
Self-Renewal of Invasive Goldenrods (Solidago spp.) as a Result of Different Mechanical Management of Fallow



Abstract
:1. Introduction
2. Materials and Methods
2.1. Research Area
2.2. Soil–Weather Conditions
2.3. Mechanical Treatments
2.4. Phenological Development Observations
2.5. Measurements of Biomass and Biometric Parameters
2.6. Statistical Analysis
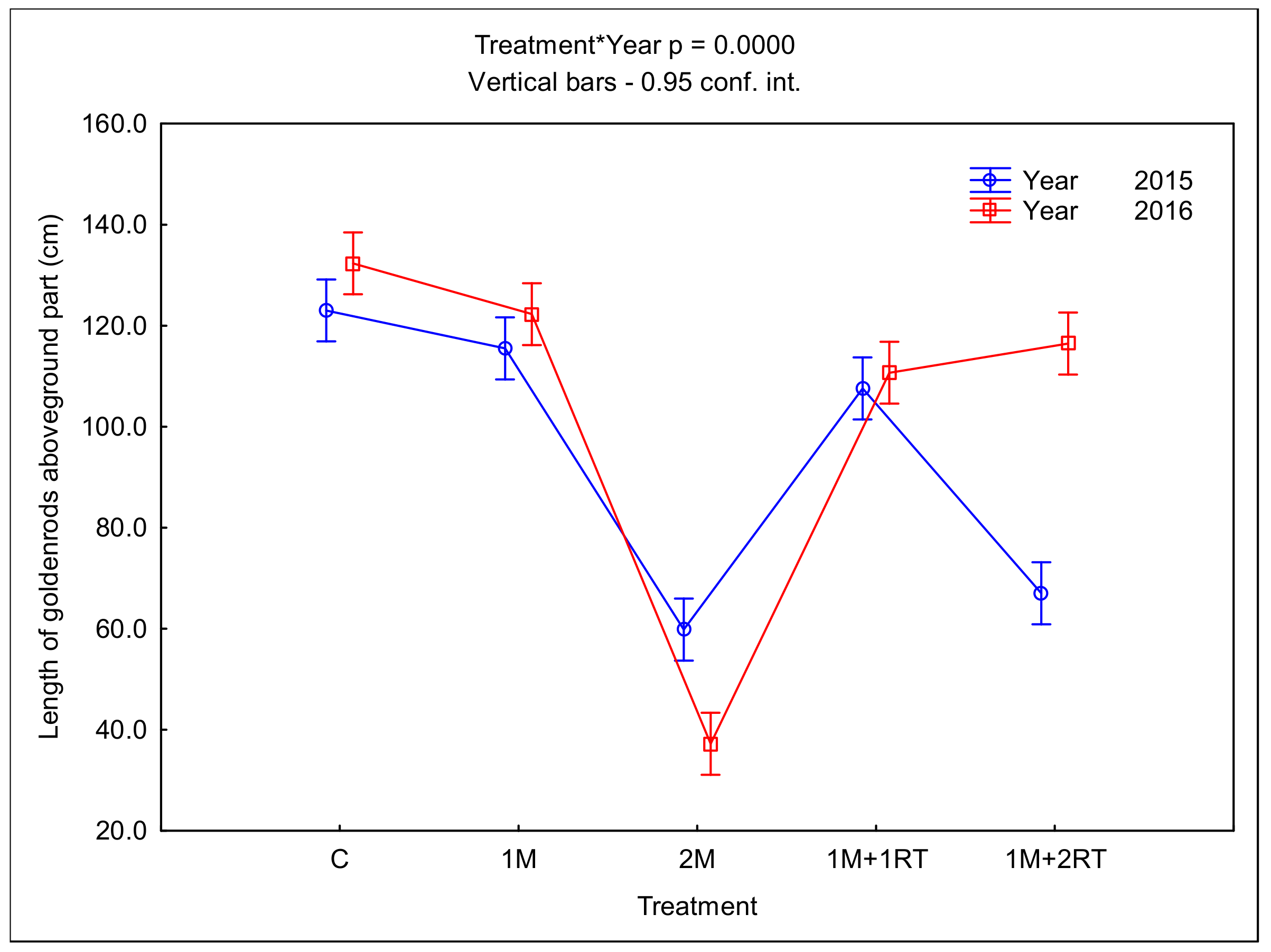
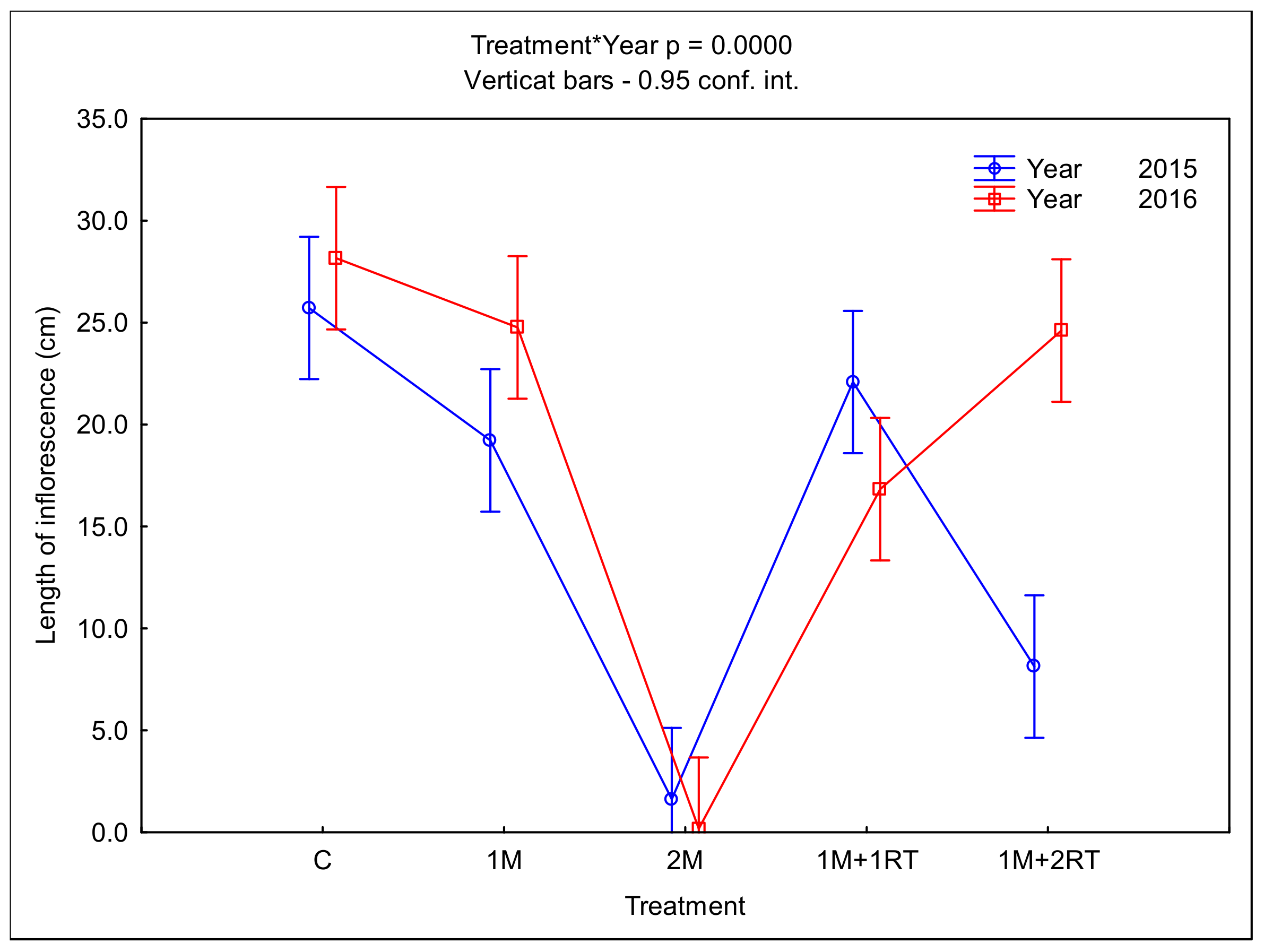
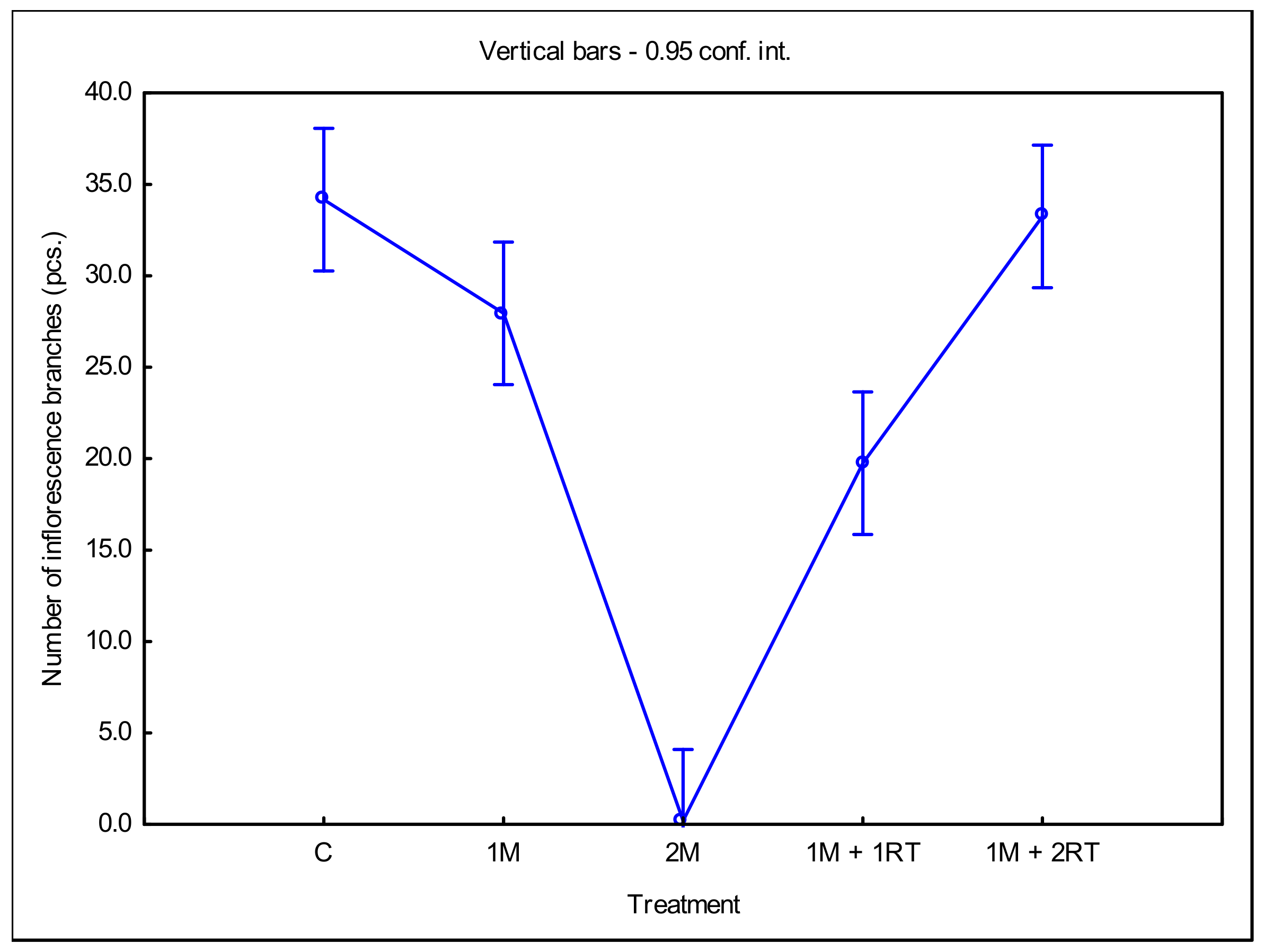
3. Results
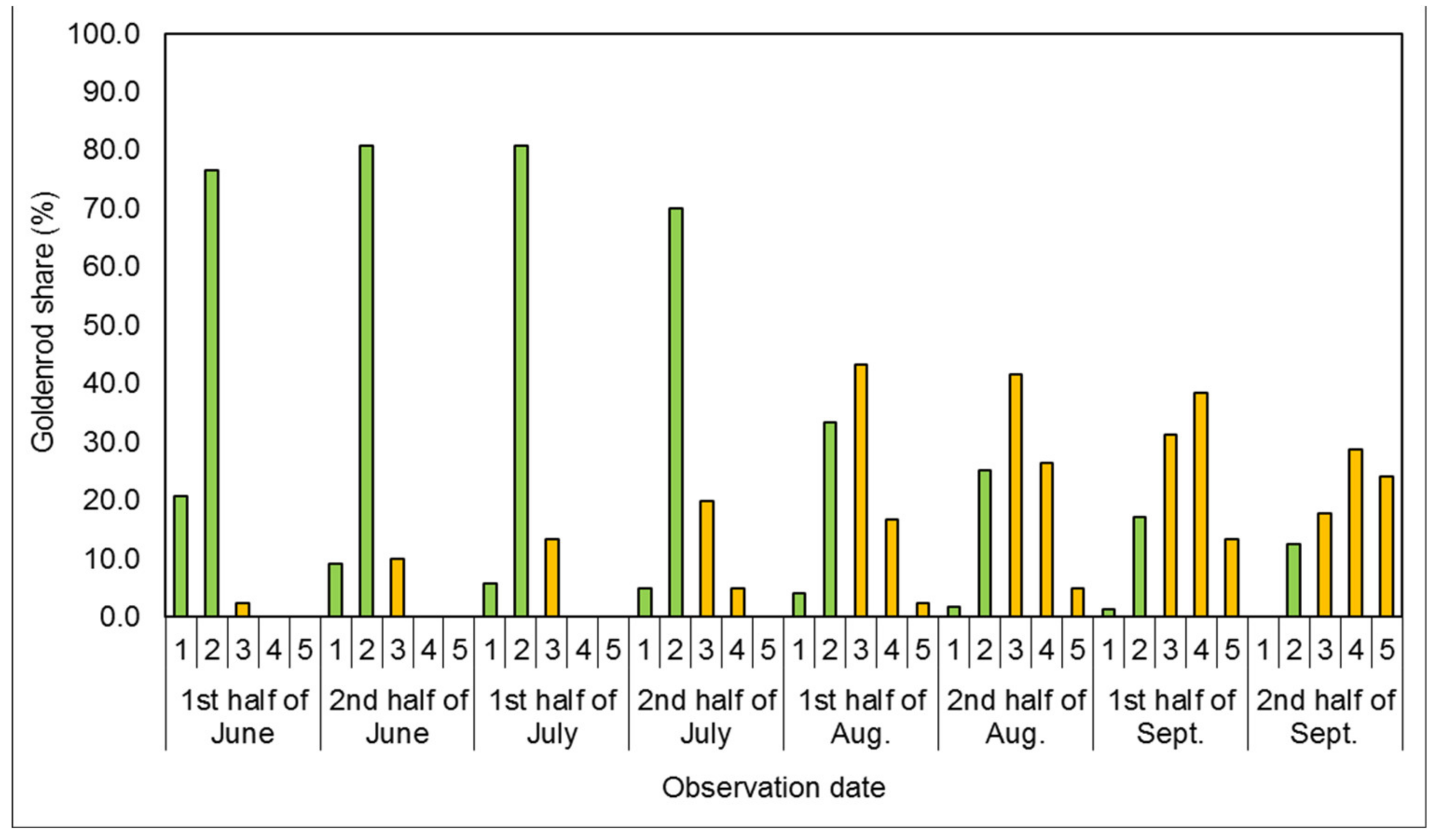
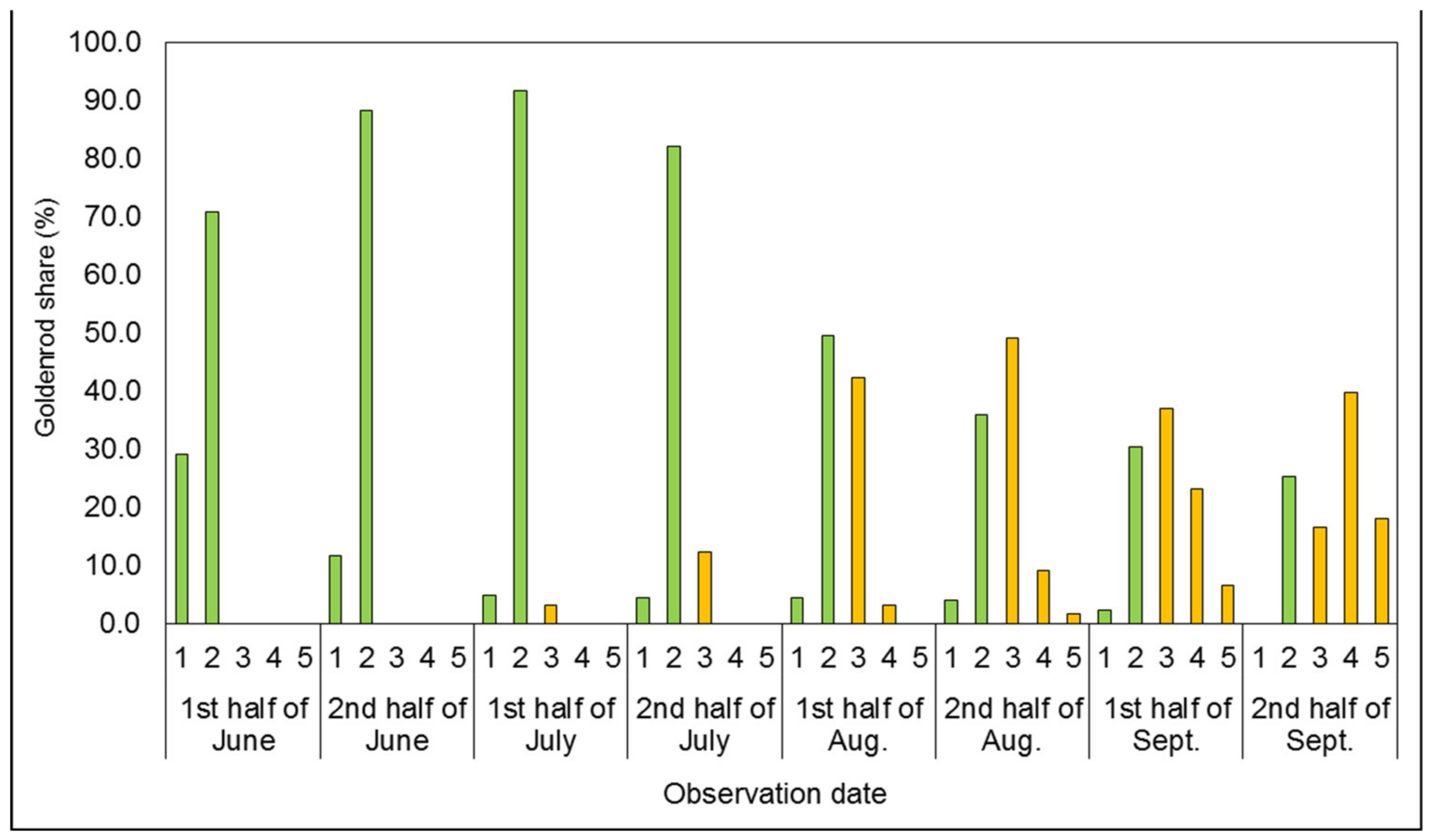
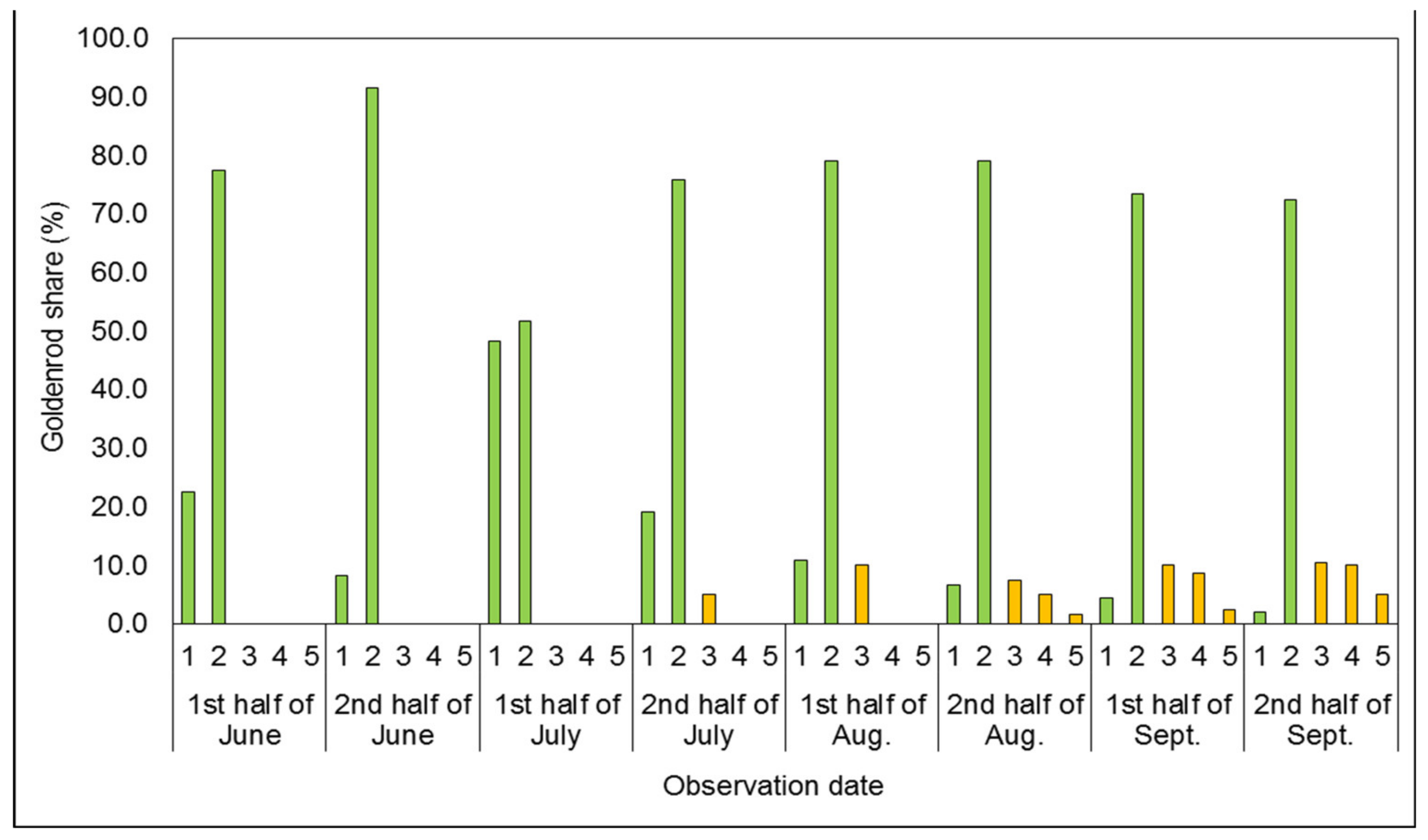
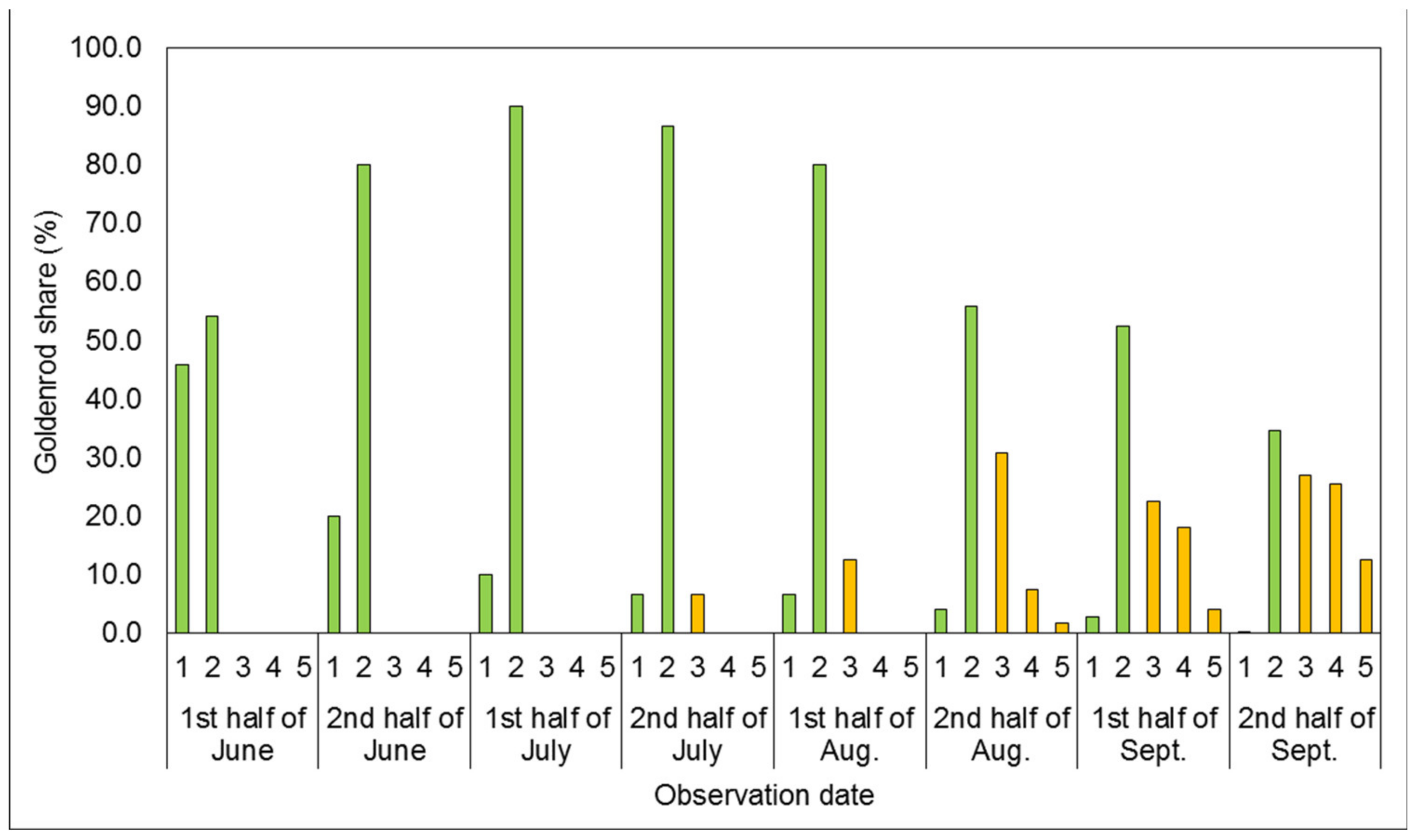
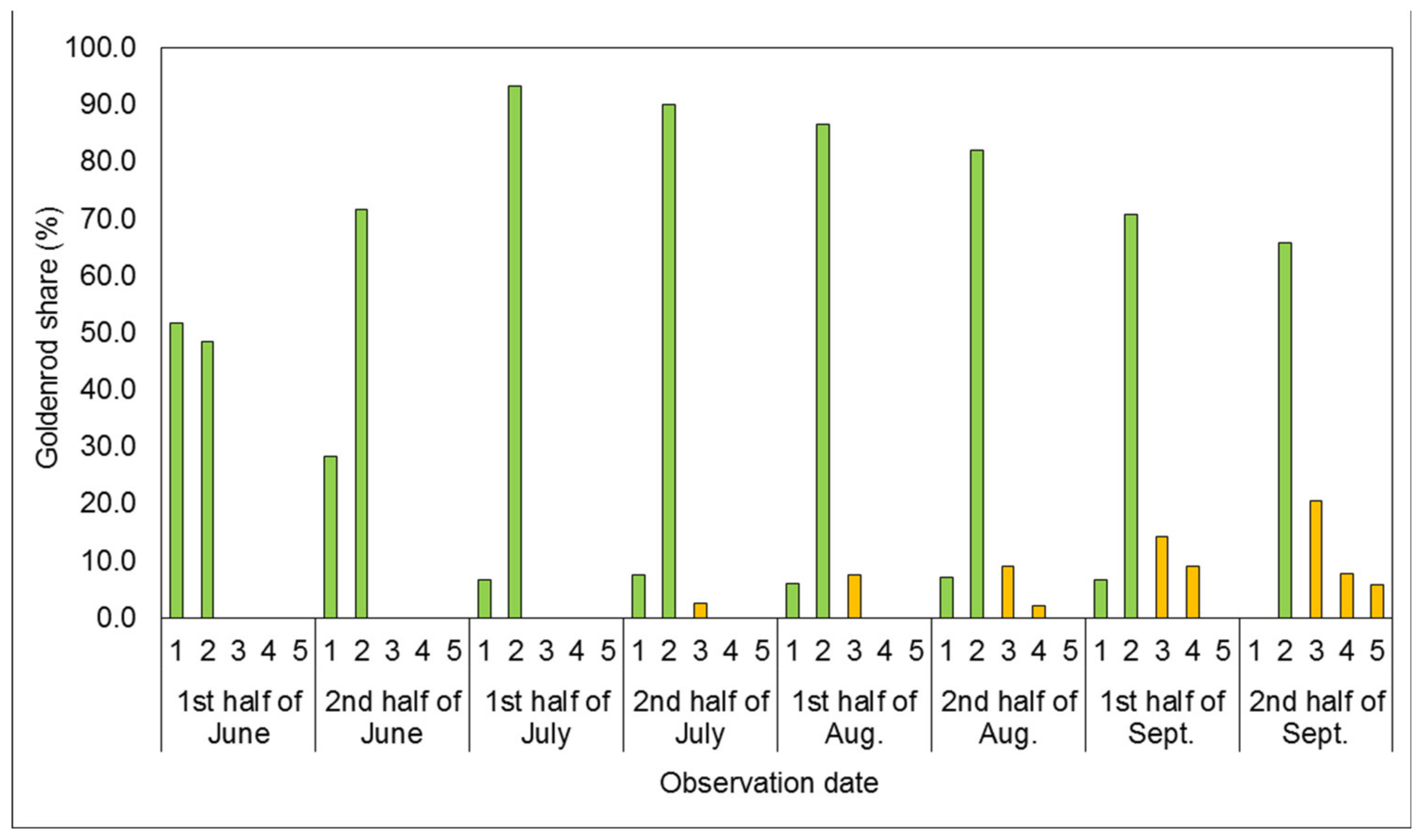
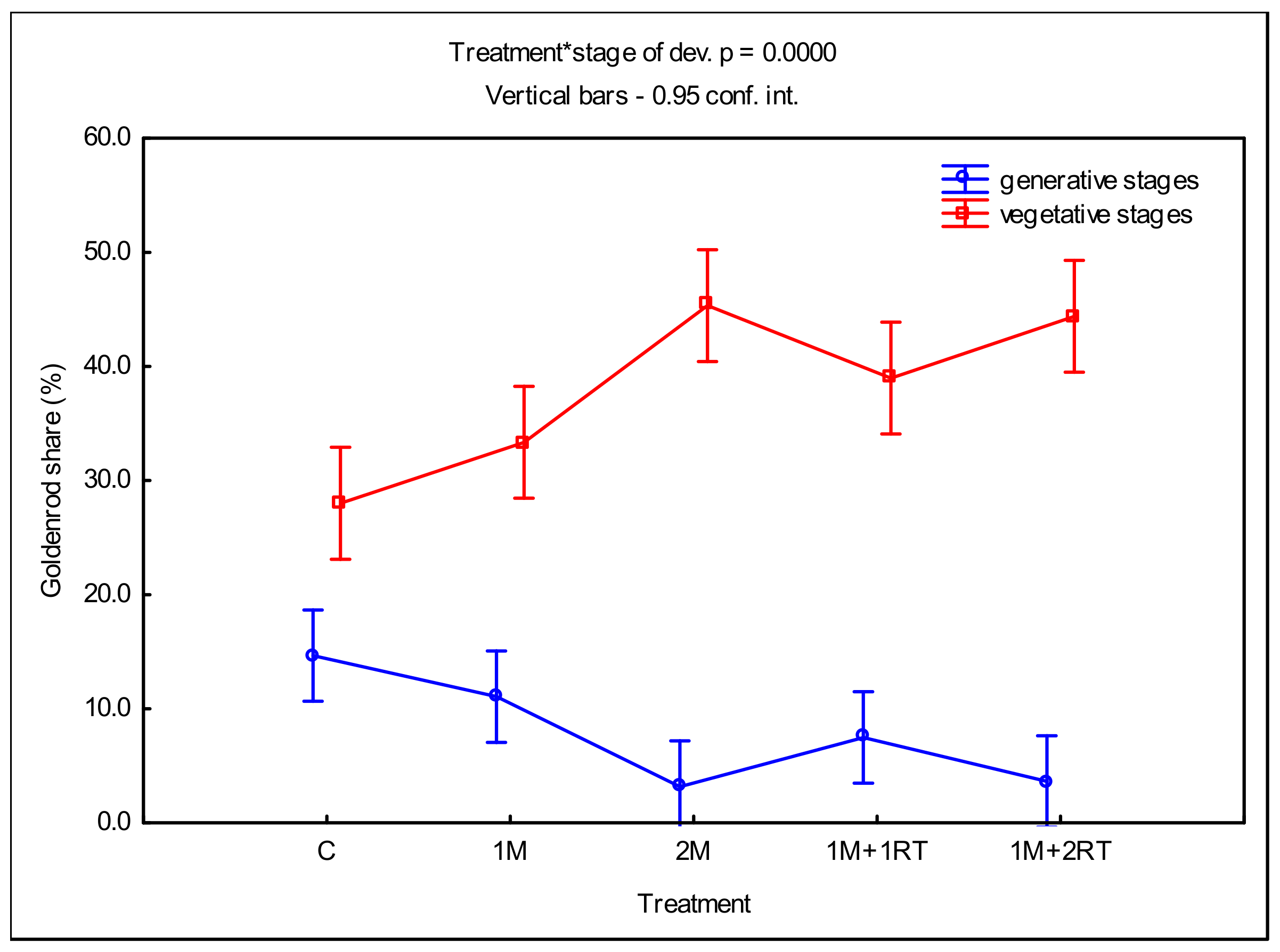
3.1. Phenological Development Observations
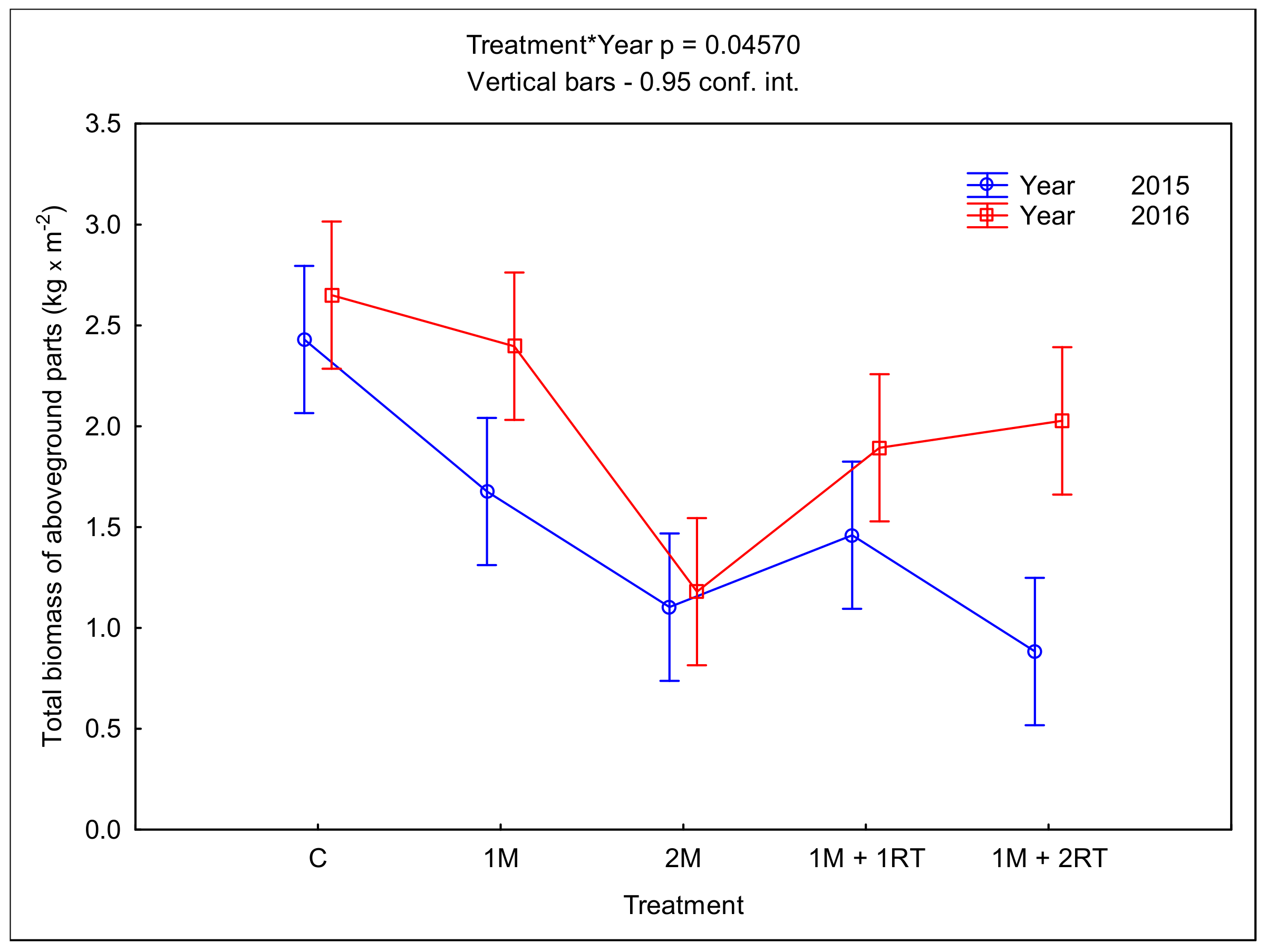
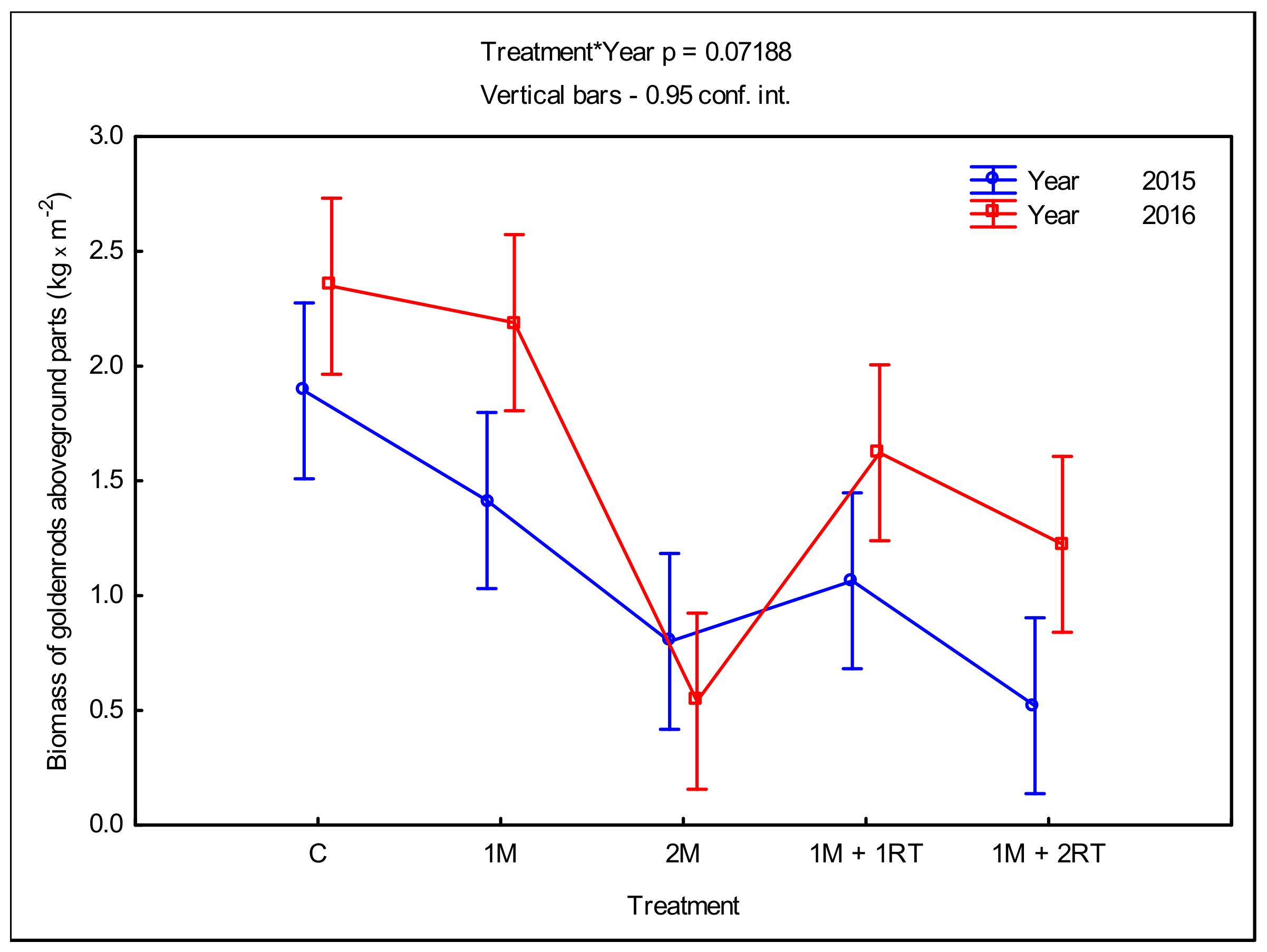
3.2. Measurements of Biomass and Biometric Parameters
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Weber, E. Current and potential ranges of three exotic goldenrods (Solidago) in Europe. Conserv. Biol. 2001, 15, 122–128. [Google Scholar] [CrossRef]
- Jakobs, G.; Weber, E.; Edwards, P.J. Introduced plants of the invasive Solidago gigantea (Asteraceae) are larger and grow denser than conspecifics in the native range. Divers. Distrib. 2004, 10, 11–19. [Google Scholar] [CrossRef]
- Tokarska-Guzik, B. The Establishment and Spread of Alien Plants Species (Kenophytes) in the Flora of Poland; University of Silesia: Katowice, Poland, 2005; p. 192. [Google Scholar]
- Tokarska-Guzik, B.; Dajdok, Z.; Zając, M.; Zając, A.; Urbisz, A.; Danielewicz, W.; Hołdyński, C. The Plants of Foreign Origin in Poland with Particular Regard to Invasive Species; Generalna Dyrekcja Ochrony Środowiska: Warsaw, Poland, 2012; p. 197. (In Polish)
- Končeková, L.; Zahradníková, E.; Pintér, E.; Halmová, D. Assessment of an impact of mechanical regulation on selected morphometric and productive parameters of invasive species Solidago canadensis population in agricultural land. Agriculture 2015, 61, 121–128. [Google Scholar] [CrossRef] [Green Version]
