
Salt-Stressed Coriander (Coriandrum sativum L.) Responses to Potassium Silicate, Humic Acid and Gamma Irradiation Pretreatments

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Seeds, Chemicals, and Physiological Treatments
2.2. Sowing, Irrigation, and Sampling
2.3. Extraction, Isolation, and Estimation of Plant Growth Regulators
2.4. Estimation of Photosynthetic Pigments
2.5. Carbohydrate Content Estimation
2.6. Extraction and Measurement of Antioxidant Defense Compounds
2.7. Extraction and Estimation of Antioxidant Enzymes
2.8. Lipid Peroxidation Estimation
2.9. Total Cellular Proteins (TCPs) Extraction and Monitoring the Expression of RuBisCOLS
2.10. Statistical Analysis
3. Results
3.1. Growth and Yield Parameters
3.2. Changes in Phytohormones
3.3. Changes in Photosynthetic Pigments and Carbohydrate Contents
3.4. Changes in Antioxidant Compounds
3.5. Changes in the Antioxidant Enzyme Pool and MDA
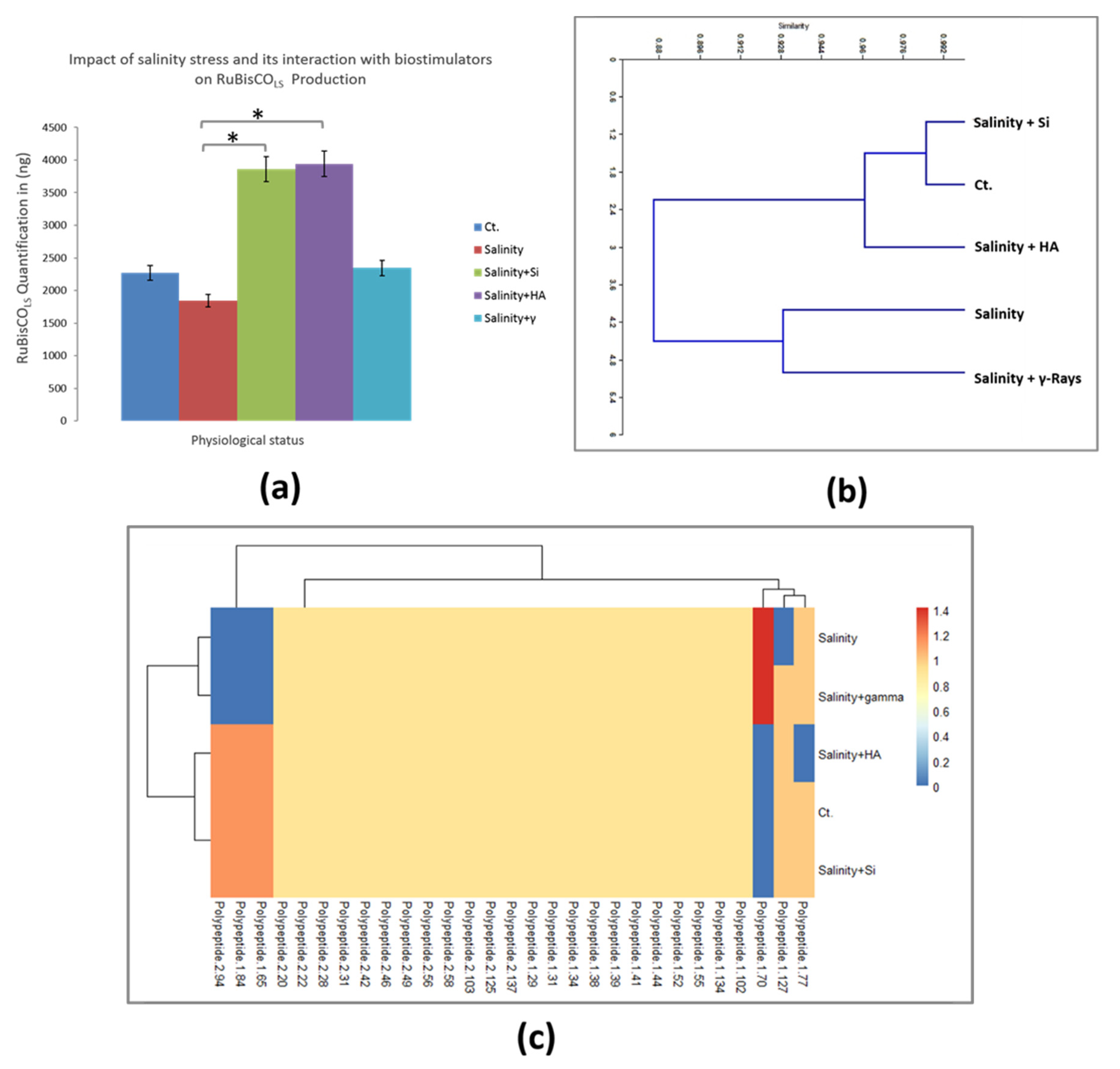
3.6. Characterization of Salinity Stress Impact on TCPs and the Expression of RuBisCOLS
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Aiazzi, M.T.; Di Rienzio, J.A.; Sosa, L. Effects of different salts on the germination and early seedling growth of Atriplex cordobensis Gandoger et Stuckert (Chenopodiaceae). Seed Sci. Technol. 2009, 37, 1. [Google Scholar] [CrossRef]
- Hong, Y.; Liu, Y.; Zhang, Y.; Jia, L.; Yang, X.; Zhang, X.; Liu, J.; Luan, Y. Genome-wide characterization of homeobox-leucine zipper gene family in tomato (Solanum lycopersicum) and functional analysis of SlHDZ34 (III sub-family member) under salinity stress. Environ. Exp. Bot. 2021, 192, 104652. [Google Scholar] [CrossRef]
- Meng, S.; Su, L.; Li, Y.; Wang, Y.; Zhang, C.; Zhao, Z. Nitrate and ammonium contribute to the distinct nitrogen metabolism of populus simonii during moderate salt stress. PLoS ONE 2016, 11, e0150354. [Google Scholar] [CrossRef]
- Abdel Latef, A.A.H.; Omer, A.M.; Badawy, A.A.; Osman, M.S.; Ragaey, M.M. Strategy of Salt Tolerance and Interactive Impact of Azotobacter chroococcum and/or Alcaligenes faecalis Inoculation on Canola (Brassica napus L.) Plants Grown in Saline Soil. Plants 2021, 10, 110. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Dubey, R.S. Involvement of oxidative stress and role of antioxidative defense system in growing rice seedlings exposed to toxic concentrations of aluminum. Plant Cell Rep. 2007, 26, 11. [Google Scholar] [CrossRef] [PubMed]
- Widodo, J.; Patterson, H.; Newbigin, E.; Tester, M.; Bacic, A.; Roessner, U. Metabolic responses to salt stress of barley (Hordeum vulgare L.) cultivars, Sahara and Clipper, which differ in salinity tolerance. J. Exp. Bot. 2009, 60, 14. [Google Scholar] [CrossRef] [PubMed]
