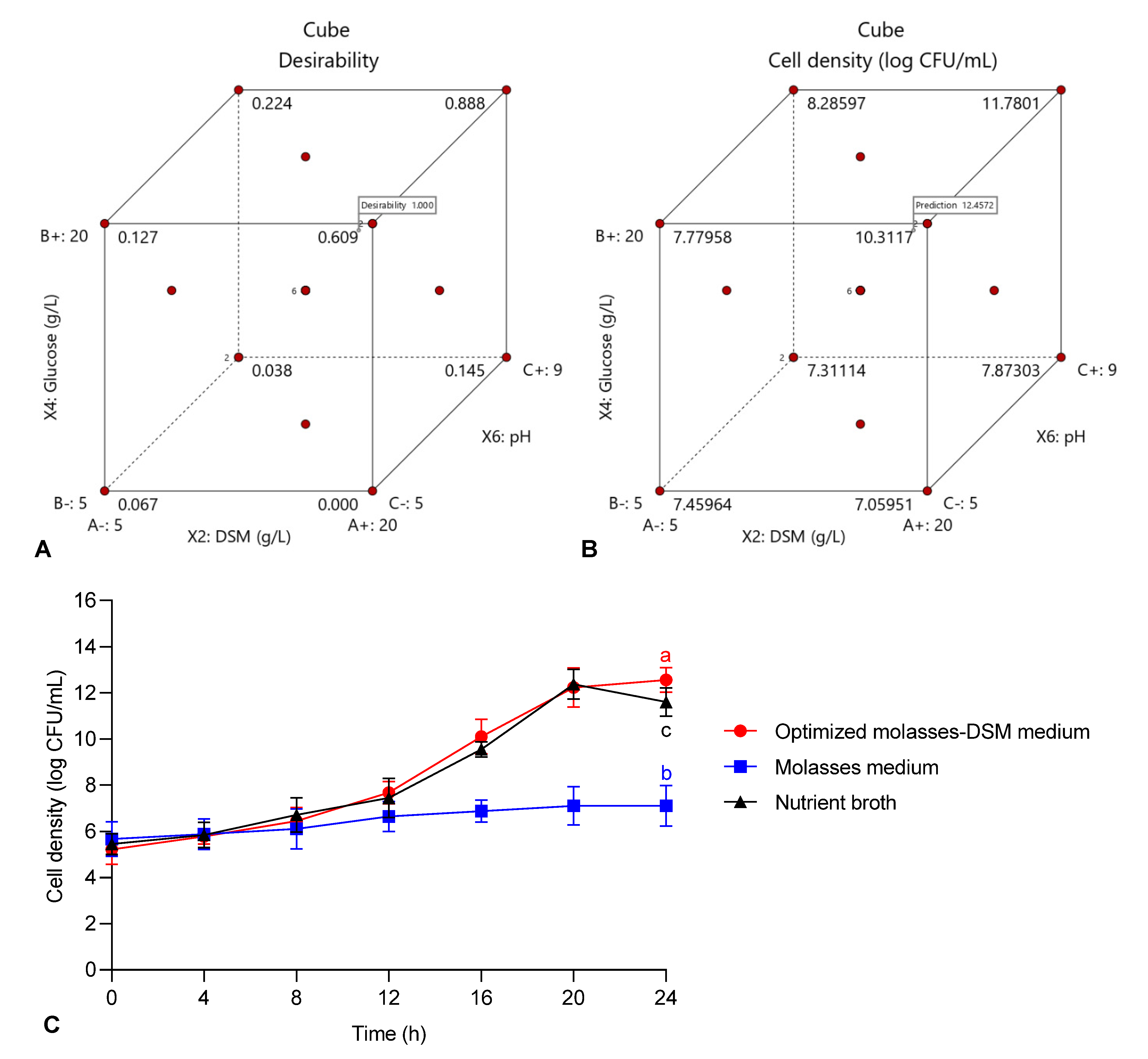
3.4. Validation of the Quadratic Optimization Model
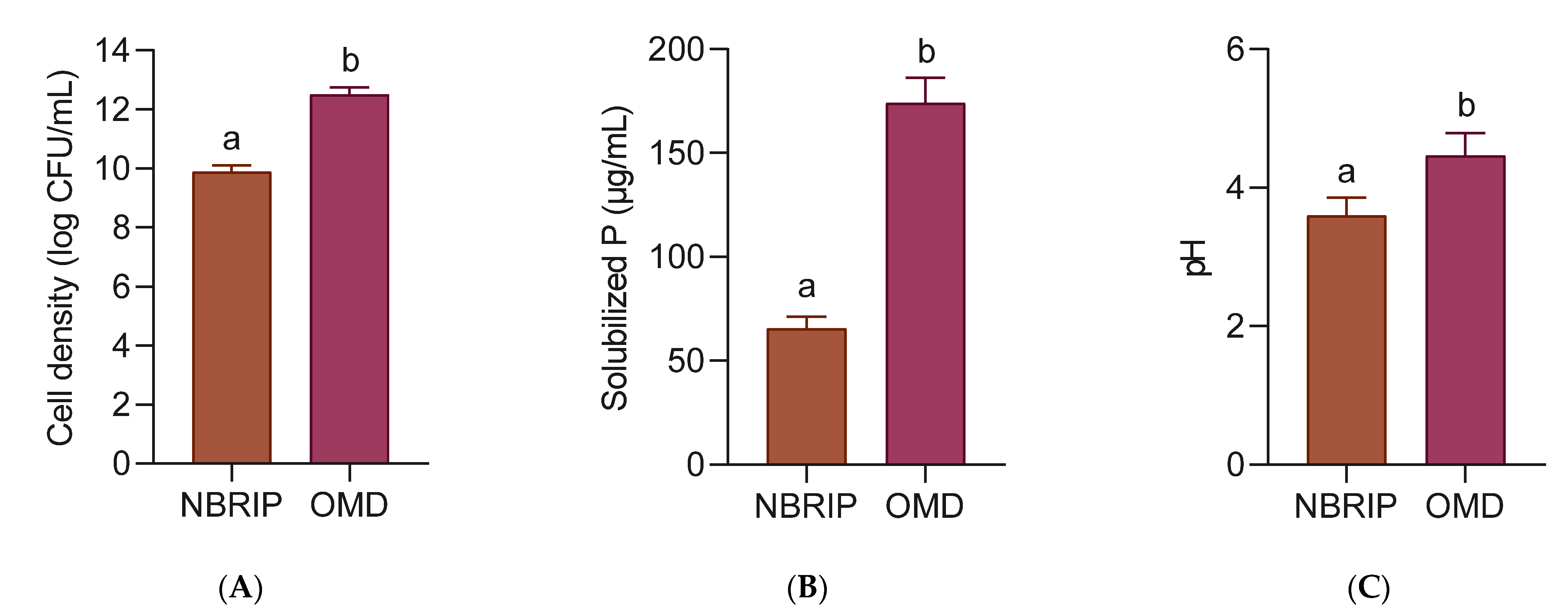
3.5. P Solubilization of CIRP between Standard NBRIP and Optimized Medium
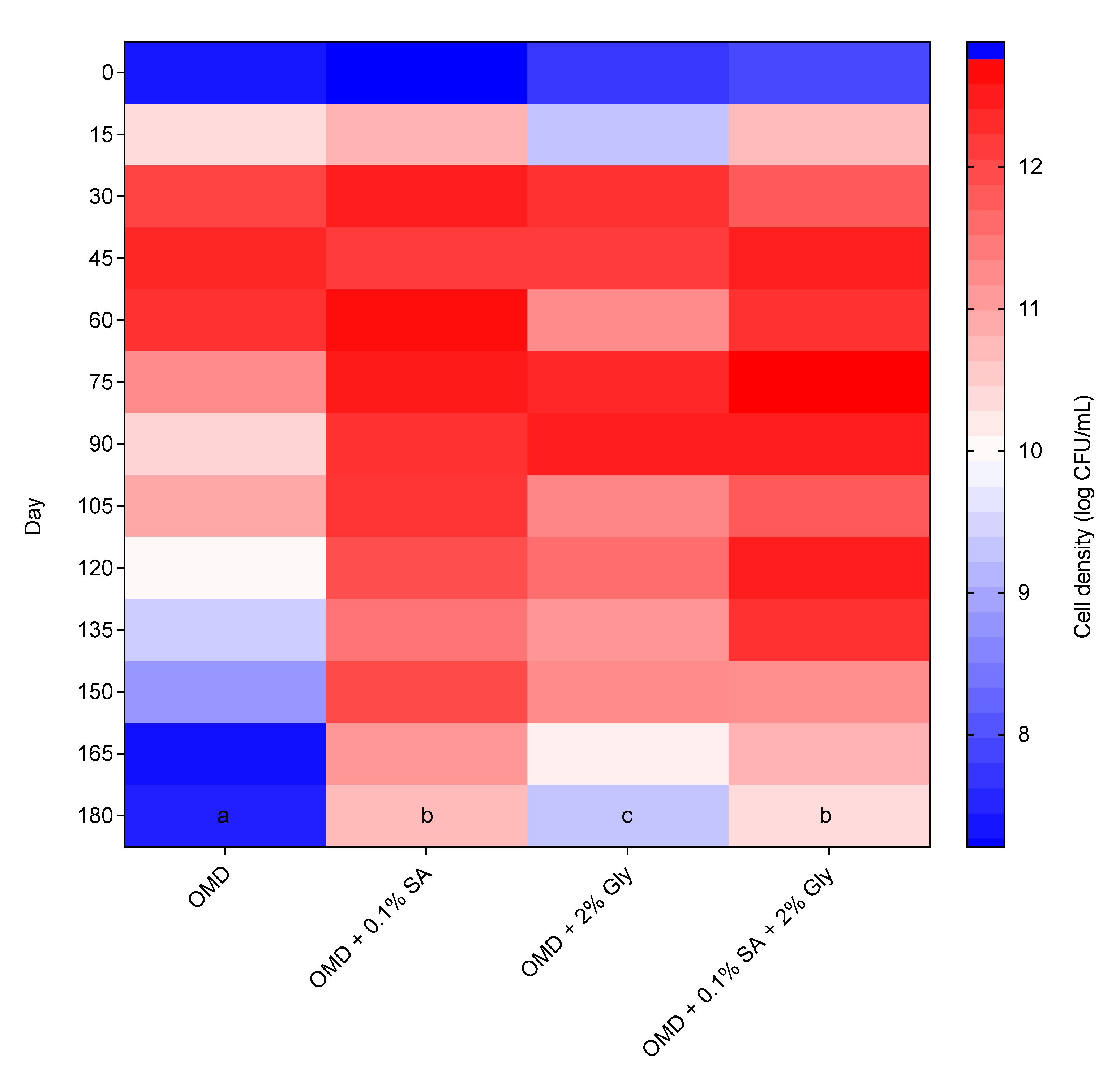
3.6. Effects of Cell Protectants on the Shelf-Life of Biofertilizer
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Roslan, M.A.M.; Zulkifli, N.N.; Sobri, Z.M.; Zuan, A.T.K.; Cheak, S.C.; Abdul Rahman, N.A. Seed biopriming with P- and K-solubilizing Enterobacter hormaechei sp. improves the early vegetative growth and the P and K uptake of okra (Abelmoschus esculentus) seedling. PLoS ONE 2020, 15, e0232860. [Google Scholar] [CrossRef] [PubMed]