- Rajdus, T.; Svehlakova, H.; Plohak, P.; Stalmachova, B. Management of invasive species Solidago canadensis in Ostrava region (Czech Republic). IOP Conf. Ser. Earth Environ. Sci. 2020, 444, 012046. [Google Scholar] [CrossRef]
- Pal, R.W.; Chen, S.; Nagy, D.U.; Callaway, R.M. Impacts of Solidago gigantea on other species at home and away. Biol. Invasions 2015, 17, 3317–3325. [Google Scholar] [CrossRef]
- Nagy, D.U.; Rauschert, E.S.J.; Henn, T.; Cianfaglione, K.; Stranczinger, S.; Pal, R.W. The more we do, the less we gain? Balancing effort and efficacy in managing the Solidago gigantea invasion. Weed Res. 2020, 60, 232–240. [Google Scholar] [CrossRef] [Green Version]
- Rosef, L.; Ingebrigtsen, H.H.; Heegaard, E. Vegetative propagation of Solidago canadensis—Do fragment size and burial depth matter? Weed Res. 2020, 60, 132–141. [Google Scholar] [CrossRef] [Green Version]
- Weber, E. Invasive Plant Species of the World: A Reference Guide to Environmental Weeds; CABI Publishing: Oxon, UK, 2003; p. 581. [Google Scholar]
- Shen, G.-H.; Yao, H.-M.; Guan, L.-Q.; Qian, Z.-G.; Ao, Y.-S. Distribution and infestation of Solidago canadensis L. in Shanghai suburbs and its chemical control. Acta Agric. Shanghai 2005, 21, 1–4. [Google Scholar]
- Mei, D.; Jian-Zhong, L.; Wen-Ju, Z.C.; Bo, L. Canada goldenrod (Solidago canadensis): An invasive alien weed rapidly spreading in China. Acta Phytotax. Sin. 2006, 44, 72–85. [Google Scholar]
- Zhang, C.B.; Wang, J.; Quin, B.Y.; Li, W.H. Effects of the invader Solidago canadensis on soil properties. Appl. Soil Ecol. 2009, 43, 163–169. [Google Scholar] [CrossRef]
- Xu, Z.; Peng, H.; Feng, Z.; Abdulsalih, N. Predicting current and future invasion of Solidago canadensis: A study from China. Pol. J. Ecol. 2014, 62, 263–271. [Google Scholar] [CrossRef]
- Dong, M.; Lu, B.R.; Zhang, H.B.; Chen, J.K.; Li, B. Role of sexual reproduction in the spread of an invasive clonal plant Solidago canadensis revealed using intersimple sequence repeat markers. Plant Spec. Biol. 2006, 21, 13–18. [Google Scholar] [CrossRef]
- Weber, E. Biological flora of Central Europe: Solidago altissima L. Flora 2000, 195, 123–134. [Google Scholar] [CrossRef]