- Nadeem, M.; Anjum, F.M.; Khan, M.I.; Tehseen, S.; El-Ghorab, A.; Sultan, J.I. Nutritional and medicinal aspects of coriander (Coriandrum sativum L.): A review. Br. Food J. 2013, 115, 5. [Google Scholar] [CrossRef]
- Shilev, S. Plant-growth-promoting bacteria mitigating soil salinity stress in plants. Appl. Sci. 2020, 10, 7326. [Google Scholar] [CrossRef]
- Rizwan, M.; Ali, S.; Ibrahim, M.; Farid, M.; Adrees, M.; Bharwana, S.A.; Zia-Ur-Rehman, M.; Qayyum, M.F.; Abbas, F. Mechanisms of silicon-mediated alleviation of drought and salt stress in plants: A review. Environ. Sci. Pollut. Res. 2015, 22, 20. [Google Scholar] [CrossRef]
- Khan, W.U.D.; Aziz, T.; Maqsood, M.A.; Sabir, M.; Ahmad, H.R.; Ramzani, P.M.A.; Naseem, M. Silicon: A Beneficial Nutrient Under Salt Stress, Its Uptake Mechanism and Mode of Action. In Soil Science: Agricultural and Environmental Prospectives; Hakeem, K., Akhtar, J., Sabir, M., Eds.; Springer: Cham, Switzerland, 2016. [Google Scholar] [CrossRef]
- Almeida, D.M.; Margarida Oliveira, M.; Saibo, N.J.M. Regulation of Na+ and K+ homeostasis in plants: Towards improved salt stress tolerance in crop plants. Genet. Mol. Biol. 2017, 40, 1. [Google Scholar] [CrossRef]
- Pesaran, A.A. Moisture Transport in Silica Gel Particle Beds; University of California: Los Angeles, CA, USA, 1983. [Google Scholar]
- El-Serafy, R.S.; El-Sheshtawy, A.N.A.; Atteya, A.K.G.; Al-Hashimi, A.; Abbasi, A.M.; Al-Ashkar, I. Seed priming with silicon as a potential to increase salt stress tolerance in lathyrus odoratus. Plants 2021, 10, 2140. [Google Scholar] [CrossRef] [PubMed]
- Çimrin, K.M.; Türkmen, Ö.; Turan, M.; Tuncer, B. Phosphorus and humic acid application alleviate salinity stress of pepper seedling. Afr. J. Biotechnol. 2010, 9, 36. [Google Scholar] [CrossRef]
- Mackowiak, C.L.; Grossl, P.R.; Bugbee, B.G. Beneficial Effects of Humic Acid on Micronutrient Availability to Wheat. Soil Sci. Soc. Am. J. 2001, 65, 6. [Google Scholar] [CrossRef] [PubMed]
- Canellas, L.P.; Olivares, F.L.; Okorokova-Façanha, A.L.; Façanha, A.R. Humic acids isolated from earthworm compost enhance root elongation, lateral root emergence, and plasma membrane H+-ATPase activity in maize roots. Plant Physiol. 2002, 130, 4. [Google Scholar] [CrossRef]
- Chen, Y.; Magen, H.; Clapp, C.E. Mechanisms of plant growth stimulation by humic substances: The role of organo-iron complexes. Soil Sci. Plant Nutr. 2004, 50, 7. [Google Scholar] [CrossRef]
- Delfine, S.; Tognetti, R.; Desiderio, E.; Alvino, A. Effect of foliar application of N and humic acids on growth and yield of durum wheat. Agronomie 2005, 25, 2. [Google Scholar] [CrossRef]
- Türkmen, O.; Demr, S.; Ensoy, S.; Dursum, A. Effects of Arbuscular Mycorrhizal Fungus and Humic Acid on the Seedling Development and Nutrient Content of Pepper Grown under Saline Soil Conditions. J. Biol. Sci. 2005, 5, 5. [Google Scholar] [CrossRef]
