
Allelopathy and Allelopathic Substances of Fossil Tree Species Metasequoia glyptostroboides


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Extraction
2.3. Determination of Allelopathic Activity
2.4. Separation of M. glyptostroboides Extract
2.5. Isolation of Allelopathic Substance from Fractions A and B
2.6. Allelopathic Activity of Compound 1 and 2
2.7. Statistical Analysis
3. Results
3.1. Allelopathic Activity of Fallen Leaves of M. Glyptostroboides
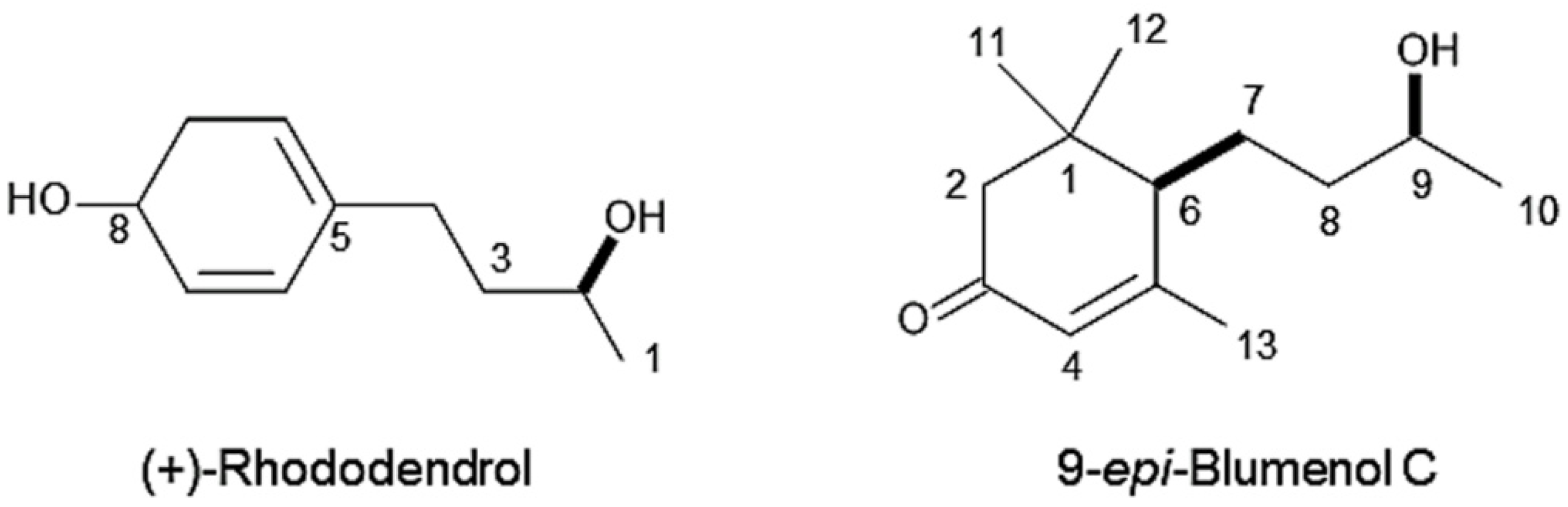
3.2. Identification of Allelopathic Substances
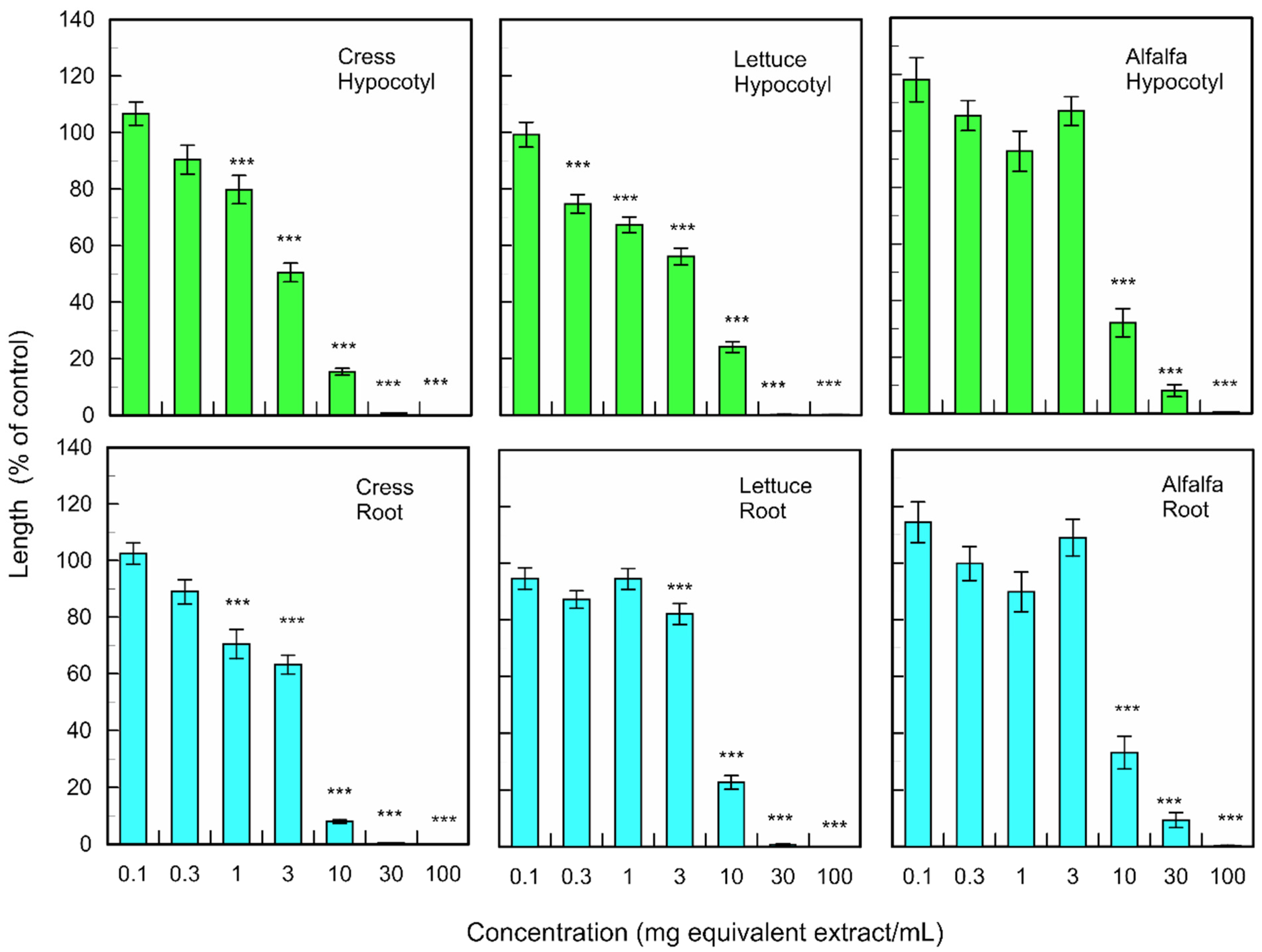
3.3. Allelopathic Activity of Isolated Compounds
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Miki, S. On the change of flora in Eastern Asia since Tertiary Period. J. Jpn. Bot. 1941, 11, 237–303. [Google Scholar]
- Hu, H.H. On the new family Metasequoiaceae and on Metasequoia glyptostroboides, a living species of the genus Metasequoia found in Szechuan and Hupeh. Bull Fan. Mem. Inst. Biol. Bot. 1948, 1, 153–161. [Google Scholar]
- Ma, J. The chronology of the ‘‘Living Fossil’’ Metasequoia glyptostroboides (Taxodiaceae): A review (1943–2003). Harv. Pap. Bot. 2003, 8, 9–18. [Google Scholar]
- Liu, Y.J.; Li, C.S.; Wang, Y.F. Studies on fossil Metasequoia from north-east China and their taxonomic implications. Bot. J. Linn. Soc. 1999, 130, 267–297. [Google Scholar] [CrossRef]
- Liu, Y.J.; Arens, N.C.; Li, C.S. Range change in Metasequoia: Relationship to palaeoclimate. Bot. J. Linn. Soc. 2007, 154, 115–127. [Google Scholar] [CrossRef]
- Florin, R. On Metasequoia, living and fossil. Bot. Notiser. 1952, 1, 1–29. [Google Scholar]
- Florin, R. The distribution of conifer and taxad genera in time and space. Acta Horti. Bergiani. 1963, 20, 121–326. [Google Scholar]
- Wu, M.; Yao, L.; Ai, X.; Zhu, J.; Zhu, Q.; Wang, J.; Huang, X.; Hong, J. The reproductive characteristics of core germplasm in a native Metasequoia glyptostroboides population. Biodivers. Sci. 2020, 28, 303–313. [Google Scholar] [CrossRef]
- Bate-Smith, E.C. Leuco-anthocyanins. 1. Detection and identification of anthocyanidins formed from leuco-anthocyanins in plant tissues. Biochem. J. 1954, 58, 122–125. [Google Scholar] [CrossRef] [Green Version]
- Hattori, S.; Yoshida, S.; Hasegawa, M. Occurrence of shikimic acid in the leaves of Gymnosperms. Physiol. Plant. 1954, 7, 283–289. [Google Scholar] [CrossRef]
