
Detection of Cacao Mild Mosaic Virus (CaMMV) Using Nested PCR and Evidence of Uneven Distribution in Leaf Tissue


Abstract
:1. Introduction
2. Materials and Methods
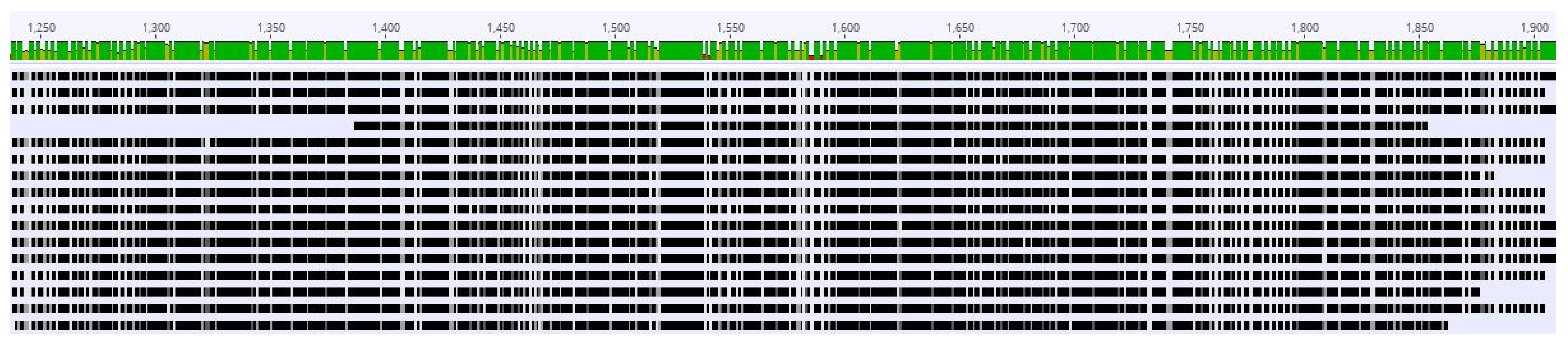
2.1. Confirmation of CaMMV
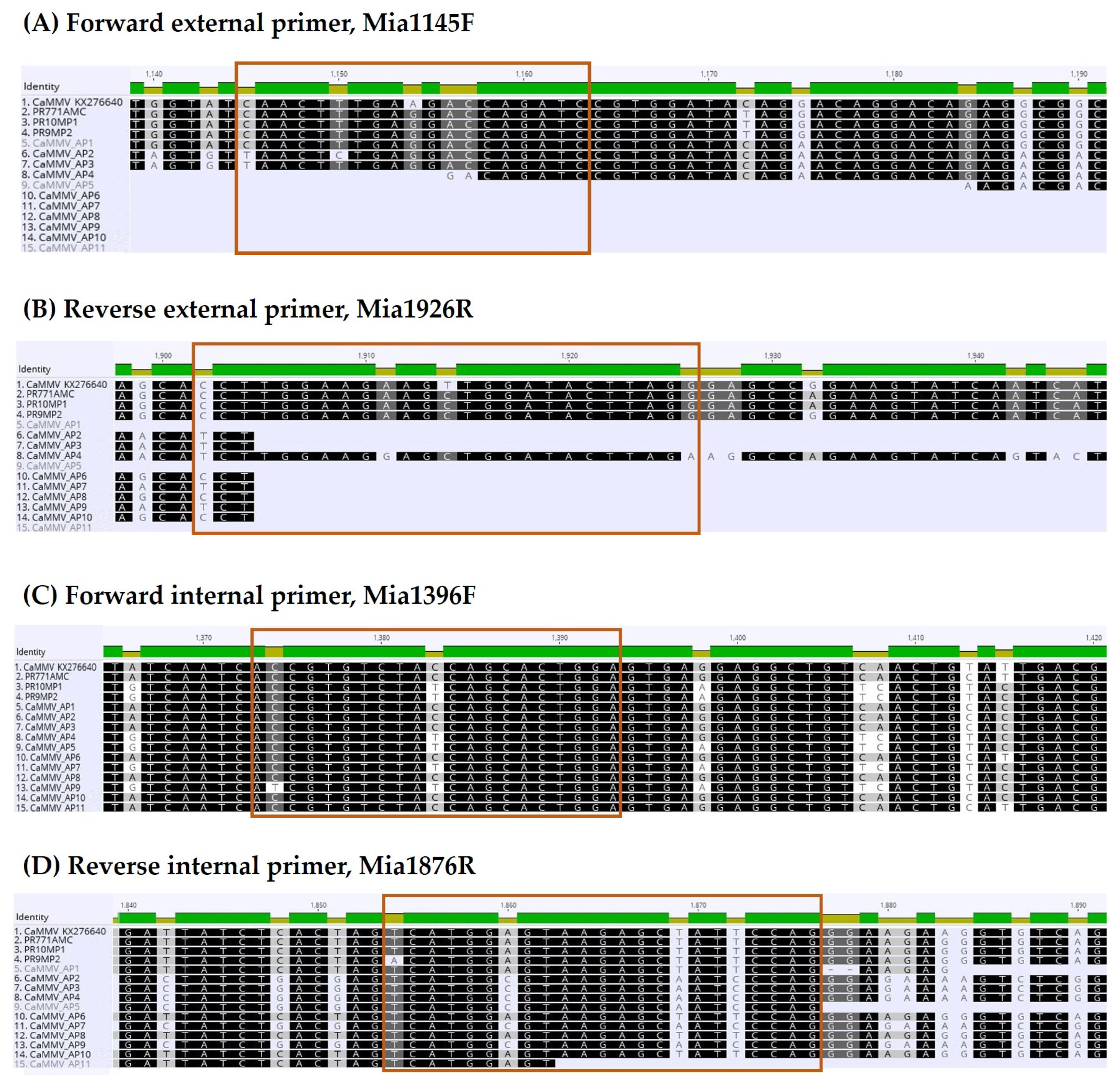
2.2. Primer Design
2.3. Nested PCR
2.4. Detection of CaMMV in Theobroma cacao
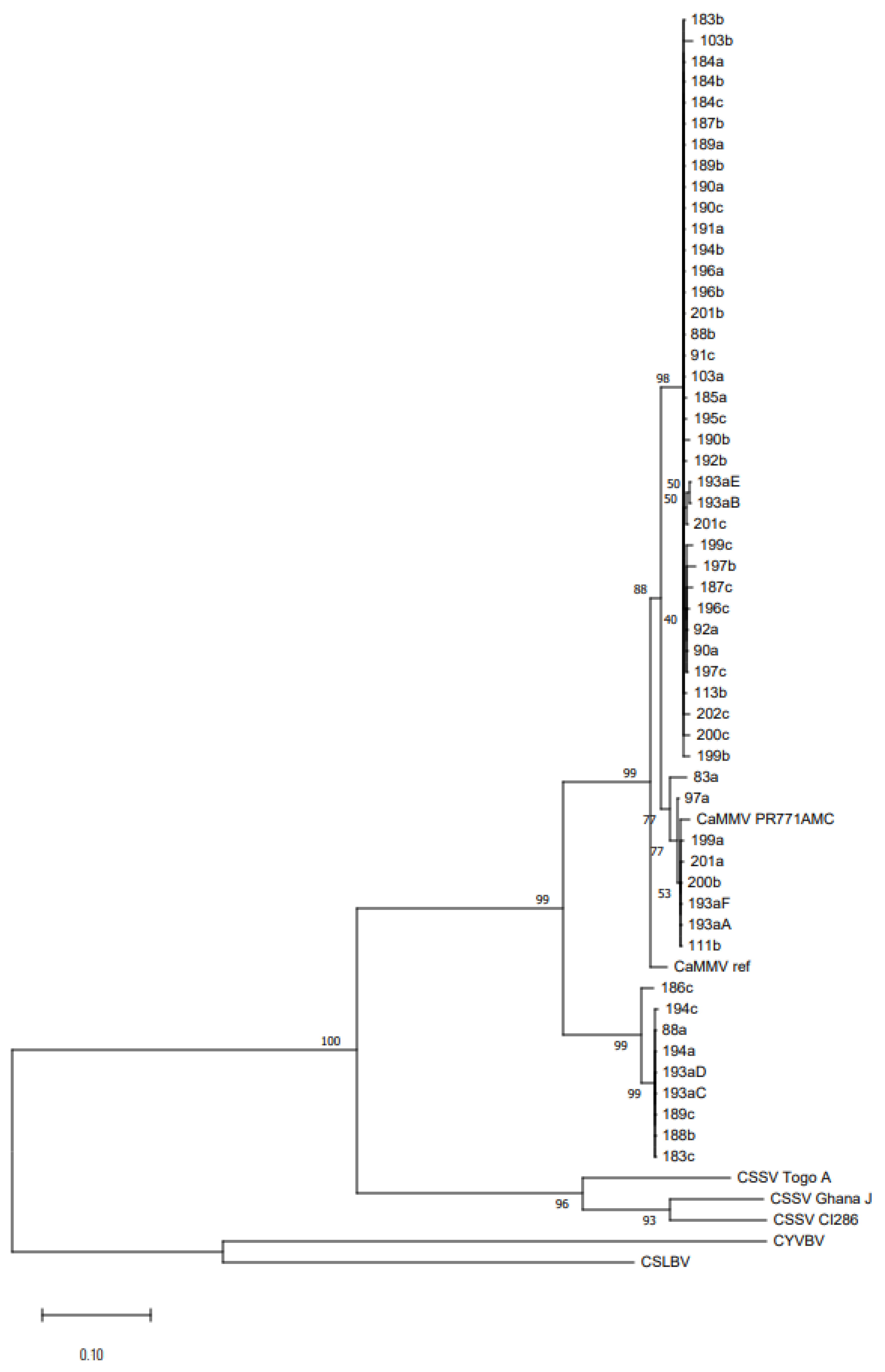
2.5. Phylogenetic Comparison
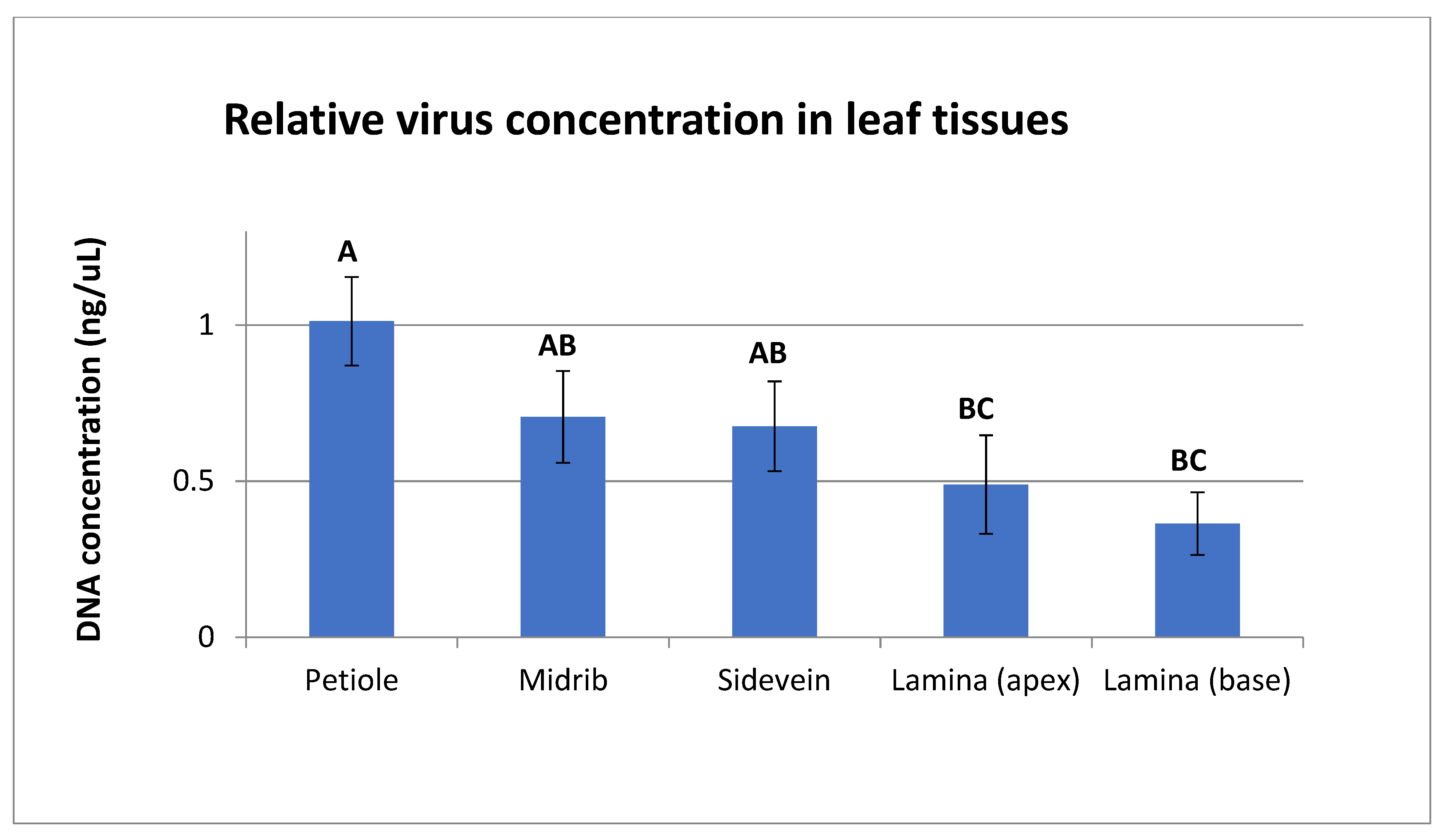
2.6. Quantification of Virus Titer in Different Leaf Tissues
3. Results
3.1. Confirmation of CaMMV
3.2. Primer Design and Nested PCR
3.3. Detection of CaMMV in Theobroma cacao
3.4. Phylogenetic Comparison
3.5. Quantification of Virus Titer in Different Leaf Tissues
4. Discussion
5. Conclusions
Supplementary Materials
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Posnette, A. Virus diseases of cacao in Trinidad. Trop. Agric. 1944, 21, 105–107. [Google Scholar]
- Cope, F. Statistical studies in the effects of virus infection upon yield in clonal cacao. Rep. Cacao Res. 1945-1951 1953, 4, 126–129. [Google Scholar]
- Swarbrick, J. Cacao virus in Trinidad. Trop. Agric. 1961, 38, 245–249. [Google Scholar]
- Sreenivasan, T. The Enigma of the ICS 76 Plants at Reading, UK; International Cocoa Quarantine Center: Reading, UK, 2009. [Google Scholar]