- Aydin, A.; Kant, C.; Turan, M. Humic acid application alleviate salinity stress of bean (Phaseolus vulgaris L.) plants decreasing membrane leakage. Afr. J. Agric. Res. 2012, 7, 7. [Google Scholar] [CrossRef]
- Qi, W.; Zhang, L.; Xu, H.; Wang, L.; Jiao, Z. Physiological and molecular characterization of the enhanced salt tolerance induced by low-dose gamma irradiation in Arabidopsis seedlings. Biochem. Biophys. Res. Commun. 2014, 450, 2. [Google Scholar] [CrossRef]
- Beyaz, R.; Yildiz, M. The Use of Gamma Irradiation in Plant Mutation Breeding. In Plant Engineering; Reed Business Information: Singapore, 2017. [Google Scholar] [CrossRef]
- Beyaz, R. Impact of gamma irradiation pretreatment on the growth of common vetch (Vicia sativa L.) seedlings grown under salt and drought stress. Int. J. Radiat. Biol. 2020, 96, 2. [Google Scholar] [CrossRef]
- Elhamahmy, A.M.; Elsadany, O.; Eid, M.; Abdelazeem, S.; Gerish, S.; Kalaji, M.H.; Wróbel, J.; Elsheery, N. Impact of gamma irradiation pretreatment on biochemical and molecular responses of potato growing under salt stress. Chem. Biol. Technol. Agric. 2021, 8, 35. [Google Scholar] [CrossRef]
- Shindy, W.W.; Smith, O.E. Identification of Plant Hormones from Cotton Ovules. Plant Physiol. 1975, 55, 3. [Google Scholar] [CrossRef]
- Hassanein, R.A.; Hussein, O.S.; Abdelkader, A.F.; Farag, I.A.; Hassan, Y.E.; Ibrahim, M. Metabolic activities and molecular investigations of the ameliorative impact of some growth biostimulators on chilling-stressed coriander (Coriandrum sativum L.) plant. BMC Plant Biol. 2021, 21, 1. [Google Scholar] [CrossRef]
- Kelen, M.; Demiralay, E.Ç.; Şen, S.; Alsancak, G.Ö. Separation of abscicic acid, indole-3-acetic acid, gibberellic acid in 99R (vitis berlandieri x vitis rupestris) and rose oil (Rosa damascene Mill.) reversed phase liquid chromatography. Turk. J. Chem. 2004, 28, 603–610. [Google Scholar]
- Metzner, H.; Rau, H.A.; Senger, H. Untersuchungen zur Synchronisierbarkeit einzelner Pigmentmangel-Mutanten von Chlorella. Planta 2004, 65, 186–194. [Google Scholar] [CrossRef]
- Homme, P.M.; Gonalez, B.; Billard, L. Carbohydrate content, fructane and sucroce enzyme activities in roots, stubble and leaves of rye grass (Lolium perenne L.) as affected by source/sink modification after cutting. J. Plant Physiol. 1992, 140, 282–291. [Google Scholar] [CrossRef]
- Whistler, R.L.; Wolform, M.L.; Bemiller, J.N.; Shafizadeh, F. Anthrone coluorimetric method. In Methods in Carbohydrate Chemistry; Acadimic Press: New York, NY, USA; London, UK, 1962; Volume 1, p. 384. [Google Scholar]