- O’Callaghan, M. Microbial inoculation of seed for improved crop performance: Issues and opportunities. Appl. Microbiol. Biotechnol. 2016, 100, 5729–5746. [Google Scholar] [CrossRef] [PubMed]
- Stephens, J.H.G.; Rask, H.M. Inoculant production and formulation. Field Crop. Res. 2000, 65, 249–258. [Google Scholar] [CrossRef]
- Arora, N.K.; Khare, E.; Maheshwari, D.K. Plant Growth Promoting Rhizobacteria: Constraints in Bioformulation, Commercialization, and Future Strategies. In Plant Growth Promoting Rhizobacteria; Maheshwari, D.K., Ed.; Springer: Berlin/Heidelberg, Germany, 2010; Volume 18, pp. 97–116. Available online: http://link.springer.com/10.1007/978-3-642-13612-2 (accessed on 1 April 2021).
- Berninger, T.; González López, Ó.; Bejarano, A.; Preininger, C.; Sessitsch, A. Maintenance and assessment of cell viability in formulation of non-sporulating bacterial inoculants. Microb. Biotechnol. 2018, 11, 277–301. [Google Scholar] [CrossRef] [Green Version]
- Sahu, P.K.; Brahmaprakash, G.P. Formulations of biofertilizers—Approaches and advances. In Microbial Inoculants in Sustainable Agricultural Productivity; Singh, D.P., Singh, H.B., Prabha, R., Eds.; Springer: New Delhi, India, 2016; pp. 179–198. Available online: http://link.springer.com/10.1007/978-81-322-2644-4 (accessed on 1 April 2021).
