
Are Wild Blueberries a Crop with Low Photosynthetic Capacity? Chamber-Size Effects in Measuring Photosynthesis



Abstract
:1. Introduction
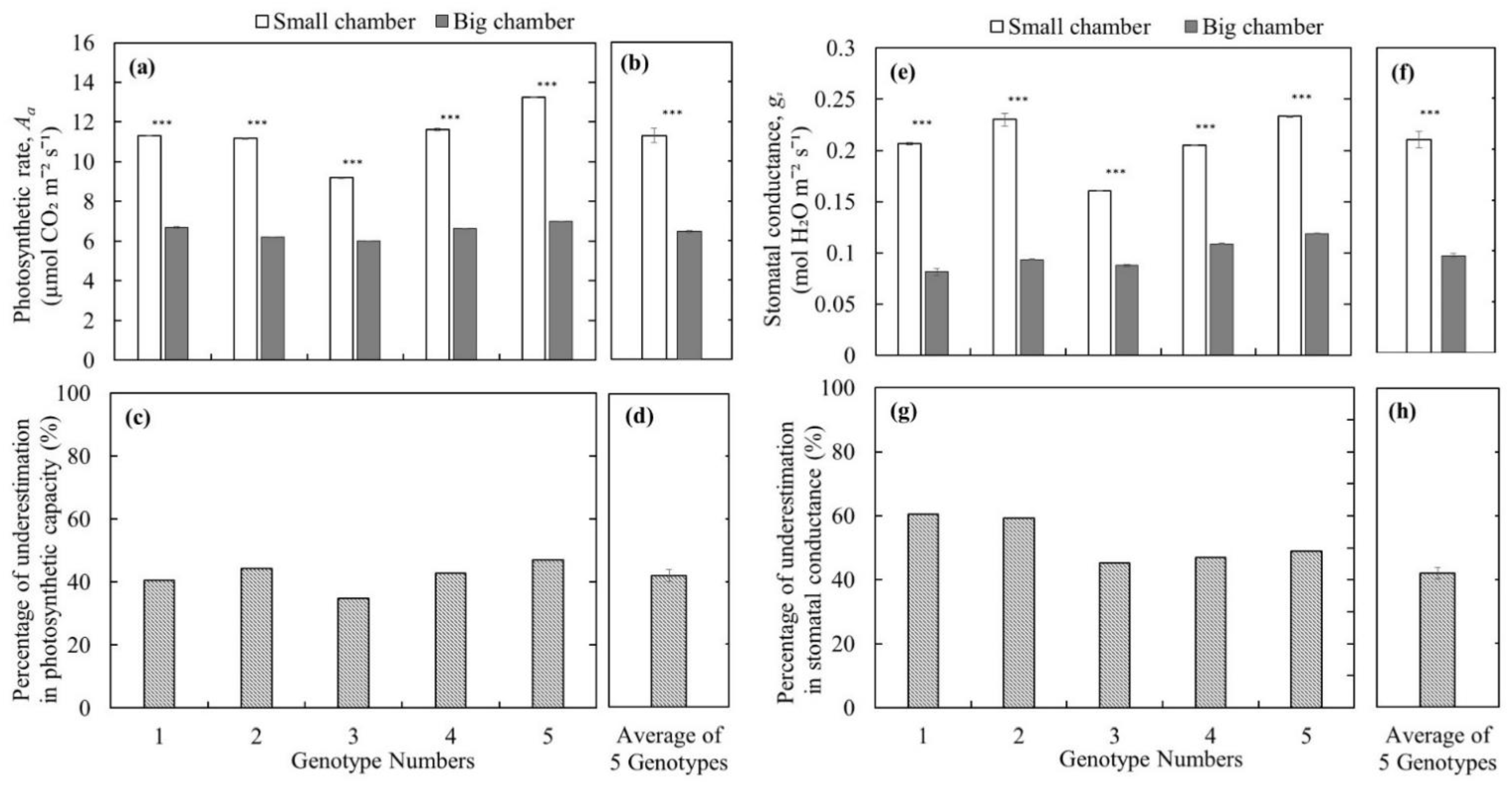
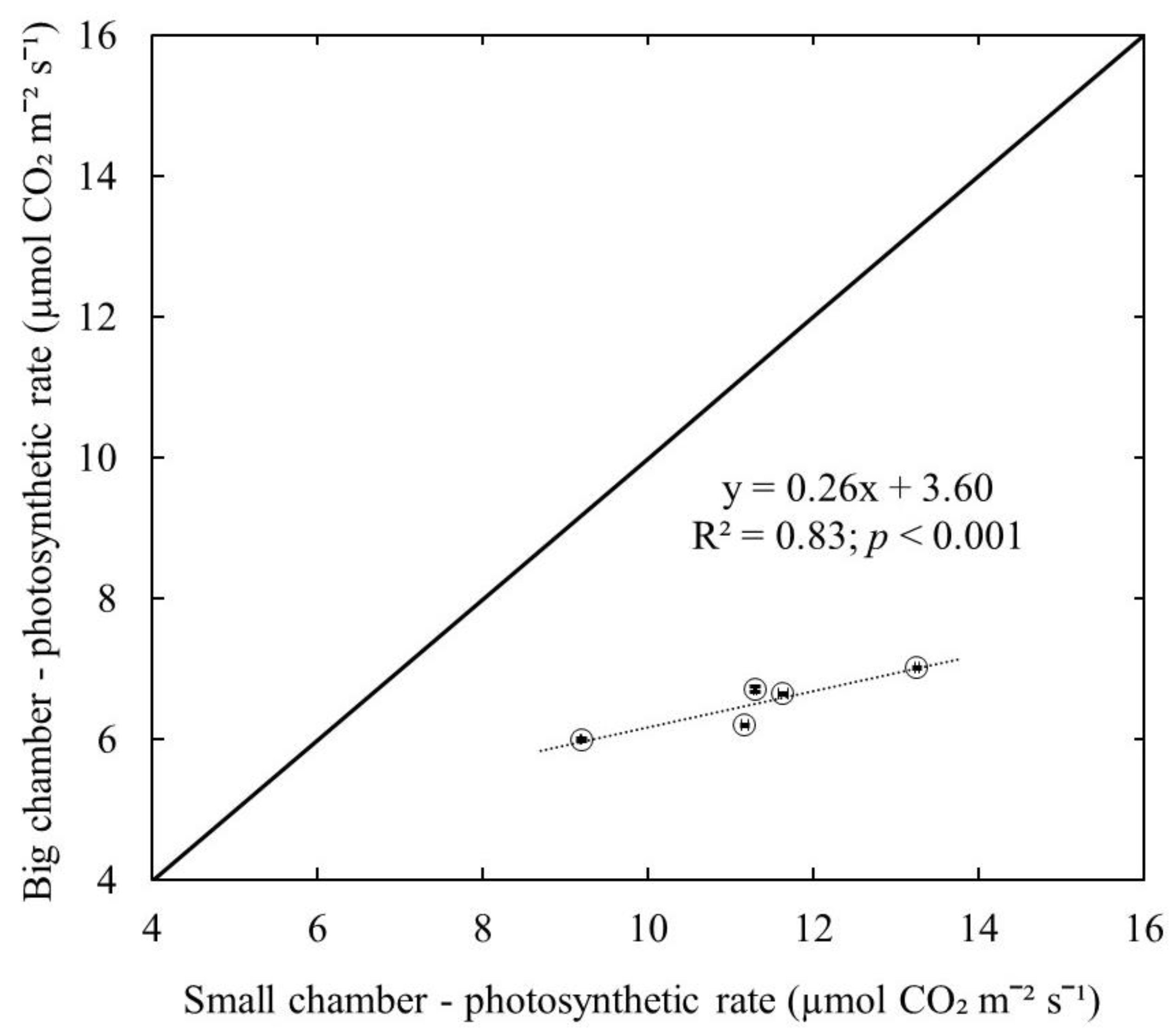