- Puig, A.; Ramos-Sobrinho, R.; Keith, C.; Kitchen, N.; Gutierrez, O.; Goenaga, R.; Brown, J.K. First Report of Cacao Mild Mosaic Virus Associated with Symptomatic Commercial Cacao (Theobroma cacao) Trees in Puerto Rico. Plant Dis. 2020, 104, 3089. [Google Scholar] [CrossRef]
- Ramos-Sobrinho, R.; Ferro, M.M.; Nagata, T.; Puig, A.S.; Von Keith, C.; Britto, D.S.; Gutierrez, O.A.; Marelli, J.-P.; Brown, J.K. Complete genome sequences of three newly discovered cacao mild mosaic virus isolates from Theobroma cacao L. in Brazil and Puerto Rico and evidence for recombination. Arch. Virol. 2021, 166, 2027–2031. [Google Scholar] [CrossRef]
- Fisher, H.; Haun, J.; Ackerman, W. Cacao seedling production and distribution through plant quarantine. Cacao 1960, 5, 1–8. [Google Scholar]
- Thresh, J. Quarantine arrangements for intercepting Cocoa material infected with West African viruses. Plant Prot. Bull. FAO 1960, 8, 89–92. [Google Scholar]
- Posnette, A.; Palma, M. Observations on cacao on the Paria peninsula, Venezuela. Trop. Agric. 1944, 21, 130–132. [Google Scholar]
- Muller, E.; Ravel, S.; Agret, C.; Abrokwah, F.; Dzahini-Obiatey, H.; Galyuon, I.; Kouakou, K.; Jeyaseelan, E.; Allainguillaume, J.; Wetten, A. Next generation sequencing elucidates cacao badnavirus diversity and reveals the existence of more than ten viral species. Virus Res. 2018, 244, 235–251. [Google Scholar] [CrossRef] [Green Version]