- Garcha, S.; Kansal, R.; Gosal, S.K. Molasses growth medium for production of Rhizobium sp. Based biofertilizer. Indian J. Biochem. Biophys. 2019, 56, 378–383. [Google Scholar]
- Wang, H.Y.; Shen, L.I.U.; Zhai, L.M.; Zhang, J.Z.; Ren, T.Z.; Fan, B.Q.; Liu, H. Preparation and utilization of phosphate biofertilizers using agricultural waste. J. Integr Agric. 2015, 14, 158–167. [Google Scholar] [CrossRef]
- Singh, D.P.; Prabha, R.; Renu, S.; Sahu, P.K.; Singh, V. Agrowaste bioconversion and microbial fortification have prospects for soil health, crop productivity, and eco-enterprising. Int. J. Recycl. Org. Waste Agric. 2019, 8, 457–472. [Google Scholar] [CrossRef] [Green Version]
- Ravindran, R.; Jaiswal, A.K. Exploitation of food industry waste for high-value products. Trends Biotechnol. 2016, 34, 58–69. [Google Scholar] [CrossRef] [Green Version]
- Unsoed, P.; AS, Y.; Sumadi, S.; Simarmata, T. Viability of Trichoderma harzianum grown on different carrier formulation. KnE Life Sci. 2017, 2, 95. [Google Scholar] [CrossRef] [Green Version]
- Cajamarca, S.M.N.; Martins, D.; da Silva, J.; Fontenelle, M.R.; Guedes, Í.M.R.; de Figueiredo, C.C.; Pacheco Lima, C.E. Heterogeneity in the chemical composition of biofertilizers, potential agronomic use, and heavy metal contents of different agro-industrial wastes. Sustainability 2019, 11, 7. [Google Scholar] [CrossRef] [Green Version]
- Shah, Z.H.; Rehman, H.M.; Akhtar, T.; Alsamadany, H.; Hamooh, B.T.; Mujtaba, T.; Daur, I.; Al Zahrani, Y.; Alzahrani, H.A.; Ali, S.; et al. Humic substances: Determining potential molecular regulatory processes in plants. Front. Plant. Sci. 2018, 9, 263. [Google Scholar] [CrossRef] [Green Version]
- Vu, V.H.; Kim, K. High-cell-density fed-batch culture of Saccharomyces cerevisiae KV-25 using molasses and corn steep liquor. J. Microbiol. Biotechnol. 2009, 19, 1603–1611. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Zhou, J.; Wang, L.; Ma, Z.; Zhao, G.; Ge, Z.; Zhu, H.; Qiao, J. Improving nitrogen source utilization from defatted soybean meal for nisin production by enhancing proteolytic function of Lactococcus lactis F44. Sci Rep. 2017, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.; Kim, W.; Hwang, I.K. Optimization of the extraction and purification of oligosaccharides from defatted soybean meal. Int. J. Food Sci. Technol. 2003, 38, 337–342. [Google Scholar] [CrossRef]
- Zhang, K.; Yu, C.; Yang, S.T. Effects of soybean meal hydrolysate as the nitrogen source on seed culture morphology and fumaric acid production by Rhizopus oryzae. Process. Biochem. 2015, 50, 173–179. [Google Scholar] [CrossRef]
- Su, L.W.; Cheng, Y.H.; Hsiao, F.S.H.; Han, J.C.; Yu, Y.H. Optimization of mixed solid-state fermentation of soybean meal by Lactobacillus species and Clostridium butyricum. Pol. J. Microbiol. 2018, 67, 297–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biradar, B.J.P.; Santhosh, G.P. Cell protectants, adjuvants, surfactant and preservative and their role in increasing the shelf life of liquid inoculant formulations of Pseudomonas fluorescens. Int. J. Pure Appl. Biosci. 2018, 6, 116–122. [Google Scholar] [CrossRef]
