
Effects of Combining the Genes Controlling Anthocyanin and Melanin Synthesis in the Barley Grain on Pigment Accumulation and Plant Development

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Marker-Assisted Creation of the NIL
2.2. Phenotyping
2.3. Anthocyanin Extraction and Measurement
2.4. Cytological Analysis
2.5. Statistical Analysis
3. Results
3.1. Construction of the NIL
3.2. Comparative Analysis of the Pigmentation Time Course
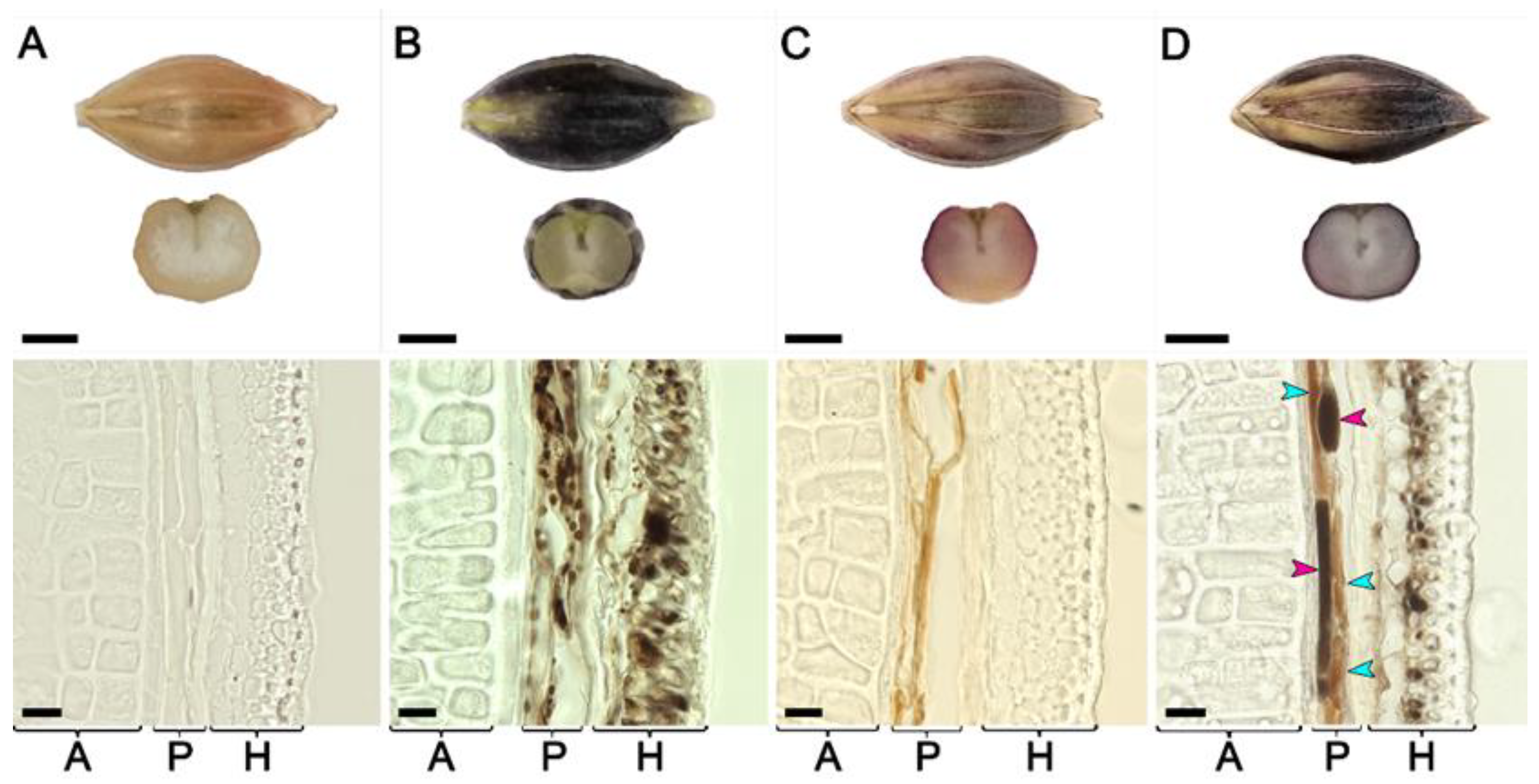
3.3. Cytological Analysis of Grain Pigmentation
3.4. The Anthocyanin Content of the Grain of NILs
3.5. Evaluation of Yield Components of the NILs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- FAOSTAT: Food and Agriculture Organization of the United Nations. Available online: https://www.fao.org/faostat/en/#home (accessed on 1 November 2021).