- To test whether using the big leaf chamber (branch level measurements) underestimates the leaf photosynthetic capacity compared to the measurements with a small leaf chamber enclosing a single leaf (leaf level measurements), and whether wild blueberry plants are not a crop with low photosynthetic capacity;
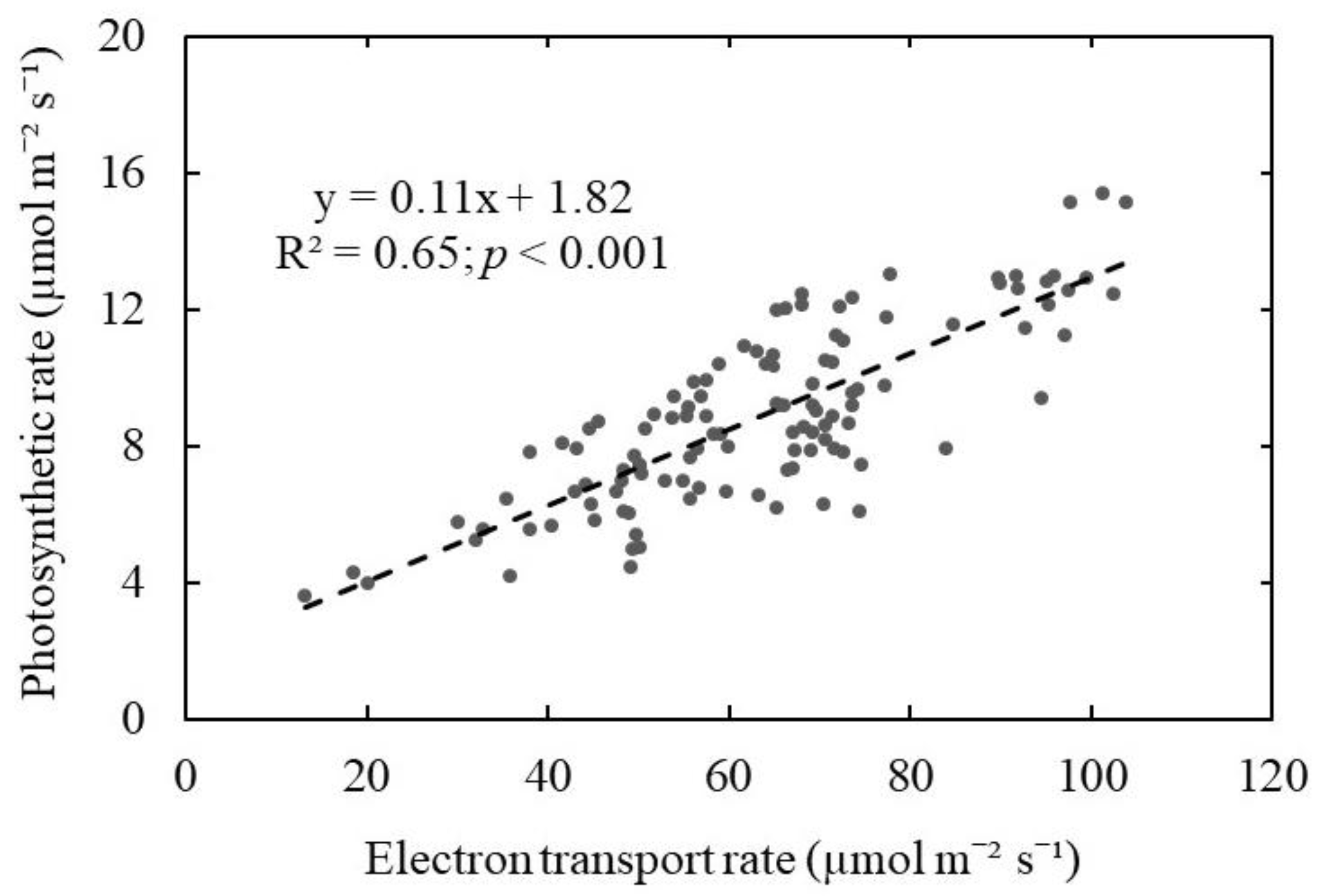
- To test whether the electron transport rate measured by a chlorophyll fluorescence meter can be used to predict the photosynthetic rate measured by the gas exchange system for wild blueberry plants.