- Sivakumar, G.K.K. Enhancing the shelf life of Azotobacter and Azospirillum bioinoculants by development of liquid formulations. Int. Arch. Appl. Sci. Technol. 2019, 10, 132–136. [Google Scholar]
- Lobo, C.B.; Juárez Tomás, M.S.; Viruel, E.; Ferrero, M.A.; Lucca, M.E. Development of low-cost formulations of plant growth-promoting bacteria to be used as inoculants in beneficial agricultural technologies. Microbiol. Res. 2019, 219, 12–25. [Google Scholar] [CrossRef]
- Thirumal, G.; Reddy, R.S.; Triveni, S.; Damodarachari, K.; Bhavya, K. Evaluate the shelf life of Rhizobium carrier based biofertilizer stored at different temperatures at different intervals. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 753–759. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.K.; Lur, H.S.; Lo, K.J.; Cheng, K.C.; Chuang, C.C.; Tang, S.J.; Yang, Z.W.; Liu, C.T. Evaluation of the effects of different liquid inoculant formulations on the survival and plant-growth-promoting efficiency of Rhodopseudomonas palustris strain PS3. Appl. Microbiol. Biotechnol. 2016, 100, 7977–7987. [Google Scholar] [CrossRef] [PubMed]
- Shu, G.; Lei, Z.; Chen, H.; Wang, P.; Huang, D. Screening of medium compounds using a two-level factorial design for Saccharomyces boulardii. Sci. Study Res. 2016, 17, 45–54. [Google Scholar]
- Box, G.E.P.; Hunter, J.S.; Hunter, W.G. Statistics for Experimenters: Design, Innovation, and Discovery; Wiley: New York, NY, USA, 2005; p. 672. [Google Scholar]
- Nautiyal, C.S. An efficient microbiological growth medium for screening phosphate solubilizing microorganisms. FEMS Microbiol. Lett. 1999, 170, 265–270. Available online: https://academic.oup.com/femsle/article-lookup/doi/10.1111/j.1574-6968.1999.tb13383.x (accessed on 1 April 2021). [CrossRef] [PubMed]
- Gupta, A.P.; Neue, H.U.; Singh, V.P. Phosphorus determination in rice plants containing variable manganese content by the Phospho-Molybdo-Vanadate (yellow) and phosphomolybdate (blue) colorimetric methods. Commun. Soil Sci. Plant. Anal. 1993, 24, 1309–1318. [Google Scholar] [CrossRef]
- Gopi, G.K.; Meenakumari, K.S.; Nysanth, N.S.; Subha, P. An optimized standard liquid carrier formulation for extended shelf-life of plant growth promoting bacteria. Rhizosphere 2019, 11, 100160. [Google Scholar] [CrossRef]
- Sugiyama, T.; Sugito, H.; Mamiya, K.; Suzuki, Y.; Ando, K.; Ohnuki, T. Hexavalent chromium reduction by an actinobacterium Flexivirga alba ST13T in the family Dermacoccaceae. J. Biosci. Bioeng. 2012, 113, 367–371. [Google Scholar] [CrossRef] [PubMed]
- Ikegami, K.; Hirose, Y.; Sakashita, H.; Maruyama, R.; Sugiyama, T. Role of polyphenol in sugarcane molasses as a nutrient for hexavalent chromium bioremediation using bacteria. Chemosphere 2020, 250. [Google Scholar] [CrossRef]
- White, P.; Johnson, L.A. Corn, Chemistry and Technology, 2nd ed.; American Association of Cereal Chemists: Saint Paul, MN, USA, 2003. [Google Scholar]