- Newton, A.C.; Flavell, A.J.; George, T.S.; Leat, P.; Mullholland, B.; Ramsay, L.; Revoredo-Giha, C.; Russell, J.; Steffenson, B.J.; Swanston, J.S.; et al. Crops That Feed the World 4. Barley: A Resilient Crop? Strengths and Weaknesses in the Context of Food Security. Food Secur. 2011, 3, 141–178. [Google Scholar] [CrossRef]
- Gangopadhyay, N.; Hossain, M.; Rai, D.; Brunton, N. A Review of Extraction and Analysis of Bioactives in Oat and Barley and Scope for Use of Novel Food Processing Technologies. Molecules 2015, 20, 10884–10909. [Google Scholar] [CrossRef]
- Ge, X.; Jing, L.; Zhao, K.; Su, C.; Zhang, B.; Zhang, Q.; Han, L.; Yu, X.; Li, W. The Phenolic Compounds Profile, Quantitative Analysis and Antioxidant Activity of Four Naked Barley Grains with Different Color. Food Chem. 2021, 335, 127655. [Google Scholar] [CrossRef]
- Kim, M.J.; Hyun, J.N.; Kim, J.A.; Park, J.C.; Kim, M.Y.; Kim, J.G.; Lee, S.J.; Chun, S.C.; Chung, I.M. Relationship between Phenolic Compounds, Anthocyanins Content and Antioxidant Activity in Colored Barley Germplasm. J. Agric. Food Chem. 2007, 55, 4802–4809. [Google Scholar] [CrossRef] [PubMed]
- Solano, F. Melanin and Melanin-Related Polymers as Materials with Biomedical and Biotechnological Applications—Cuttlefish Ink and Mussel Foot Proteins as Inspired Biomolecules. Int. J. Mol. Sci. 2017, 18, 1561. [Google Scholar] [CrossRef] [PubMed]
- Suriano, S.; Savino, M.; Codianni, P.; Iannucci, A.; Caternolo, G.; Russo, M.; Pecchioni, N.; Troccoli, A. Anthocyanin Profile and Antioxidant Capacity in Coloured Barley. Int. J. Food Sci. Technol. 2019, 54, 2478–2486. [Google Scholar] [CrossRef]
- Grotewold, E. The Science of Flavonoids; Springer: New York, NY, USA, 2006; ISBN 9780387288215. [Google Scholar]
- Ceccarelli, S.; Grando, S.; van Leur, J.A.G. Genetic Diversity in Barley Landraces from Syria and Jordan. Euphytica 1987, 36, 389–405. [Google Scholar] [CrossRef]
- Brunetti, C.; di Ferdinando, M.; Fini, A.; Pollastri, S.; Tattini, M. Flavonoids as Antioxidants and Developmental Regulators: Relative Significance in Plants and Humans. Int. J. Mol. Sci 2013, 14, 3540–3555. [Google Scholar] [CrossRef] [Green Version]
- Sharma, A.; Shahzad, B.; Rehman, A.; Bhardwaj, R.; Landi, M.; Zheng, B. Response of Phenylpropanoid Pathway and the Role of Polyphenols in Plants under Abiotic Stress. Molecules 2019, 24, 2452. [Google Scholar] [CrossRef] [Green Version]
- Williamson, G.; Manach, C. Bioavailability and Bioefficacy of Polyphenols in Humans. II. Review of 93 Intervention Studies. Am. J. Clin. Nutr. 2005, 81, 243S–255S. [Google Scholar] [CrossRef]
- Pandey, K.B.; Rizvi, S.I. Plant Polyphenols as Dietary Antioxidants in Human Health and Disease. Oxidative Med. Cell. Longev. 2009, 2, 270. [Google Scholar] [CrossRef] [Green Version]
- Olas, B. Berry Phenolic Antioxidants—Implications for Human Health? Front. Pharmacol. 2018, 9, 78. [Google Scholar] [CrossRef]