- Juvik, O.J.; Nguyen, X.H.T.; Andersen, H.L.; Fossen, T. Growing with dinosaurs: Natural products from the Cretaceous relict Metasequoia glyptostroboides Hu & Cheng—A molecular reservoir from the ancient world with potential in modern medicine. Phytochem. Rev. 2016, 15, 161–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, S.; Umezawa, T. Biosynthesis of lignans and norlignans. J. Wood Sci. 2007, 53, 273–284. [Google Scholar] [CrossRef]
- Bajpai, V.K.; Rahman, A.; Kang, S.C. Chemical composition and anti-fungal properties of the essential oil and crude extracts of Metasequoia glyptostroboides Miki ex Hu. Ind. Crop Prod. 2007, 26, 28–35. [Google Scholar] [CrossRef]
- Bajpai, V.K.; Kang, S.C. Antifungal activity of leaf essential oil and extracts of Metasequoia glyptostroboides Miki ex Hu. J. Am. Oil Chem. Soc. 2010, 87, 327–336. [Google Scholar] [CrossRef]
- Asahi, M.; Sakan, F. Antifungal activity of diterpenes isolated from Metasequoia glyptostroboides Hu Et Cheng. J. Agr. Chem. Soc. Jpn. 1984, 58, 887–889. [Google Scholar]
- Bajpai, V.K.; Kang, S.C. Antimycotic potential of a diterpenoid taxoquinone against Candida species isolated from Metasequoia glyptostroboides. Bangl. J. Pharmacol. 2014, 9, 154–160. [Google Scholar] [CrossRef] [Green Version]
- Bajpai, V.K.; Kang, S.C. Antibacterial abietane-type diterpenoid, taxodone from Metasequoia glyptostroboides Miki ex Hu. J. Biosci. 2010, 35, 533–538. [Google Scholar] [CrossRef]
- Bajpai, V.K.; Kang, S.C. Isolation and characterization of biologically active secondary metabolites from Metasequoia glyptostroboides Miki Ex Hu. J. Food Safety 2011, 31, 276–283. [Google Scholar] [CrossRef]
- Nakatani, M.; Amano, K.; Shibata, K.; Komura, H.; Hase, T. Metaseol, a symmetrical diphenylmethane from Metasequoia glyptostroboides. Phytochemistry 1991, 30, 1034–1036. [Google Scholar] [CrossRef]
- Belz, R.G. Allelopathy in crop/weed interactions—An update. Pest. Manag. Sci. 2007, 63, 308–326. [Google Scholar] [CrossRef]
- Macías, F.A.; Molinillo, J.M.G.; Varela, R.M.; Galindo, J.G.G. Allelopathy—A natural alternative for weed control. Pest Manag. Sci. 2007, 63, 327–348. [Google Scholar] [CrossRef]
- Chengxu, W.; Mingxing, Z.; Xuhui, C.; Bo, Q. Review on allelopathy of exotic invasive plants. Procedia Engin. 2011, 18, 240–246. [Google Scholar] [CrossRef] [Green Version]
- Rice, E.L. Allelopathy, 2nd ed.; Academic Press: Orlando, FL, USA, 1984; 422p. [Google Scholar]
- Duke, S.O.; Dayan, F.E.; Romagni, J.G.; Rimando, A.M. Natural products as sources of herbicide, current status and future trends. Weed Res. 2000, 40, 99–111. [Google Scholar] [CrossRef]