- Weber, E.; Jakobs, G. Biological flora of central Europe: Solidago gigantea Ation. Flora 2005, 200, 109–118. [Google Scholar] [CrossRef]
- Hitchmough, J.; De la Fleur, M.; Findlay, C. Establishing North American prairie vegetation in urban parks in Northern England. Part 1. Effect of sowing season, sowing rate and soil type. Landsc. Urban. Plan. 2004, 66, 75–90. [Google Scholar] [CrossRef]
- Walck, J.L.; Baskin, J.M.; Baskin, C.C. Relative competitive abilities and growth characteristics of a narrowly endemic and geographically widespread Solidago species (Asteraceae). Am. J. Bot. 1999, 86, 820–828. [Google Scholar] [CrossRef] [Green Version]
- Popiela, A.; Łysko, A.; Sotek, Z.; Ziarnek, K. Preliminary results of studies on the distribution of invasive alien vascular plant species occurring in semi-natural and natural habitats in NW Poland. Biodivers. Res. Conserv. 2015, 37, 21–35. [Google Scholar] [CrossRef] [Green Version]
- Szymura, M.; Szymura, T.H. Distribution of goldenrods (Solidago spp.) in Lower Silesia and their impact on biodiversity of invaded vegetation. Acta Botanica Silesiaca 2011, 6, 195–212. [Google Scholar] [CrossRef] [Green Version]
- Domaradzki, K.; Badowski, M. Possibility of chemical reduction of Solidago gigantea Aiton occurrence on fallow lands. Zesz. Nauk. UP Wroc. Rol. C 2012, 584, 17–24. (In Polish) [Google Scholar]
- Pużyńska, K.; Stokłosa, A.; Stupnicka-Rodzynkiewicz, E. The impact of ecological conditions on Solidago sp. occurrence. Zesz. Nauk. UP Wroc. Rol. C 2012, 584, 89–98. (In Polish) [Google Scholar]
- Domaradzki, K.; Jezierska-Domaradzka, A.; Badowski, M.; Matkowski, A. Occurrence and the possibility of late goldenrod (Solidago gigantea Aiton) control. Prog. Plant Prot. 2018, 58, 209–215. (In Polish) [Google Scholar]
- Abhilasha, D.; Quintana, N.; Vivanco, J.; Joshi, J. Do allelopathic compounds in invasive Solidago canadensis s.l. restrain the native European flora? J. Ecol. 2008, 96, 993–1001. [Google Scholar] [CrossRef]