2. Materials and Methods
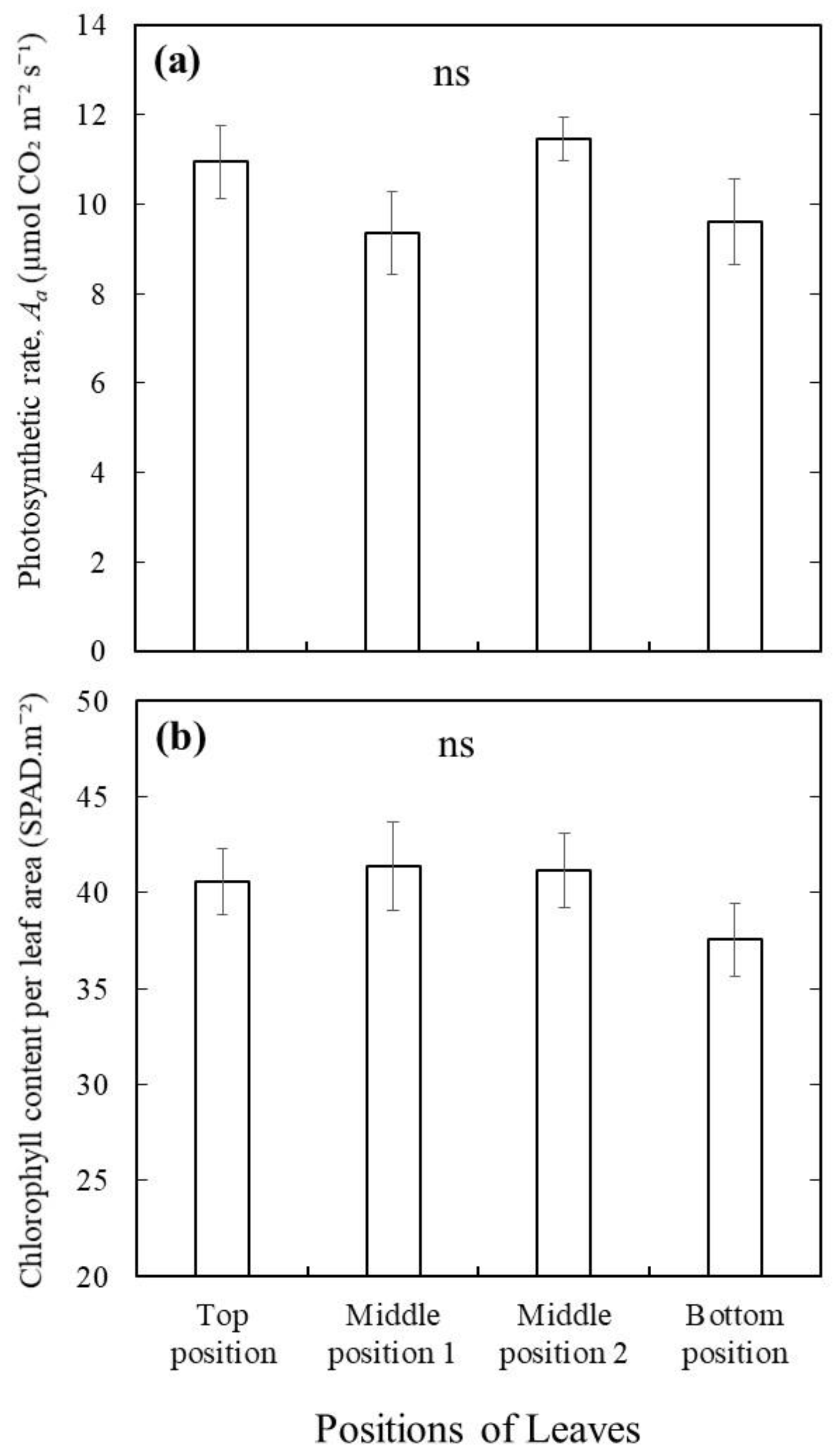
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Evans, L.T. Plant physiologist as midwife. Search 1977, 8, 262–268. [Google Scholar]
- Kramer, P.J. The role of physiology in crop improvement. In Linking Research to Crop Production; Springer: Boston, MA, USA, 1990; pp. 51–62. [Google Scholar]
- Schrader, L.E. Contributions from biochemistry and plant physiology. Mov. Up Yield Curve Adv. Obs. 1980, 39, 25–43. [Google Scholar]
- Whan, B.R.; Carlton, G.P.; Siddique, K.H.M.; Regan, K.L.; Turner, N.C.; Anderson, W.K. Integration of breeding and physiology: Lessons from a water-limited environment. Int. Crop. Sci. I 1993, 607–614. [Google Scholar] [CrossRef]
- El-Sharkawy, M.A. How can calibrated research-based models be improved for use as a tool in identifying genes controlling crop tolerance to environmental stresses in the era of genomics—from an experimentalist’s perspective. Photosynthetica 2005, 43, 161–176. [Google Scholar] [CrossRef]
- Lawson, T.; Kramer, D.M.; Raines, C.A. Improving yield by exploiting mechanisms underlying natural variation of photosynthesis. Curr. Opin. Biotechnol. 2012, 23, 215–220. [Google Scholar] [CrossRef]
- Faralli, M.; Lawson, T. Natural genetic variation in photosynthesis: An untapped resource to increase crop yield potential? Plant J. 2020, 101, 518–528. [Google Scholar] [CrossRef]
- Long, S.P.; Zhu, X.G.; Naidu, S.L.; Ort, D.R. Can improvement in photosynthesis increase crop yields? Plant Cell Environ. 2006, 29, 315–330. [Google Scholar] [CrossRef]
- Taiz, L.; Zeiger, E.; Møller, I.M.; Murphy, A. Plant Physiology and Development, 6th ed.; Sinauer Associates Incorporated: Sunderland, MA, USA, 2015. [Google Scholar]
- De Wit, C.T. Photosynthesis of Leaf Canopies; Centre for Agricultural Publications and Documentation: Wageningen, The Netherlands, 1965; Pudoc (Agricultural research reports 663) - 57. [Google Scholar]
- Duncan, W.G.R.S.; Loomis, R.; Williams, W.; Hanau, R. A model for simulating photosynthesis in plant communities. Hilgardia 1967, 38, 181–205. [Google Scholar] [CrossRef] [Green Version]
- Baker, D.N. Simulation for research and crop management. In Proceedings of World Soybean Research Conference II, 1980; Corbin, F.T., Ed.; Westview Press: Boulder, CO, USA, 1979; pp. 533–546. [Google Scholar]