- Probowati, W.; Somowiyarjo, S.; Hartono, S. Molecular characterization of Mosaic Virus from the cocoa trees showing mosaic symptoms in Yogyakarta, Indonesia. Biodiversitas J. Biol. Divers. 2019, 20, 3698–3704. [Google Scholar] [CrossRef]
- Thresh, J. The control of cacao swollen shoot disease in West Africa: A review of the present situation. Tech. Bull. West Afr. Cocoa Res. Inst. 1958, 4, 36. [Google Scholar]
- Muller, E. Cacao Swollen Shoot Virus (CSSV): History, biology, and genome. In Cacao Diseases; Springer: Cham, Switzerland, 2016; pp. 337–358. [Google Scholar]
- Chingandu, N.; Sreenivasan, T.N.; Surujdeo-Maharaj, S.; Umaharan, P.; Gutierrez, O.A.; Brown, J.K. Molecular characterization of previously elusive badnaviruses associated with symptomatic cacao in the New World. Arch. Virol. 2017, 162, 1363–1371. [Google Scholar] [CrossRef]
- Chingandu, N.; Dongo, L.; Gutierrez, O.A.; Brown, J.K. The previously unidentified, divergent badnavirus species Cacao red vein-banding virus is associated with Cacao swollen shoot disease in Nigeria. Plant Dis. 2019, 103, 1302–1308. [Google Scholar] [CrossRef]