- Mohd Ridza, P.N.Y.; Ya’Acob, A.; Zainol, N.; Mortan, S.H. Application of two level factorial design to study the microbe growth inhibition by pineapple leaves juice. IOP Conf. Ser. Mater. Sci. Eng. 2020, 736, 022011. [Google Scholar] [CrossRef]
- Seshachala, U.; Tallapragada, P. Phosphate Solubilizers from the Rhizospher of Piper nigrum L. in Karnataka, India. Chil. J. Agric. Res. 2012, 72, 397–403. [Google Scholar] [CrossRef] [Green Version]
- Alori, E.T.; Glick, B.R.; Babalola, O.O. Microbial phosphorus solubilization and its potential for use in sustainable agriculture. Front. Microbiol. 2017, 8, 971. [Google Scholar] [CrossRef] [Green Version]
- Goldstein, A.H. Bioprocessing of rock phosphate ore: Essential technical considerations for the development of a successful commercial technology. In Proceedings of the 4th International Fertilizer Association Technical Conference, IFA, Paris, France, 9 May 2000; p. 220. [Google Scholar]
- Nelson, T.S.; Shieh, T.R.; Wodzinski, R.J.; Ware, J.H. The availability of phytate phosphorus in soybean meal before and after treatment with a mold phytase. Poult. Sci. 1968, 47, 1842–1848. [Google Scholar] [CrossRef]
- Herrmann, L.; Lesueur, D. Challenges of formulation and quality of biofertilizers for successful inoculation. Appl. Microbiol. Biotechnol. 2013, 97, 8859–8873. [Google Scholar] [CrossRef]
- Sandra, C.; Rebeca, B.R. Polymers selection for a liquid inoculant of Azospirillum brasilense based on the Arrhenius thermodynamic model. Afr. J. Biotechnol. 2015, 14, 2547–2553. [Google Scholar]
- Campos, D.C.; Acevedo, F.; Morales, E.; Aravena, J.; Amiard, V.; Jorquera, M.A.; Inostroza, N.G.; Rubilar, M. Microencapsulation by spray drying of nitrogen-fixing bacteria associated with lupin nodules. World J. Microbiol. Biotechnol. 2014, 30, 2371–2378. [Google Scholar] [CrossRef] [PubMed]
- Bashan, Y.; Gonzalez, L.E. Long-term survival of the plant-growth-promoting bacteria Azospirillum brasilense and Pseudomonas fluorescens in dry alginate inoculant. Appl. Microbiol. Biotechnol. 1999, 51, 262–266. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Symbols | Factors | Units | Coded Low | Coded High |
|---|---|---|---|---|
| X1 | Molasses | g/L | −1 ↔ 10 | +1 ↔ 60 |
| X2 | DSM | g/L | −1 ↔ 0 | +1 ↔ 10 |
| X3 | Yeast extract | g/L | −1 ↔ 0 | +1 ↔ 10 |
| X4 | Glucose | g/L | −1 ↔ 0 | +1 ↔ 10 |
| X5 | Sucrose | g/L | −1 ↔ 0 | +1 ↔ 10 |
| X6 | pH | −1 ↔ 4 | +1 ↔ 7 | |
| X7 | Temperature | °C | −1 ↔ 27 | +1 ↔ 37 |
| X8 | Inoculum size | % v/v | −1 ↔ 1 | +1 ↔ 4 |
| X9 | Agitation | rpm | −1 ↔ 100 | +1 ↔ 200 |
| Characteristic | Molasses | DSM |
|---|---|---|
| pH (H2O) | 4.53 ± 0.38 | 6.51 ± 0.59 |
| Moisture content (%) | 85.37 ± 1.22 | 11.33 ± 2.73 |
| Soluble sugar a (mg/g) | wmb | dmb |
| Sucrose | 138.73 ± 1.67 | 76.60 ± 1.29 |
| Glucose | 283.53 ± 5.66 | nd |
| Fructose | 122.70 ± 17.67 | nd |
| Galactose | 11.23 ± 4.97 | nd |
| Mannose | 77.23 ± 23.06 | nd |
| Elemental analysis | ||
| Macro-element (mg/g dmb) | ||
| Carbon, C b | 557.8 ± 11.8 | 419.32 ± 21.1 |
| Nitrogen, N b | 4.01 ± 0.2 | 76.17 ± 1.1 |
| Sulphur, S b | 1.2 ± 0.1 | 3.38 ± 0.2 |