- Bunce, J.A.; Heichel, G.H. Measurements and modeling of photosynthesis in field crops. Crit. Rev. Plant Sci. 1986, 4, 47–77. [Google Scholar] [CrossRef]
- Penning de Vries, F.W.T.; Jansen, D.M.; Ten Berge, H.F.M.; Bakema, A. Simulation of Ecophysiological Processes of Growth in Several Annual Crops; PUDOC: Wageningen, The Netherlands, 1989. [Google Scholar]
- Boote, K.J.; Loomis, R.S. Modeling Crop Photosynthesis—From Biochemistry to Canopy; Crop Science Society of America and American Society of Agronomy: Madison, WI, USA, 1991. [Google Scholar]
- Hicklenton, P.R.; Reekie, J.Y.; Gordon, R.J.; Percival, D.C. Seasonal patterns of photosynthesis and stomatal conductance in lowbush blueberry plants managed in a two-year production cycle. HortScience 2000, 35, 55–59. [Google Scholar] [CrossRef]
- Percival, D.; Murray, A.; Stevens, D. Drought stress dynamics of wild blueberry (Vaccinium angustifolium Aiton). Acta Hortic. 2003, 618, 353–362. [Google Scholar] [CrossRef]
- Percival, D.; Kaur, J.; Hainstock, L.J.; Privé, J.P. Seasonal changes in photochemistry, light use efficiency and net photosynthetic rates of wild blueberry (Vaccinium angustifolium Ait.). Can. J. Plant Sci. 2012, 92, 1135–1143. [Google Scholar] [CrossRef] [Green Version]
- Tasnim, R.; Calderwood, L.; Annis, S.; Drummond, F.; Zhang, Y.J.; The future of wild blueberries: Testing warming impacts using open-top chambers. Spire Maine J. Conserv. Sustain. 2020. Available online: https://umaine.edu/spire/2020/02/10/wildblueberries/ (accessed on 22 February 2021).
- Van der Kloet, S.P. Systematics, distribution, and nomenclature of the polymorphic Vaccinium Angustifolium. Rhodora 1978, 80, 358–376. [Google Scholar]
- Sanderson, K.R.; Cutcliffe, J.A. Effect of sawdust mulch on yields of select clones of lowbush blueberry. Can. J. Plant Sci. 1991, 71, 1263–1266. [Google Scholar] [CrossRef] [Green Version]
- Forsyth, F.R.; Hall, I.V. Effect of leaf maturity, temperature, carbon dioxide concentration, and light intensity on rate of photosynthesis in clonal lines of the lowbush blueberry, Vaccinium angustifolium Ait. under laboratory conditions. Can. J. Bot. 1965, 43, 893–900. [Google Scholar] [CrossRef]