- Fenesi, A.; Vágási, C.I.; Beldean, M.; Földesi, R.; Kolcsár, L.-P.; Shapiro, J.T.; Török, E.; Kovács-Hostyánszki, A. Solidago canadensis impacts on native plant and pollinator communities in different-aged old fields. Basic Appl. Ecol. 2015, 16, 335–346. [Google Scholar] [CrossRef] [Green Version]
- Moroń, D.; Lenda, M.; Skórka, P.; Szentgyörgyi, H.; Settele, J.; Wojciechowski, M. Wild pollinator communities are negatively affected by invasion of alien goldenrods in grassland landscapes. Biol. Conserv. 2009, 142, 1322–1332. [Google Scholar] [CrossRef]
- Skórka, P.; Lenda, M.; Tryjanowski, P. Invasive alien goldenrods negatively affect grassland bird communities in Eastern Europe. Biol. Conserv. 2010, 143, 856–861. [Google Scholar] [CrossRef]
- Tokarska-Guzik, B.; Bzdęga, K.; Nowak, T.; Urbisz, A.; Węgrzynek, B.; Dajdok, Z. Proposed List of Alien Plant Species that May Pose a Threat to the Nature of Poland and the European Union; University of Silesia: Katowice, Poland, 2015; p. 178. (In Polish) [Google Scholar]
- Bielecka, A.; Królak, E.; Biardzka, E. Habitat conditions of Canadian goldenrod in a selected region of Eastern Poland. J. Ecol. Eng. 2017, 18, 76–81. [Google Scholar] [CrossRef] [Green Version]
- Ye, X.-Q.; Yan, Y.-N.; Wu, M.; Yu, F.-H. High capacity of nutrient accumulation by invasive Solidago canadensis in a coastal grassland. Front. Plant Sci. 2019, 10, 575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozak, M.; Pudełko, R. Impact assessment of the long-term fallowed land on agricultural soils and the possibility of their return to agriculture. Agriculture 2021, 11, 148. [Google Scholar] [CrossRef]
- Chapuis-Lardy, L.; Vanderhoeven, S.; Dassonville, N.; Koutika, L.-S.; Meerts, P. Effect of the exotic invasive plant Solidago gigantea on soil phosphorus status. Biol. Fertil. Soils 2006, 42, 481–489. [Google Scholar] [CrossRef] [Green Version]
- Herr, C.; Chapuis-Lardy, L.; Dassonville, N.; Vanderhoeven, S.; Meerts, P. Seasonal effect of the exotic invasive plant Solidago gigantea on soil pH and P fractions. J. Plant Nutr. Soil Sci. 2007, 170, 729–738. [Google Scholar] [CrossRef] [Green Version]