- Cantrell, C.C.; Dayan, F.E.; Duke, O. Natural products as sources for new pesticide. J. Nat. Prod. 2012, 75, 1231–1242. [Google Scholar] [CrossRef] [PubMed]
- Pan, H.; Lundgren, L.N. Rhododendrol glycosides and phenyl glucoside esters from inner bark of Betula pubescens. Phytochemistry 1994, 36, 79–83. [Google Scholar] [CrossRef]
- Chattopadhyay, S.K.; Srivastava, S.; Sashidhara, K.V.; Tripathi, A.K.; Bhattacharya, A.K.; Negi, A.S. Betuligenol derivative with growth inhibition and antifeedant activity. Bioorg. Med. Chem. Lett. 2004, 14, 1729–1731. [Google Scholar] [CrossRef] [PubMed]
- Matsunami, K.; Otsuka, H.; Takeda, Y. Structural revisions of blumenol C glucoside and byzantionoside B. Chem. Pharm. Bull. 2010, 58, 438–441. [Google Scholar] [CrossRef] [Green Version]
- Harborne, J.B.; Williams, C.A. Leaf survey of flavonoids and simple phenols in the genus Rhododendron. Phytochemistry 1971, 10, 2727–2744. [Google Scholar] [CrossRef]
- Ito, S.; Wakamatsu, K. Biochemical mechanism of rhododendrol-induced leukoderma. Int. J. Mol. Sci. 2018, 19, 552. [Google Scholar] [CrossRef] [Green Version]
- Matsunaga, K.; Suzuki, K.; Ito, A.; Tanemura, A.; Abe, Y.; Suzuki, T.; Yoshikawa, M.; Sumikawa, Y.; Yagami, A.; Masui, Y.; et al. Rhododendrol-induced leukoderma update I: Clinical findings and treatment. J. Dermatol. 2021, 48, 961–968. [Google Scholar] [CrossRef]
- Das, K.R.; Iwasaki, K.; Suenaga, K.; Kato-Noguchi, H. Isolation and identification of two phytotoxic compounds from the medicinal plant Cassia alata L. Weed Biol. Manag. 2020, 20, 3–11. [Google Scholar] [CrossRef]
- Galbraith, M.N.; Horn, D.H.S. Structures of the natural products blumenols A, B, and C. J. Chem. Soc. Chem. Commun. 1972, 1972, 113–114. [Google Scholar] [CrossRef]
- Sun, G.; Putkaradze, N.; Bohnacker, S.; Jonczyk, R.; Fida, T.; Hoffmann, T.; Bernhardt, R.; Härtl, K.; Schwab, W. Six uridine-diphosphate glycosyltransferases catalyze the glycosylation of bioactive C13-apocarotenols. Plant Physiol. 2020, 184, 1744–1761. [Google Scholar] [CrossRef]
- Ohmiya, A.; Kishimoto, S.; Aida, R.; Yoshioka, S.; Sumitomo, K. Carotenoid cleavage dioxygenase (CmCCD4a) contributes to white color formation in chrysanthemum petals. Plant Physiol. 2006, 142, 1193–1201. [Google Scholar] [CrossRef] [Green Version]
- Walter, M.H.; Floss, D.S.; Strack, D. Apocarotenoids: Hormones, mycorrhizal metabolites and aroma volatiles. Planta 2010, 232, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Nisar, N.; Li, L.; Lu, S.; Khin, N.C.; Pogson, B.J. Carotenoid metabolism in plants. Mol. Plant 2015, 8, 68–82. [Google Scholar] [CrossRef] [Green Version]