- Tena, N.; Martín, J.; Asuero, A.G. State of the Art of Anthocyanins: Antioxidant Activity, Sources, Bioavailability, and Therapeutic Effect in Human Health. Antioxidants 2020, 9, 451. [Google Scholar] [CrossRef] [PubMed]
- Tikhonova, M.A.; Shoeva, O.Y.; Tenditnik, M.V.; Ovsyukova, M.V.; Akopyan, A.A.; Dubrovina, N.I.; Amstislavskaya, T.G.; Khlestkina, E.K. Evaluating the Effects of Grain of Isogenic Wheat Lines Differing in the Content of Anthocyanins in Mouse Models of Neurodegenerative Disorders. Nutrients 2020, 12, 3877. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Liu, Y.; Pu, Z.; Wang, J.; Zheng, Y.; Li, Y.; Wei, Y. Regulation, Evolution, and Functionality of Flavonoids in Cereal Crops. Biotechnol. Lett. 2013, 35, 1765–1780. [Google Scholar] [CrossRef] [PubMed]
- Zeven, A.C. Wheats with Purple and Blue Grains: A Review. Euphytica 1991, 56, 243–258. [Google Scholar] [CrossRef]
- Abdel-Aal, E.S.M.; Young, J.; Rabalski, I. Anthocyanin Composition in Black, Blue, Pink, Purple, and Red Cereal Grains. J. Agric. Food Chem. 2006, 54, 4696–4704. [Google Scholar] [CrossRef] [PubMed]
- Glagoleva, A.Y.; Shoeva, O.Y.; Khlestkina, E.K. Melanin Pigment in Plants: Current Knowledge and Future Perspectives. Front. Plant Sci. 2020, 11, 770. [Google Scholar] [CrossRef]
- Mostert, A.B.; Powell, B.J.; Pratt, F.L.; Hanson, G.R.; Sarna, T.; Gentle, I.R.; Meredith, P. Role of Semiconductivity and Ion Transport in the Electrical Conduction of Melanin. Proc. Natl. Acad. Sci. USA 2012, 109, 8943–8947. [Google Scholar] [CrossRef] [Green Version]
- Wünsche, J.; Deng, Y.; Kumar, P.; di Mauro, E.; Josberger, E.; Sayago, J.; Pezzella, A.; Soavi, F.; Cicoira, F.; Rolandi, M.; et al. Protonic and Electronic Transport in Hydrated Thin Films of the Pigment Eumelanin. Chem. Mater. 2015, 27, 436–442. [Google Scholar] [CrossRef]
- Gracheva, N.V.; Zheltobryukhov, V.F. Sorption Properties of Sunflower Husk Melanins. Pharm. Chem. J. 2019, 53, 337–341. [Google Scholar] [CrossRef]
- Korpacheva, S.; Serasutdinova, K.; Lomovsky, I.; Chugunova, O. Technological Aspects of Obtaining Melanin and Powder from Buckwheat Hull and Their Use in Food Technology. E3S Web Conf. 2021, 296, 07007. [Google Scholar] [CrossRef]
- Cockram, J.; White, J.; Zuluaga, D.L.; Smith, D.; Comadran, J.; Macaulay, M.; Luo, Z.; Kearsey, M.J.; Werner, P.; Harrap, D.; et al. Genome-Wide Association Mapping to Candidate Polymorphism Resolution in the Unsequenced Barley Genome. Proc. Natl. Acad. Sci. USA 2010, 107, 21611–21616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Himi, E.; Taketa, S. Isolation of Candidate Genes for the Barley Ant1 and Wheat Rc Genes Controlling Anthocyanin Pigmentation in Different Vegetative Tissues. Mol. Genet. Genom. 2015, 290, 1287–1298. [Google Scholar] [CrossRef] [PubMed]