- Ding, S.-W.; Mackenzie, A.; Torronen, M.; Gibbs, A. Nucleotide sequence of the virion protein gene of cacao yellow mosaic tymovirus. Nucleic Acids Res. 1990, 18, 5886. [Google Scholar] [CrossRef] [PubMed]
- Kenten, R. The purification and some properties of cocoa necrosis virus, a serotype of tomato black ring virus. Ann. Appl. Biol. 1972, 71, 119–126. [Google Scholar] [CrossRef]
- Brunt, A.; Kenten, R.; Gibbs, A.; Nixon, H. Further studies on cocoa yellow mosaic virus. Microbiology 1965, 38, 81–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muller, E.; Sackey, S. Molecular variability analysis of five new complete cacao swollen shoot virus genomic sequences. Arch. Virol. 2005, 150, 53–66. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Hall, B.G. Building phylogenetic trees from molecular data with MEGA. Mol. Biol. Evol. 2013, 30, 1229–1235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, K. Estimation of the number of nucleotide substitutions when there are strong transition-transversion and G+ C-content biases. Mol. Biol. Evol. 1992, 9, 678–687. [Google Scholar] [PubMed] [Green Version]
- Baker, R.; Dale, W. Virus diseases of cacao in Trinidad II. Trop. Agric. 1947, 24, 127–130. [Google Scholar]