- Hou, X.; Rivers, J.; León, P.; McQuinn, R.P.; Pogson, B.J. Synthesis and function of apocarotenoid signals in plants. Trends Plant Sci. 2016, 21, 792–803. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Takano, Y.; Matsuura, H.; Yoshihara, T. Antifungal compounds from the root and root exudate of Zea mays. Biosci. Biotechnol. Biochem. 2004, 68, 1366–1368. [Google Scholar] [CrossRef] [Green Version]
- Walter, M.H.; Strack, D. Carotenoids and their cleavage products: Biosynthesis and functions. Nat. Prod. Rep. 2011, 28, 663–692. [Google Scholar] [CrossRef]
- Maier, W.; Schmidt, J.; Nimtz, M.; Wray, V.; Strack, D. Secondary products in mycorrhizal roots of tobacco and tomato. Phytochemistry 2000, 54, 473–479. [Google Scholar] [CrossRef]
- Wang, M.; Schäfer, M.; Li, D.; Halitschke, R.; Dong, C.; McGale, E.; Paetz, C.; Song, Y.; Li, S.; Dong, J.; et al. Blumenols as shoot markers of root symbiosis with arbuscular mycorrhizal fungi. eLife 2018, 7, e37093. [Google Scholar] [CrossRef] [PubMed]
- Adolfsson, L.; Nziengui, H.; Abreu, I.N.; Šimura, J.; Beebo, A.; Herdean, A.; Aboalizadeh, J.; Široká, J.; Moritz, T.; Novák, O.; et al. Enhanced secondary- and hormone metabolism in leaves of arbuscular mycorrhizal Medicago truncatula. Plant Physiol. 2017, 175, 392–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brundrett, M.C.; Tedersoo, L. Evolutionary history of mycorrhizal symbioses and global host plant diversity. New Phytologist. 2018, 220, 1108–1115. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis, 3rd ed.; Academic Press: London, UK, 2008; 815p. [Google Scholar]
- Luginbuehl, L.H.; Menard, G.N.; Kurup, S.; Van Erp, H.; Radhakrishnan, G.V.; Breakspear, A.; Oldroyd, G.E.D.; Eastmond, P.J. Fatty acids in arbuscular mycorrhizal fungi are synthesized by the host plant. Science 2017, 356, 1175–1178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinzone, P.; Potts, D.; Pettibone, G.; Warren, R. Do novel weapons that degrade mycorrhizal mutualisms promote species invasion? Plant Ecol. 2018, 219, 539–548. [Google Scholar] [CrossRef]
- Mallik, A.U. Allelopathy and competition in coniferous forests. Environ. For. Sci. 1998, 54, 309–315. [Google Scholar] [CrossRef]
- Kato-Noguchi, H.; Takeshita, S.; Kimura, F.; Ohno, O.; Suenaga, K. A novel allelopathic active substance in Ginkgo biloba. J. Plant Physiol. 2013, 170, 1595–1599. [Google Scholar] [CrossRef]
- Kato-Noguchi, H.; Takeshita, S. Contribution of a phytotoxic compound to the allelopathy of Ginkgo biloba. Plant Signal. Behav. 2013, 8, e26999. [Google Scholar] [CrossRef] [Green Version]