| Phosphorus, P c | 16.72 ± 7.35 | 2.09 ± 0.49 |
| Potassium, K d | 4.84 ± 0.15 | 43.4 ± 4.22 |
| Calcium, Ca d | 6.24± 1.05 | 4.24 ± 1.14 |
| Magnesium, Mg d | 1.06 ± 0.91 | 2.39 ± 0.62 |
| Sodium, Na d | 0.51 ± 1.03 | 14.07 ± 0.54 |
| Micro-element (µg/g dmb) | ||
| Iron, Fe d | 42.1 ± 0.8 | 124.4 ± 5.4 |
| Boron, B d | 4.88 ± 0.19 | 366.1 ± 40.4 |
| Manganese, Mn d | 4.75 ± 0.11 | 38.2 ± 14.11 |
| Molybdenum, Mo d | 0.23 ± 0.08 | 6.2 ± 1.41 |
| Zinc, Zn d | 4.13 ± 0.18 | nd |
| Run | X1 | X2 | X3 | X4 | X5 | X6 | X7 | X8 | X9 | Cell Density (log CFU/mL) |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | − | − | − | − | − | + | + | + | + | 7.30 |
| 2 | − | + | − | + | − | + | − | + | − | 10.81 |
| 3 | + | + | − | + | − | − | + | − | + | 8.58 |
| 4 | − | − | + | + | + | + | + | + | − | 10.24 |
| 5 | + | + | − | − | + | + | − | − | + | 9.40 |
| 6 | − | − | − | + | + | − | − | + | + | 6.95 |
| 7 | + | − | − | − | + | − | + | + | + | 6.50 |
| 8 | + | + | − | − | − | + | + | + | − | 10.04 |
| 9 | + | + | − | + | + | − | − | + | − | 7.75 |
| 10 | + | + | + | − | + | − | + | − | − | 9.39 |
| 11 | + | + | + | + | − | + | − | − | − | 12.42 |
| 12 | − | + | + | − | − | + | + | − | − | 12.45 |
| 13 | − | + | − | − | − | − | − | − | + | 8.29 |
| 14 | − | + | + | − | + | + | − | + | + | 11.99 |
| 15 | + | − | − | + | − | + | − | + | + | 8.34 |
| 16 | − | + | + | + | − | − | + | + | + | 11.17 |
| 17 | + | − | + | − | − | + | + | − | + | 9.98 |
| 18 | + | + | + | − | − | − | − | + | + | 10.07 |
| 19 | − | − | + | + | − | + | − | − | + | 10.75 |
| 20 | + | − | + | + | + | − | − | − | + | 7.69 |
| 21 | + | − | + | − | + | + | − | + | − | 9.15 |
| 22 | + | − | + | + | − | − | + | + | − | 8.33 |
| 23 | + | − | − | − | − | − | − | − | − | 6.36 |
| 24 | − | + | + | + | + | − | − | − | − | 10.16 |
| 25 | − | + | − | − | + | − | + | + | − | 7.79 |
| 26 | − | − | + | − | − | − | − | + | − | 8.04 |
| 27 | − | − | + | − | + | − | + | − | + | 7.73 |
| 28 | − | − | − | − | + | + | − | − | − | 6.75 |
| 29 | − | + | − | + | + | + | + | − | + | 10.49 |
| 30 | + | + | + | + | + | + | + | + | + | 12.27 |
| 31 | + | − | − | + | + | + | + | − | − | 7.66 |
| 32 | − | − | − | + | − | − | + | − | − | 6.57 |
| Source | Sum of Squares | df | Mean Square | F-Value | p-Value | Importance (Remarks) |
|---|---|---|---|---|---|---|
| Model | 101.27 | 9 | 11.25 | 75.77 | <0.0001 | Significant |
| X1 | 0.3913 | 1 | 0.3913 | 2.63 | 0.1188 | 7 |
| X2 | 37.73 | 1 | 37.73 | 254.09 | <0.0001 | 1 (Significant) |
| X3 | 32.50 | 1 | 32.50 | 218.85 | <0.0001 | 2 (Significant) |
| X4 | 2.50 | 1 | 2.50 | 16.84 | 0.0005 | 4 (Significant) |
| X5 | 1.80 | 1 | 1.80 | 12.09 | 0.0021 | 5 (Significant) |
| X6 | 25.73 | 1 | 25.73 | 173.28 | <0.0001 | 3 (Significant) |
| X7 | 0.0752 | 1 | 0.0752 | 0.5061 | 0.4843 | 9 |
| X8 | 0.1369 | 1 | 0.1369 | 0.9222 | 0.3473 | 8 |
| X9 | 0.4060 | 1 | 0.4060 | 2.73 | 0.1124 | 6 |
| Residual | 3.27 | 22 | 0.1485 | |||
| Cor Total | 104.53 | 31 |
| Run | X2: DSM (g/L) | X4: Glucose (g/L) | X6: pH | Cell Density (log CFU/mL) | |
|---|---|---|---|---|---|