- Szymura, M. Evaluation of ability for generative and vegetative reproduction of goldenrods occurred in Poland. Zesz. Nauk. UP Wroc. Rol. C 2012, 585, 103–112. (In Polish) [Google Scholar]
- Szymura, M.; Szymura, T.H. The dynamics of growth and flowering of invasive Solidago species. Steciana 2015, 19, 143–152. [Google Scholar] [CrossRef]
- Bing-Yao, S.; Jian-Zhong, T.; Zhi-Gang, W.; Fu-Gen, G.; Ming-De, Z. Allelopathic effects of extracts from Solidago canadensis L. against seed germination and seedling growth of some plants. J. Environ. Sci. 2006, 18, 304–309. [Google Scholar]
- Yang, R.Y.; Mei, L.X.; Tang, J.J.; Chen, X. Allelopathic effects of invasive Solidago canadensis L. on germination and growth of native Chinese plant species. Allelopath. J. 2007, 19, 241–248. [Google Scholar]
- Baličević, R.; Ravlić, M.; Živković, T. Allelopathic effect of invasive species giant goldenrod (Solidago gigantea Ait.) on crops and weeds. Herbologia 2015, 15, 19–29. [Google Scholar] [CrossRef]
- Wu, R.; Wu, B.; Cheng, H.; Wang, S.; Wie, M.; Wang, C. Drought enhanced the allelopathy of goldenrod on the seed germination and seedling growth performance of lettuce. Pol. J. Environ. Stud. 2021, 30, 423–432. [Google Scholar] [CrossRef]
- Ren, G.; He, M.; Li, G.; Anandkumar, A.; Dai, Z.; Zou, C.B.; Hu, Z.; Ran, Q.; Du, D. Effects of Solidago canadensis invasion and climate warming on soil net N mineralization. Pol. J. Environ. Stud. 2020, 29, 3285–3294. [Google Scholar] [CrossRef]
- Pokojski, W. Drought hazard in Poland and climatic water balance. Prace Stud. Geogr. 2015, 57, 103–110. (In Polish) [Google Scholar]
- Kirpluk, I.; Bomanowska, A. The occurrence of alien species in the settlement areas of the Kampinos National Park and its vicinity (Central Poland). Biodivers. Res. Conserv. 2015, 39, 79–90. [Google Scholar] [CrossRef] [Green Version]
- Świerszcz, S.; Szymura, M.; Wolski, K.; Szymura, T.H. Comparison of methods for restoring meadows invaded by Solidago species. Pol. J. Environ. Stud. 2017, 26, 1251–1258. [Google Scholar] [CrossRef]