- Kato-Noguchi, H.; Kimura, F.; Ohno, O.; Suenaga, K. Involvement of allelopathy in inhibition of understory growth in red pine forests. J. Plant Physiol. 2017, 218, 66–73. [Google Scholar] [CrossRef]
- Kato-Noguchi, H.; Kurniadie, D. Allelopathy and allelopathic substances of mango (Mangifera indica L.). Weed Biol. Manag. 2020, 20, 131–138. [Google Scholar] [CrossRef]
- Kato-Noguchi, H. Phytotoxic substances involved in teak allelopathy and agroforestry. Appl. Sci. 2021, 11, 3314. [Google Scholar] [CrossRef]
- Kato-Noguchi, H.; Kurniadie, D. Allelopathy of Lantana camara as an Invasive plant. Plants 2021, 10, 1028. [Google Scholar] [CrossRef] [PubMed]
- Bais, H.P.; Weir, T.L.; Perry, L.G.; Gilroy, S.; Vivanco, J.M. The role of root exudates in rhizosphere interactions with plants and other organisms. Annu. Rev. Plant Biol. 2006, 57, 233–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonanomi, G.; Sicurezza, M.G.; Caporaso, S.; Esposito, A.; Mazzoleni, S. Phytotoxicity dynamics of decaying plant materials. New Phytol. 2006, 169, 571–578. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Root | Hypocotyl/Coleoptil |
|---|---|---|
| Cress | 7.1 | 6.8 |
| Lettuce | 3.9 | 2.2 |
| Alfalfa | 7.2 | 7.4 |
| Lolium multiflorum | 10.6 | 7.6 |
| Phleum pretense | 12.4 | 8.1 |
| Vulpia myuros | 2.1 | 4.8 |
| (+)-Rhododendrol | 9-epi-Blumenol C | |||
|---|---|---|---|---|
| Species | Root | Hypocotyl | Root | Coleoptil |
| Cress | 9.6 | 10.6 | 2780 | 2240 |
| Lolium multiflorum | 23.1 | 7.2 | 3290 | 2950 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matuda, Y.; Iwasaki, A.; Suenaga, K.; Kato-Noguchi, H. Allelopathy and Allelopathic Substances of Fossil Tree Species Metasequoia glyptostroboides. Agronomy 2022, 12, 83. https://doi.org/10.3390/agronomy12010083
Matuda Y, Iwasaki A, Suenaga K, Kato-Noguchi H. Allelopathy and Allelopathic Substances of Fossil Tree Species Metasequoia glyptostroboides. Agronomy. 2022; 12(1):83. https://doi.org/10.3390/agronomy12010083
Chicago/Turabian StyleMatuda, Yuki, Arihiro Iwasaki, Kiyotake Suenaga, and Hisashi Kato-Noguchi. 2022. "Allelopathy and Allelopathic Substances of Fossil Tree Species Metasequoia glyptostroboides" Agronomy 12, no. 1: 83. https://doi.org/10.3390/agronomy12010083
APA StyleMatuda, Y., Iwasaki, A., Suenaga, K., & Kato-Noguchi, H. (2022). Allelopathy and Allelopathic Substances of Fossil Tree Species Metasequoia glyptostroboides. Agronomy, 12(1), 83. https://doi.org/10.3390/agronomy12010083