- Shoeva, O.Y.; Kukoeva, T.V.; Börner, A.; Khlestkina, E.K. Barley Ant1 Is a Homolog of Maize C1 and Its Product Is Part of the Regulatory Machinery Governing Anthocyanin Synthesis in the Leaf Sheath. Plant Breed. 2015, 134, 400–405. [Google Scholar] [CrossRef]
- Shoeva, O.Y.; Mock, H.-P.; Kukoeva, T.V.; Börner, A.; Khlestkina, E.K. Regulation of the Flavonoid Biosynthesis Pathway Genes in Purple and Black Grains of Hordeum Vulgare. PLoS ONE 2016, 11, e0163782. [Google Scholar] [CrossRef] [PubMed]
- Mol, J.; Grotewold, E.; Koes, R. How Genes Paint Flowers and Seeds. Trends Plant Sci. 1998, 3, 212–217. [Google Scholar] [CrossRef]
- Strygina, K.V.; Börner, A.; Khlestkina, E.K. Identification and Characterization of Regulatory Network Components for Anthocyanin Synthesis in Barley Aleurone. BMC Plant Biol. 2017, 17, 184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, Z.; Jia, Y.; Tan, C.; Zhang, X.-Q.; Angessa, T.; Broughton, S.; Westcott, S.; Dai, F.; Zhang, G.; Sun, D.; et al. Genetic Mapping and Evolutionary Analyses of the Black Grain Trait in Barley. Front. Plant Sci. 2019, 9, 1921. [Google Scholar] [CrossRef]
- Song, C.; Xiang, D.B.; Yan, L.; Song, Y.; Zhao, G.; Wang, Y.H.; Zhangb, B.L. Changes in Seed Growth, Levels and Distribution of Flavonoids during Tartary Buckwheat Seed Development. Plant Prod. Sci. 2016, 19, 518–527. [Google Scholar] [CrossRef] [Green Version]
- Druka, A.; Franckowiak, J.; Lundqvist, U.; Bonar, N.; Alexander, J.; Houston, K.; Radovic, S.; Shahinnia, F.; Vendramin, V.; Morgante, M.; et al. Genetic Dissection of Barley Morphology and Development. Plant Physiol. 2011, 155, 617–627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varshney, R.K.; Marcel, T.C.; Ramsay, L.; Russell, J.; Röder, M.S.; Stein, N.; Waugh, R.; Langridge, P.; Niks, R.E.; Graner, A. A High Density Barley Microsatellite Consensus Map with 775 SSR Loci. TAG. Theor. Appl. Genet. Theor. Und Angew. Genet. 2007, 114, 1091–1103. [Google Scholar] [CrossRef] [Green Version]
- Gordeeva, E.I.; Glagoleva, A.Y.; Kukoeva, T.V.; Khlestkina, E.K.; Shoeva, O.Y. Purple-Grained Barley (Hordeum vulgare L.): Marker-Assisted Development of NILs for Investigating Peculiarities of the Anthocyanin Biosynthesis Regulatory Network. BMC Plant Biol. 2019, 19, 49–57. [Google Scholar] [CrossRef]
- Abdel-Aal, E.-S.M.; Hucl, P. A Rapid Method for Quantifying Total Anthocyanins in Blue Aleurone and Purple Pericarp Wheats. Cereal Chem. 1999, 76, 350–354. [Google Scholar] [CrossRef]
- Shoeva, O.Y.; Mursalimov, S.R.; Gracheva, N.V.; Glagoleva, A.Y.; Börner, A.; Khlestkina, E.K. Melanin Formation in Barley Grain Occurs within Plastids of Pericarp and Husk Cells. Sci. Rep. 2020, 10, 179. [Google Scholar] [CrossRef]
- Francavilla, A.; Joye, I.J. Anthocyanins in Whole Grain Cereals and Their Potential Effect on Health. Nutrients 2020, 12, 2922. [Google Scholar] [CrossRef]