- Brzank, M.; Piekut, K.; Dąbrowski, P.; Pawluśkiewicz, B. The succession and regression of plant species on lowland hay meadows in Poland. Pol. J. Environ. Stud. 2019, 28, 1567–1577. [Google Scholar] [CrossRef]
- Frelich, M.; Bzdęga, K. Management of invasive plant species in the valley of the River Ślepiotka in Katowice—The example of the REURIS project. Environ. Soc. Econ. Stud. 2014, 2, 26–37. [Google Scholar] [CrossRef] [Green Version]
- Nowak, A.; Kącki, Z. Species from goldenrod genus—Solidago sp. In Invasive Plant Species in Polish Wetland Ecosystems; Dajdok, Z., Pawlaczyk, P., Eds.; Klub Przyrodników: Świebodzin, Poland, 2009; pp. 80–86. (In Polish) [Google Scholar]
- Jezierska-Domaradzka, A.; Domaradzki, K. Solidago canadensis L. as a potential energy plant—The risk to the environment and the estimation of natural raw material resources in the chosen fallow fields in Wołów County (Lower Silesia). Zesz. Nauk. UP Wroc. Rol. C 2012, 584, 43–52. (In Polish) [Google Scholar]
- Rola, J.; Rola, H. Solidago spp. as bioindicator of fallow occurrence on arable area. Fragm. Agron. 2010, 27, 122–131. (In Polish) [Google Scholar]
- Feledyn-Szewczyk, B.; Matyka, M.; Staniak, M. Comparison of the effect of perennial energy crops and agricultural crops on weed flora diversity. Agronomy 2019, 9, 695. [Google Scholar] [CrossRef] [Green Version]
- Szépligeti, M.; Kun, R.; Bartha, S.; Bodonczi, L.; Szentirmai, I. Experience gained from the control of giant goldenrod in the Őrség National Park. In Practical Experiences in Invasive Alien Plant Control, 2nd ed.; Csiszár, Á., Korda, M., Eds.; Duna-Ipoly National Park Directorate: Budapest, Hungary, 2017; pp. 135–139. [Google Scholar]
- Szymura, M.; Szymura, T.H.; Wolski, K. Invasive Solidago species: How large area do they occupy and what would be the cost of their removal? Pol. J. Ecol. 2016, 64, 25–34. [Google Scholar] [CrossRef]
- Boutin, C.; Strandberg, B.; Carpenter, D.; Mathiassen, S.K.; Thomas, P.J. Herbicide impact on non-target plant reproduction: What are the toxicological and ecological implications? Environ. Pollut. 2014, 185, 295–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- IUSS Working Group World Reference Base. World Reference Base for Soil Resources 2014, Update 2015—International Soil Classification System for Naming Soils and Creating Legends for Soil Maps; Report No. 106; FAO: Rome, Italy, 2015. [Google Scholar]