- Kirkpatrick, T. Insect Pests of Cacao and Insect Vectors of Cacao Virus Disease. Rep. Cacao Res. 1953, 5, 122–125. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Comments | Primer Name | Sequence (5′–3′) | Length | Amplicon (bp) |
|---|---|---|---|---|
| Nested PCR (outer) | Mia1145F | YAACTTTGAGGACCAGATC | 19 bp | 806 |
| Mia1926 | YCTAAGTATCCARCTYCTTCCAAGR | 25 bp | ||
| Nested PCR (inner) | Mia1396F | ACCGTGTCTAYCAGCACTGGA | 21 bp | 503 |
| Mia1876R | CTGGRATWGCTCTTACKCCATGW | 23 bp | ||
| Semi-quantitative, virus-specific | Mia1396F | ACCGTGTCTAYCAGCACTGGA | 21 bp | 289 |
| Mia1667R | GACCACCGTCAGCCAGAC | 18 bp | ||
| Semi-quantitative, host-specific (for standardization) | TcACTF | CAGACTTTGAGTTCACTTGACACAG | 25 bp | ~200 |
| TcACTR | AGTGTCTGGATTGGAGGATCTATCT | 25 bp | ||
| Original diagnostic [5] | CaMMV-1112F | TACGTAACCATCATTGATGC | 20 bp | 867 |
| CaMMV-1959R | CTTCTTTACGTTCAATGTCC | 20 bp |
| Isolate | Species | Host | Origin | GenBank |
|---|---|---|---|---|
| 83a | Cacao mild mosaic virus | Theobroma cacao | USA | MZ325886 |
| 201a | Cacao mild mosaic virus | Theobroma cacao | USA | MZ325887 |
| 200b | Cacao mild mosaic virus | Theobroma cacao | USA | MZ325888 |
| 199a | Cacao mild mosaic virus | Theobroma cacao | USA | MZ325889 |
| 88a | Cacao mild mosaic virus | Theobroma cacao | USA | MZ325890 |
| 183c | Cacao mild mosaic virus | Theobroma cacao | USA | MZ325891 |