- Zhang, Y.; Yin, L.; Huang, L.; Tekliye, M.; Xia, X.; Li, J.; Dong, M. Composition, Antioxidant Activity, and Neuroprotective Effects of Anthocyanin-Rich Extract from Purple Highland Barley Bran and Its Promotion on Autophagy. Food Chem. 2021, 339, 127849. [Google Scholar] [CrossRef]
- Dhua, S.; Kumar, K.; Kumar, Y.; Singh, L.; Sharanagat, V.S. Composition, Characteristics and Health Promising Prospects of Black Wheat: A Review. Undefined 2021, 112, 780–794. [Google Scholar] [CrossRef]
- Kobylyansky, V.D.; Lukyanova, M.V. Flora of Cultivated Plants, Barley; Agropromizdat: Leningrad, Russia, 1990; Volume 2. [Google Scholar]
- Pecket, R.C.; Small, C.J. Occurrence, Location and Development of Anthocyanoplasts. Phytochemistry 1980, 19, 2571–2576. [Google Scholar] [CrossRef]
- Coutinho, J.W.; Rodrigues, A.C.; Appezzato-da-Glória, B.; Oliveira, E.M.; Oliveira, F.M.C.; Lusa, M.G. Plastid Role in Phytomelanin Synthesis in Piptocarpha Axillaris (Less.) Baker Stems (Asteraceae, Vernonieae). Protoplasma 2021, 258, 963–977. [Google Scholar] [CrossRef]
- Downie, A.B.; Zhang, D.; Dirk, L.M.A.; Thacker, R.R.; Pfeiffer, J.A.; Drake, J.L.; Levy, A.A.; Butterfield, D.A.; Buxton, J.W.; Snyder, J.C. Communication between the Maternal Testa and the Embryo and/or Endosperm Affect Testa Attributes in Tomato. Plant Physiol. 2003, 133, 145–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, K. il A BHLH Protein Partially Controls Proanthocyanidin and Phytomelanin Pigmentation in the Seed Coats of Morning Glory Ipomoea Tricolor. Hortic. Environ. Biotechnol. 2012, 53, 304–309. [Google Scholar] [CrossRef]
- Bellido, G.G.; Beta, T. Anthocyanin Composition and Oxygen Radical Scavenging Capacity (ORAC) of Milled and Pearled Purple, Black, and Common Barley. J. Agric. Food Chem. 2009, 57, 1022–1028. [Google Scholar] [CrossRef] [PubMed]
- Ji, Z.-J.; Wang, X.-G.; Zeng, Y.-X.; Ma, L.-Y.; Li, X.-M.; Liu, B.-X.; Yang, C.-D. Comparison of Physiological and Yield Traits between Purple- and White-Pericarp Rice Using SLs. Breed. Sci. 2012, 62, 71. [Google Scholar] [CrossRef] [Green Version]
- Gould, K.S. Nature’s Swiss Army Knife: The Diverse Protective Roles of Anthocyanins in Leaves. J. Biomed. Biotechnol. 2004, 2004, 314. [Google Scholar] [CrossRef] [Green Version]
- Guidi, L.; Brunetti, C.; Fini, A.; Agati, G.; Ferrini, F.; Gori, A.; Tattini, M. UV Radiation Promotes Flavonoid Biosynthesis, While Negatively Affecting the Biosynthesis and the de-Epoxidation of Xanthophylls: Consequence for Photoprotection? Environ. Exp. Bot. 2016, 127, 14–25. [Google Scholar] [CrossRef]
- Morgounov, A.; Karaduman, Y.; Akin, B.; Aydogan, S.; Baenziger, P.S.; Bhatta, M.; Chudinov, V.; Dreisigacker, S.; Govindan, V.; Güler, S.; et al. Yield and Quality in Purple-Grained Wheat Isogenic Lines. Agronomy 2020, 10, 86. [Google Scholar] [CrossRef] [Green Version]