- Woś, A. The Climate of Poland in the Second Half of the 20th Century; Adam Mickiewicz University: Poznań, Poland, 2010; p. 489. (In Polish) [Google Scholar]
- Dąbkowska, T.; Stupnicka-Rodzynkiewicz, E.; Bintsanga-Malounguidi, P. The effect of weather conditions and weed control measures on maize weeds development, particularly on Echinochloa crusgalli. Ann. UMCS Sec. E Agric. 2007, 52, 117–126. (In Polish) [Google Scholar]
- Zhang, Y.; Yang, X.; Zhu, Y.; Li, L.; Zhang, Y.; Li, J.; Song, X.; Qiang, S. Biological control of Solidago canadensis using a bioherbicide isolate of Sclerotium rolfsii SC64 increased the biodiversity in invaded habitats. Biol. Control 2019, 139, 104093. [Google Scholar] [CrossRef]
- Tang, W.; Kuang, J.; Qiang, S. Biological control of the invasive alien weed Solidago canadensis: Combining an indigenous fungal isolate of Sclerotium rolfsii SC64 with mechanical control. Biocontrol Sci. Technol. 2013, 23, 1123–1136. [Google Scholar] [CrossRef]
- Yuan, G. Study on the control of Solidago canadensis by spraying metsulfuron-methyl. J. Anhui Agric. Sci. 2008, 32, 14188–14189. [Google Scholar]
- Hartmann, E.; Schuldes, H.; Kubler, R.; Konold, W. Neophyten: Biologie, Verbreitung und Kontrolle Ausgewählter Arten; Ecomed: Landsberg, Germany, 1995; p. 301. [Google Scholar]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | 10-Days Period | Month | |||||||
|---|---|---|---|---|---|---|---|---|---|
| IV | V | VI | VII | VIII | IX | X | IV–X | ||
| 2015 | 1 | 4.2 | 13.3 | 19.1 | 21.1 | 23.3 | 15.2 | 9.5 | 15.1 |
| 2 | 9.7 | 13.2 | 17.7 | 20.0 | 21.8 | 17.8 | 6.8 | 15.3 | |
| 3 | 12.1 | 12.6 | 15.8 | 20.7 | 19.6 | 11.9 | 6.8 | 14.2 | |
| Mean | 8.7 | 13.0 | 17.5 | 20.6 | 21.5 | 15.0 | 7.7 | 14.9 | |
| 2016 | 1 | 10.0 | 12.5 | 16.3 | 19.0 | 18.7 | 18.5 | 8.9 | 14.8 |
| 2 | 10.3 | 11.7 | 18.1 | 18.2 | 16.6 | 16.5 | 7.4 | 14.1 | |
| 3 | 6.5 | 18.2 | 20.0 | 20.9 | 18.6 | 10.6 | 7.3 | 14.6 | |