- Hedge, J.E.; Hofreiter, B.T. Anthrone coluorimetric method. In Methods in Carbohydrate Chemistry; Academic Press: New York, NY, USA, 1962; Volume 1, pp. 380–394. [Google Scholar]
- Zvaigzne, G.; Karklina, D.; Seglina, D.; Krasnova, I. Antioxidant in various citrus fruits juices. Chemine Technol. 2009, 3, 56–61. [Google Scholar]
- Marinova, D.; Ribarova, F.; Atanassova, M. Total phenolic and total flavonoids in Bulgarian fruits and vegetables. J. Univ. Chem. Technol. Metall. 2005, 40, 255–260. [Google Scholar]
- Malik, C.P.; Singh, S.M. Plant Enzymology and Histoenzymology; Kalyani Puplishers: New Delhi, India, 1980; Volume 53, p. 286. [Google Scholar]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 1. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 1. [Google Scholar] [CrossRef]
- Aebi, H. Methods of Enzymatic Analysis. Catalase; Bergmeyer, H.U., Ed.; Verlag Chemie/Academic Press Inc.: Weinheim, Germany; NewYork, NY, USA, 1974. [Google Scholar]
- Hammerschmidt, R.; Nuckles, E.M.; Kuć, J. Association of enhanced peroxidase activity with induced systemic resistance of cucumber to Colletotrichum lagenarium. Physiol. Plant Pathol. 1982, 20, 1. [Google Scholar] [CrossRef]
- Oktay, M.; Kufrevioğlu, O.I.; Kocacaliskan, I.; Sakiroğlu, H. Polyphenol oxidase from Amasya apple. J. Food Sci 1995, 60, 495–499. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts. I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 1. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 5259. [Google Scholar] [CrossRef] [PubMed]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. Past: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Snedecor, G.W.; Cochran, W.G. Statistical Methods, 7th ed.; Iowa State University Press: Ames, IA, USA, 1982. [Google Scholar]
- Kassambara, A.; Mundt, F. Factoextra: Extract and Visualize the Results of Multivariate Data Analyses, R package version 1.0.7. Available online: https://CRAN.R-project.org/package=factoextra (accessed on 31 March 2021).
- Suzuki, R.; Shimodaira, H. Hierarchical Clustering with P-Values via Multiscale Bootstrap Resampling, R package. 2006. Available online: http://www.is.titech.ac.jp/~shimo/prog/pvclust/ (accessed on 31 March 2021).
- Igbari, A.; Nodza, G.; Adeusi, A.; Ogundipe, O. Morphological characterization of mango (Mangifera indica L.) cultivars from south-west Nigeria. Ife J. Sci. 2019, 21, 155–163. [Google Scholar] [CrossRef]
- Soetewey, A. Correlation Coeffificient and Correlation Test in R. Available online: https://www.statsandr.com/blog/correlation-coefficient-and-correlation-test-in-r/ (accessed on 20 February 2021).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Kassambara, A. ggpubr: “ggplot2” Based Publication Ready Plots, Version R Package Version 0.3. 0; Github: San Francisco, CA, USA, 2020.