- Tasnim, R.; Drummond, F.; Zhang, Y.J. Climate Change Patterns of Wild Blueberry Fields in Downeast, Maine over the Past 40 Years. Water 2021, 13, 594. [Google Scholar] [CrossRef]
- Flexas, J.; Díaz-Espejo, A.; Berry, J.A.; Cifre, J.; Galmés, J.; Kaldenhoff, R.; Medrano, H.; Ribas-Carbó, M. Analysis of leakage in IRGA’s leaf chambers of open gas exchange systems: Quantification and its effects in photosynthesis parameterization. J. Exp. Bot. 2007, 58, 1533–1543. [Google Scholar] [CrossRef] [Green Version]
- Pons, T.L.; Welschen, R.A.M. Overestimation of respiration rates in commercially available clamp-on leaf chambers. Complications with measurement of net photosynthesis. Plant Cell Environ. 2002, 25, 1367–1372. [Google Scholar] [CrossRef]
- Sadras, V.O.; Lawson, C.; Montoro, A. Photosynthetic traits in Australian wheat varieties released between 1958 and 2007. Field Crop. Res. 2012, 134, 19–29. [Google Scholar] [CrossRef]
- Thomas, S.C. Photosynthetic capacity peaks at intermediate size in temperate deciduous trees. Tree Physiol. 2010, 30, 555–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ort, D.; Long, S.P. Converting solar energy into crop production. In Plants, Genes, and Crop Biotechnology; Chrispeels, M.J., Sadava, D.E., Eds.; Jones and Bartlett Publisher International: Burlington, MA, USA, 2003; pp. 240–269. [Google Scholar]
- Long, S.P.; Zhu, X.G.; Naidu, S.L.; Raines, C.A.; Ort, D.R. Limits to efficiencies of primary production-constraints and opportunities. In Yields of Farmed Species: Constraints and Opportunities in the 21st Century. In Proceedings of a University of Nottingham Easter School Series, Sutton Bonington, UK, June 2004; Nottingham University Press: Sutton Bonington, UK, 2005; pp. 167–191. [Google Scholar]
- Schreiber, U. Pulse-amplitude-modulation (PAM) fluorometry and saturation pulse method: An overview. Chlorophyll Fluoresc. 2004, 279–319. [Google Scholar] [CrossRef]
- Genty, B.; Briantais, J.M.; Baker, N.R. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochim. Biophys. Acta Gen. Subj. 1989, 990, 87–92. [Google Scholar] [CrossRef]
- Genty, B.; Harbinson, J.; Briantais, J.M.; Baker, N.R. The relationship between non-photochemical quenching of chlorophyll fluorescence and the rate of photosystem 2 photochemistry in leaves. Photosynth. Res. 1990, 25, 249–257. [Google Scholar] [CrossRef] [PubMed]