| Mean | 9.0 | 14.1 | 18.1 | 19.4 | 18.0 | 15.2 | 7.8 | 14.5 | |
| 1951–2000 | Mean | 8.0 | 13.1 | 16.4 | 17.9 | 17.3 | 13.3 | 8.4 | 13.5 |
| Year | 10-Days Period | Month | |||||||
|---|---|---|---|---|---|---|---|---|---|
| IV | V | VI | VII | VIII | IX | X | IV–X | ||
| 2015 | 1 | 14.0 | 37.3 | 0.0 | 5.1 | 3.8 | 11.4 | 0.0 | 71.6 |
| 2 | 0.8 | 20.8 | 20.8 | 23.1 | 61.0 | 29.2 | 20.3 | 176.0 | |
| 3 | 27.4 | 45.7 | 15.0 | 14.2 | 4.3 | 28.5 | 5.6 | 140.7 | |
| Sum | 42.2 | 103.9 | 35.8 | 42.4 | 69.1 | 69.1 | 25.9 | 388.4 | |
| 2016 | 1 | 14.0 | 12.7 | 25.7 | 31.0 | 74.7 | 11.2 | 77.5 | 246.8 |
| 2 | 22.4 | 24.4 | 21.3 | 70.9 | 20.8 | 7.4 | 18.0 | 185.2 | |
| 3 | 15.5 | 8.9 | 9.4 | 78.2 | 18.3 | 1.3 | 32.0 | 163.6 | |
| Sum | 51.8 | 46.0 | 56.4 | 180.1 | 113.8 | 19.8 | 127.5 | 595.4 | |
| 1951–2000 | Mean | 49.0 | 76.0 | 89.0 | 93.0 | 81.0 | 54.0 | 45.0 | 487.0 |
| 2015 | 2016 |
|---|---|
| June 2. | June 10. |
| June 16. | June 25. |
| June 30. | July 9. |
| July 14. | July 23. |
| July 28. | August 6. |
| August 11. | August 20. |
| August 25. | September 3. |
| September 9. | September 17. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gala-Czekaj, D.; Synowiec, A.; Dąbkowska, T. Self-Renewal of Invasive Goldenrods (Solidago spp.) as a Result of Different Mechanical Management of Fallow. Agronomy 2021, 11, 1065. https://doi.org/10.3390/agronomy11061065
Gala-Czekaj D, Synowiec A, Dąbkowska T. Self-Renewal of Invasive Goldenrods (Solidago spp.) as a Result of Different Mechanical Management of Fallow. Agronomy. 2021; 11(6):1065. https://doi.org/10.3390/agronomy11061065
Chicago/Turabian StyleGala-Czekaj, Dorota, Agnieszka Synowiec, and Teresa Dąbkowska. 2021. "Self-Renewal of Invasive Goldenrods (Solidago spp.) as a Result of Different Mechanical Management of Fallow" Agronomy 11, no. 6: 1065. https://doi.org/10.3390/agronomy11061065
APA StyleGala-Czekaj, D., Synowiec, A., & Dąbkowska, T. (2021). Self-Renewal of Invasive Goldenrods (Solidago spp.) as a Result of Different Mechanical Management of Fallow. Agronomy, 11(6), 1065. https://doi.org/10.3390/agronomy11061065