- Negrão, S.; Schmöckel, S.M.; Tester, M. Evaluating physiological responses of plants to salinity stress. Ann. Bot. 2017, 119, 1. [Google Scholar] [CrossRef]
- Al-Ashkar, I.; Alderfasi, A.; Ben Romdhane, W.; Seleiman, M.F.; El-Said, R.A.; Al-Doss, A. Morphological and genetic diversity within salt tolerance detection in eighteen wheat genotypes. Plants 2020, 9, 287. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.H.; Zhou, D.; Cui, J.J.; Ma, H.L.; Lang, D.Y.; Wu, X.L.; Wang, Z.S.; Qiu, H.Y.; Li, M. Effect of silicon on seed germination and the physiological characteristics of glycyrrhiza uralensis under different levels of salinity. J. Hortic. Sci. Biotechnol. 2015, 90, 4. [Google Scholar] [CrossRef]
- Soltabayeva, A.; Ongaltay, A.; Omondi, J.O.; Srivastava, S. Morphological, physiological and molecular markers for salt-stressed plants. Plants 2021, 10, 243. [Google Scholar] [CrossRef]
- Fraire-Velazquez, S.; Emmanuel, V. Abiotic Stress in Plants and Metabolic Responses. In Abiotic Stress—Plant Responses and Applications in Agriculture; IntechOpen: London, UK, 2013. [Google Scholar] [CrossRef] [Green Version]
- Khaled, H.; Fawy, H. Effect of different Levels of humic acids on the nutrient content, plant growth, and soil properties under conditions of salinity. Soil Water Res. 2011, 6, 1. [Google Scholar] [CrossRef]
- Kaya, C.; Akram, N.A.; Ashraf, M.; Sonmez, O. Exogenous application of humic acid mitigates salinity stress in maize (Zea mays L.) plants by improving some key physico-biochemical attributes. Cereal Res. Commun. 2018, 46, 1. [Google Scholar] [CrossRef]
- El Mongy, M.S.; Zhou, H.; Cao, Y.; Liu, B.; Xia, Y. The effect of humic acid on endogenous hormone levels and antioxidant enzyme activity during in vitro rooting of evergreen azalea. Sci. Hortic. 2018, 227, 234–243. [Google Scholar] [CrossRef]
- Rivero, R.M.; Mestre, T.C.; Mittler, R.; Rubio, F.; Garcia-Sanchez, F.; Martinez, V. The combined effect of salinity and heat reveals a specific physiological, biochemical and molecular response in tomato plants. Plant Cell Environ. 2014, 37, 5. [Google Scholar] [CrossRef] [PubMed]
- Omer, A.M.; Osman, M.S.; Badawy, A.A. Inoculation with Azospirillum brasilense and/or Pseudomonas geniculata reinforces flax (Linum usitatissimum) growth by improving physiological activities under saline soil conditions. Bot. Stud. 2022, 63, 15. [Google Scholar] [CrossRef] [PubMed]
- Adam, S.; Murthy, S. Characterization of alterations in photosynthetic electron transport activities in maize thylakoid membranes under zinc stress. Eur. J. Exp. Biol. 2014, 4, 25–29. [Google Scholar]
- Mittler, R. ROS Are Good. Trends Plant Sci. 2017, 22, 1. [Google Scholar] [CrossRef] [PubMed]
- Szymańska, R.; Ślesak, I.; Orzechowska, A.; Kruk, J. Physiological and biochemical responses to high light and temperature stress in plants. Environ. Exp. Bot. 2017, 139, 165–177. [Google Scholar] [CrossRef]
- Naz, H.; Akram, N.A.; Ashraf, M. Impact of ascorbic acid on growth and some physiological attributes of cucumber (Cucumis Sativus) plants under water-deficit conditions. Pak. J. Bot. 2016, 48, 877–883. [Google Scholar]
- Latif, M.; Akram, N.A.; Ashraf, M. Regulation of some biochemical attributes in drought-stressed cauliflower (Brassica oleracea L.) by seed pre-treatment with ascorbic acid. J. Hortic. Sci. Biotechnol. 2016, 91, 2. [Google Scholar] [CrossRef]