- U.S Department of Agriculture-National Agricultural Statistics Service. Noncitrus Fruits and Nuts 2003 Summary. 2004. Available online: http://usda.mannlib.cornell.edu/reports/nassr/fruit/pnfbb/ (accessed on 5 May 2021).







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| References | Time | Location of Field Study | Photosynthesis Measurement Method | Net Photosynthetic Rate (µmol.m−2.s−1) | ||
|---|---|---|---|---|---|---|
| Average | Range | Vegetative Year (August) | ||||
| Hicklenton et al. (2000) | 1996 to 1997 | Agriculture and Agri-Food Canada Experimental Farm, Sheffield Mills, N.S. (45° N, 64.5° W) | ADC LCA-2 portable open-flow leaf gas exchange system (ADC Ltd., Hoddesdon, UK) with temperature- controlled big leaf chamber (PLC 2A) | 5.0 to 6.0 | 1.0 to 10.0 | 6.5 to 9.5 |
| Percival et al. (2003) | 2000 | Wild Blueberry Research Centre (WBRC), Debert, Nova Scotia (45°27′ N, 62°36′ W) | ADC LCA-4 portable leaf gas exchange system (ADC Ltd., Hoddesdon, UK) with big leaf chamber | 4.0 to 5.0 | 0.5 to 8.5 | N/A |
| Percival et al. (2012) | 1999 to 2000 | LI-6250 portable photosynthesis measurement system (LI-COR Inc.) with big leaf chamber | 3.5 to 4.0 | 0.0 to 7.6 | 5.5 to 7.6 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tasnim, R.; Zhang, Y.-J. Are Wild Blueberries a Crop with Low Photosynthetic Capacity? Chamber-Size Effects in Measuring Photosynthesis. Agronomy 2021, 11, 1572. https://doi.org/10.3390/agronomy11081572
Tasnim R, Zhang Y-J. Are Wild Blueberries a Crop with Low Photosynthetic Capacity? Chamber-Size Effects in Measuring Photosynthesis. Agronomy. 2021; 11(8):1572. https://doi.org/10.3390/agronomy11081572
Chicago/Turabian StyleTasnim, Rafa, and Yong-Jiang Zhang. 2021. "Are Wild Blueberries a Crop with Low Photosynthetic Capacity? Chamber-Size Effects in Measuring Photosynthesis" Agronomy 11, no. 8: 1572. https://doi.org/10.3390/agronomy11081572
APA StyleTasnim, R., & Zhang, Y. -J. (2021). Are Wild Blueberries a Crop with Low Photosynthetic Capacity? Chamber-Size Effects in Measuring Photosynthesis. Agronomy, 11(8), 1572. https://doi.org/10.3390/agronomy11081572