- Weltzien, E. Evaluation of Barley (Hordeum vulgare L.) Landrace Populations Originating from Different Growing Regions in the Near East*. Plant Breed. 1988, 101, 95–106. [Google Scholar] [CrossRef] [Green Version]
- Choo, T.M.; Vigier, B.; Ho, K.M.; Ceccarelli, S.; Grando, S.; Franckowiak, J.D. Comparison of Black, Purple, and Yellow Barleys. Genet. Resour. Crop Evol. 2005, 52, 121–126. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait\Line | Bowman | BLP | PLP | BP |
|---|---|---|---|---|
| Field | ||||
| Plant height, cm | 75.15 ± 2.66 | 75.03 ± 0.80 | 76.38 ± 1.16 | 76.12 ± 1.74 |
| Spike number per plant | 3.63 ± 0.67 | 4.47 ± 0.48 | 3.93 ± 0.54 | 3.57 ± 0.58 |
| Spike length, cm | 5.25 ± 0.18 | 5.52 ± 0.06 | 5.7 ± 0.33 | 5.62 ± 0.28 |
| Spike density | 11.58 ± 0.55 ab | 12.53 ± 0.38 a | 11.66 ± 0.08 b | 11.93 ± 0.38 ab |
| Number of seeds per plant | 43.92 ± 6.92 | 58.87 ± 7.48 | 51.32 ± 7.09 | 47.28 ± 7.62 |
| Grain weight per plant, g | 2.17 ± 0.35 a | 3.05 ± 0.28 b | 2.42 ± 0.30 ab | 2.40 ± 0.39 ab |
| Thousand-grain weight, g | 49.43 ± 0.60 ad | 51.88 ± 2.08 ad | 47.22 ± 0.69 b | 50.82 ± 0.60 c |
| Plots | ||||
| Plant height, cm | 78.67 ± 1.94 ab | 76.47 ± 1.10 a | 80.78 ± 1.96 b | 79.99 ± 3.49 ab |
| Spike number per plant | 4.8 ± 0.40 | 4.95 ± 0.80 | 5.0 ± 0.52 | 5.66 ± 0.93 |
| Spike length, cm | 6.72 ± 0.25 a | 6.57 ± 0.08 a | 7.38 ± 0.16 bc | 7.45 ± 0.33 c |
| Spike density | 10.28 ± 0.18 | 10.47 ± 0.08 | 10.2 ± 0.40 | 10.42 ± 0.30 |
| Number of seeds per plant | 65.57 ± 4.64 | 65.85 ± 10.36 | 72.43 ± 11.45 | 76.41 ± 16.96 |
| Grain weight per plant, g | 3.57 ± 0.27 | 3.55 ± 0.61 | 3.90 ± 0.53 | 4.13 ± 0.92 |
| Thousand grain weight, g | 54.46 ± 0.64 | 53.8 ± 0.85 | 54.03 ± 1.95 | 55.04 ± 0.35 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Glagoleva, A.; Kukoeva, T.; Mursalimov, S.; Khlestkina, E.; Shoeva, O. Effects of Combining the Genes Controlling Anthocyanin and Melanin Synthesis in the Barley Grain on Pigment Accumulation and Plant Development. Agronomy 2022, 12, 112. https://doi.org/10.3390/agronomy12010112
Glagoleva A, Kukoeva T, Mursalimov S, Khlestkina E, Shoeva O. Effects of Combining the Genes Controlling Anthocyanin and Melanin Synthesis in the Barley Grain on Pigment Accumulation and Plant Development. Agronomy. 2022; 12(1):112. https://doi.org/10.3390/agronomy12010112
Chicago/Turabian StyleGlagoleva, Anastasiya, Tatjana Kukoeva, Sergey Mursalimov, Elena Khlestkina, and Olesya Shoeva. 2022. "Effects of Combining the Genes Controlling Anthocyanin and Melanin Synthesis in the Barley Grain on Pigment Accumulation and Plant Development" Agronomy 12, no. 1: 112. https://doi.org/10.3390/agronomy12010112
APA StyleGlagoleva, A., Kukoeva, T., Mursalimov, S., Khlestkina, E., & Shoeva, O. (2022). Effects of Combining the Genes Controlling Anthocyanin and Melanin Synthesis in the Barley Grain on Pigment Accumulation and Plant Development. Agronomy, 12(1), 112. https://doi.org/10.3390/agronomy12010112