| Observed | Predicted | ||||
| 1 | 12.5 | 12.5 | 7 | 12.32 | 12.14 |
| 2 | 20 | 5 | 9 | 7.55 | 7.87 |
| 3 | 5 | 5 | 5 | 7.11 | 7.46 |
| 4 | 12.5 | 12.5 | 7 | 12.18 | 12.14 |
| 5 | 5 | 20 | 9 | 8.09 | 8.29 |
| 6 | 5 | 5 | 9 | 7.4 | 7.31 |
| 7 | 5 | 20 | 5 | 7.98 | 7.78 |
| 8 | 12.5 | 20 | 7 | 12.15 | 12.17 |
| 9 | 5 | 12.5 | 7 | 10.17 | 9.92 |
| 10 | 12.5 | 5 | 7 | 10.57 | 10.06 |
| 11 | 20 | 20 | 5 | 10.1 | 10.31 |
| 12 | 12.5 | 12.5 | 7 | 12.01 | 12.14 |
| 13 | 12.5 | 12.5 | 7 | 11.44 | 12.14 |
| 14 | 12.5 | 12.5 | 7 | 12.37 | 12.14 |
| 15 | 20 | 12.5 | 7 | 11.7 | 11.47 |
| 16 | 12.5 | 12.5 | 5 | 10.91 | 10.62 |
| 17 | 12.5 | 12.5 | 7 | 11.52 | 12.14 |
| 18 | 20 | 5 | 5 | 7.13 | 7.06 |
| 19 | 20 | 20 | 9 | 12.01 | 11.78 |
| 20 | 12.5 | 12.5 | 9 | 11.48 | 11.28 |
| Source | Sum of Squares | df | Mean Square | F-Value | p-Value | Remarks |
|---|---|---|---|---|---|---|
| Model | 72.35 | 9 | 8.04 | 42.49 | <0.0001 | significant |
| X2 | 5.98 | 1 | 5.98 | 31.62 | 0.0002 | |
| X4 | 11.17 | 1 | 11.17 | 59.02 | <0.0001 | |
| X6 | 1.09 | 1 | 1.09 | 5.75 | 0.0374 | |
| X2X4 | 4.30 | 1 | 4.30 | 22.72 | 0.0008 | |
| X2X6 | 0.46 | 1 | 0.46 | 2.45 | 0.1489 | |
| X4X6 | 0.21 | 1 | 0.21 | 1.13 | 0.3121 | |
| X22 | 5.73 | 1 | 5.73 | 30.29 | 0.0003 | |
| X42 | 2.88 | 1 | 2.88 | 15.21 | 0.0030 | |
| X62 | 3.88 | 1 | 3.88 | 20.48 | 0.0011 | |
| Residual | 1.89 | 10 | 0.19 | |||
| Lack of Fit | 1.08 | 5 | 0.22 | 1.34 | 0.3780 | not significant |
| Pure Error | 0.81 | 5 | 0.16 | |||
| Cor Total | 74.24 | 19 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roslan, M.A.M.; Sohedein, I.; Ling, P.S.; Sobri, Z.M.; Zuan, A.T.K.; Cheak, S.C.; Rahman, N.A.A. Sustainable Agronomic Valorization of Unsulfured Molasses and Defatted Soybean Meal as an Optimized Formulation of Bio-Organic Fertilizer Enriched with High Cell Density P-Solubilizing Bacteria. Agronomy 2021, 11, 996. https://doi.org/10.3390/agronomy11050996
Roslan MAM, Sohedein I, Ling PS, Sobri ZM, Zuan ATK, Cheak SC, Rahman NAA. Sustainable Agronomic Valorization of Unsulfured Molasses and Defatted Soybean Meal as an Optimized Formulation of Bio-Organic Fertilizer Enriched with High Cell Density P-Solubilizing Bacteria. Agronomy. 2021; 11(5):996. https://doi.org/10.3390/agronomy11050996
Chicago/Turabian StyleRoslan, Muhamad Aidilfitri Mohamad, Izzalan Sohedein, Puan Sheau Ling, Zulfazli M. Sobri, Ali Tan Kee Zuan, Sim Choon Cheak, and Nor Aini Abdul Rahman. 2021. "Sustainable Agronomic Valorization of Unsulfured Molasses and Defatted Soybean Meal as an Optimized Formulation of Bio-Organic Fertilizer Enriched with High Cell Density P-Solubilizing Bacteria" Agronomy 11, no. 5: 996. https://doi.org/10.3390/agronomy11050996
APA StyleRoslan, M. A. M., Sohedein, I., Ling, P. S., Sobri, Z. M., Zuan, A. T. K., Cheak, S. C., & Rahman, N. A. A. (2021). Sustainable Agronomic Valorization of Unsulfured Molasses and Defatted Soybean Meal as an Optimized Formulation of Bio-Organic Fertilizer Enriched with High Cell Density P-Solubilizing Bacteria. Agronomy, 11(5), 996. https://doi.org/10.3390/agronomy11050996