- Cvikrová, M.; Gemperlová, L.; Dobrá, J.; Martincová, O.; Prásil, I.T.; Gubis, J.; Vanková, R. Effect of heat stress on polyamine metabolism in proline-over-producing tobacco plants. Plant Sci. 2012, 182, 1. [Google Scholar] [CrossRef]
- Badawy, A.A.; Alotaibi, M.O.; Abdelaziz, A.M.; Osman, M.S.; Khalil, A.M.A.; Saleh, A.M.; Mohammed, A.E.; Hashem, A.H. Enhancement of Seawater Stress Tolerance in Barley by the Endophytic Fungus Aspergillus ochraceus. Metabolites 2021, 11, 428. [Google Scholar] [CrossRef]
- Alshaal, T.; Alsaeedi, A.; El-Ramady, H.; Almohsen, M. Enhancing seed germination and seedlings development of common bean (Phaseolus vulgaris) by SiO2 nanoparticles. Egypt J. Soil Sci. 2017, 57, 407–415. [Google Scholar] [CrossRef]
- Ahanger, M.A.; Alyemeni, M.N.; Wijaya, L.; Alamri, S.A.; Alam, P.; Ashraf, M.; Ahmad, P. Potential of exogenously sourced kinetin in protecting Solanum lycopersicum from NaCl-induced oxidative stress through up-regulation of the antioxidant system, ascorbate-glutathione cycle and glyoxalase system. PLoS ONE 2018, 13, e0202175. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, P.; Ahanger, M.A.; Alam, P.; Alyemeni, M.N.; Wijaya, L.; Ali, S.; Ashraf, M. Silicon (Si) Supplementation Alleviates NaCl Toxicity in Mung Bean [Vigna radiata (L.) Wilczek] Through the Modifications of Physio-biochemical Attributes and Key Antioxidant Enzymes. J. Plant Growth Regul. 2019, 38, 1. [Google Scholar] [CrossRef]
- Coskun, D.; Britto, D.T.; Huynh, W.Q.; Kronzucker, H.J. The role of silicon in higher plants under salinity and drought stress. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef]
- Geng, X.M.; Liu, X.; Ji, M.; Hoffmann, W.A.; Grunden, A.; Xiang, Q.Y.J. Enhancing heat tolerance of the little dogwood cornus canadensis L. f. with introduction of a superoxide reductase gene from the hyperthermophilic archaeon Pyrococcus furiosus. Front. Plant Sci. 2016, 7, 26. [Google Scholar] [CrossRef]
- Shereen, A.; Ansari, R.; Mumtaz, S.; Bughio, H.R.; Mujtaba, S.M.; Shirazi, M.U.; Khan, M.A. Impact of Gamma irradiation induced changes on growth and physiological responses of rice under saline conditions. Pak. J. Bot. 2009, 41, 2487–2495. [Google Scholar]
- Wi, S.G.; Chung, B.Y.; Kim, J.S.; Kim, J.H.; Baek, M.H.; Lee, J.W.; Kim, Y.S. Effects of gamma irradiation on morphological changes and biological responses in plants. Micron 2007, 38, 553–564. [Google Scholar] [CrossRef]
- Geng, X.; Zhang, Y.; Wang, L.; Yang, X. Pretreatment with high-dose gamma irradiation on seeds enhances the tolerance of sweet osmanthus seedlings to salinity stress. Forests 2019, 10, 406. [Google Scholar] [CrossRef]
- Krapp, A.; Hofmann, B.; Schäfer, C.; Stitt, M. Regulation of the expression of rbcS and other photosynthetic genes by carbohydrates: A mechanism for the ‘sink regulation’ of photosynthesis? Plant J. 1993, 3, 6. [Google Scholar] [CrossRef]
- Kim, D.W.; Rakwal, R.; Agrawal, G.K.; Jung, Y.H.; Shibato, J.; Jwa, N.S.; Iwahashi, Y.; Iwahashi, H.; Kim, D.H.; Shim, I.S. A hydroponic rice seedling culture model system for investigating proteome of salt stress in rice leaf. Electrophoresis 2005, 26, 4521–4539. [Google Scholar] [CrossRef]
- Parker, R.; Flowers, T.J.; Moore, A.L.; Harpham, N.V.J. An accurate and reproducible method for proteome profiling of the effects of salt stress in the rice leaf lamina. J. Exp. Bot. 2006, 57, 5. [Google Scholar] [CrossRef]