| 186c | Cacao mild mosaic virus | Theobroma cacao | USA | MZ325892 |
| 194c | Cacao mild mosaic virus | Theobroma cacao | USA | MZ325893 |
| 197b | Cacao mild mosaic virus | Theobroma cacao | USA | MZ325894 |
| 187c | Cacao mild mosaic virus | Theobroma cacao | USA | MZ325895 |
| 200c | Cacao mild mosaic virus | Theobroma cacao | USA | MZ325896 |
| 202c | Cacao mild mosaic virus | Theobroma cacao | USA | MZ325897 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Puig, A.S. Detection of Cacao Mild Mosaic Virus (CaMMV) Using Nested PCR and Evidence of Uneven Distribution in Leaf Tissue. Agronomy 2021, 11, 1842. https://doi.org/10.3390/agronomy11091842
Puig AS. Detection of Cacao Mild Mosaic Virus (CaMMV) Using Nested PCR and Evidence of Uneven Distribution in Leaf Tissue. Agronomy. 2021; 11(9):1842. https://doi.org/10.3390/agronomy11091842
Chicago/Turabian StylePuig, Alina S. 2021. "Detection of Cacao Mild Mosaic Virus (CaMMV) Using Nested PCR and Evidence of Uneven Distribution in Leaf Tissue" Agronomy 11, no. 9: 1842. https://doi.org/10.3390/agronomy11091842
APA StylePuig, A. S. (2021). Detection of Cacao Mild Mosaic Virus (CaMMV) Using Nested PCR and Evidence of Uneven Distribution in Leaf Tissue. Agronomy, 11(9), 1842. https://doi.org/10.3390/agronomy11091842