- Garg, N.; Singla, R. Growth, photosynthesis, nodule nitrogen and carbon fixation in the chickpea cultivars under salt stress. Braz. J. Plant Physiol. 2004, 16, 3. [Google Scholar] [CrossRef]
- Desingh, R.; Kanagaraj, G. Influence of salinity stress on photosynthesis and antioxidative systems. Gen. Appl. Plant Physiol. 2007, 33, 221–234. [Google Scholar]
- Ratnakar, A.; Rai, A. Influence of Nacl Salinity On The Activity Of Ribulose 1, 5-Bisphosphate Carboxylase In Three Leafy Vegetables. Octa J. Environ. Res. 2015, 3, 163–166. [Google Scholar]
- Sharkey, T.D. Effects of moderate heat stress on photosynthesis: Importance of thylakoid reactions, rubisco deactivation, reactive oxygen species, and thermotolerance provided by isoprene. Plant Cell Environ. 2005, 28, 3. [Google Scholar] [CrossRef]
- Metodiev, M.V.; Tsonev, T.D.; Popova, L.P. Effect of jasmonic acid on the stomatal and nonstomatal limitation of leaf photosynthesis in barley leaves. J. Plant Growth Regul. 1996, 15, 2. [Google Scholar] [CrossRef]
- Maslenkova, L.T.; Miteva, T.S.; Popova, L.P. Changes in the polypeptide patterns of barley seedlings exposed to jasmonic acid and salinity. Plant Physiol. 1992, 98, 2. [Google Scholar] [CrossRef]
- Hassan, N.M.; Serag, M.S.; El Feky, F.M. Changes in nitrogen content and protein profile following in vitro selection of NaCl resistant mung bean and tomato. Acta. Physiol. Plant. 2004, 26, 165–175. [Google Scholar] [CrossRef]
- Zhang, L.; Gao, M.; Zhang, L.; Li, B.; Han, M.; Alva, A.K.; Ashraf, M. Role of exogenous glycinebetaine and humic acid in mitigating drought stress-induced adverse effects in Malus robusta seedlings. Turk. J. Bot. 2013, 37, 5. [Google Scholar] [CrossRef]
- Manivannan, A.; Soundararajan, P.; Muneer, S.; Ko, C.H.; Jeong, B.R. Silicon mitigates salinity stress by regulating the physiology, antioxidant enzyme activities, and protein expression in Capsicum annuum ‘Bugwang’. Biomed. Res. Int. 2016, 2016, 3076357. [Google Scholar] [CrossRef]
- Khalifa, N.; Shehata, M.; Sakran, A.; Shehata, M.; Abdelsattar, M. Differential Expression of RuBisCO large Subunit and its Assembly in Intact Chloroplasts of Pisum sativum L. Treated with NaCl and Jasmonic Acid. Egypt. J. Bot. 2017, 57, 525–537. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| No. | Treatment | Irrigation |
|---|---|---|
| 1 | Control | Tap water |
| 2 | Salinity (150 mM NaCl) | Saline solution (150 mM NaCl) |
| 3 | Pot. silicate (80 mM) | Tap water |
| 4 | Humic acid (50 mg L−1) | Tap water |
| 5 | γ-rays (50 Gy) | Tap water |
| 6 | NaCl (150 mM) + Pot. silicate | Saline solution (150 mM NaCl) |
| 7 | NaCl (150 mM) + Humic acid | Saline solution (150 mM NaCl) |
| 8 | NaCl (150 mM) + γ-rays | Saline solution (150 mM NaCl) |
| Growth Parameters | Shoot Length (cm) | No. of Leaves/Plants | No. of Branches/ Plant | Leaves Area/ Plant (cm)2 | Fresh Weight of Shoot (g) | Dry Weight of Shoot (g) | Root Length (cm) | Fresh Weight of Root (g) | Dry Weight of Root (g) | |
|---|---|---|---|---|---|---|---|---|---|---|
| Treatment | ||||||||||
| Control | 11.27 ± 0.1 d | 16 ± 1.53 d | 5 ± 0.58 d | 30.2 ± 1.67 e | 0.59 ± 0.044 de | 0.09 ± 0.001 f | 6.17 ± 0.55 cd | 0.043 ± 0.004 g | 0.011 ± 0.00 d | |
| Salinity (150 mM NaCl) | 9.83 ± 0.21 e | 12 ± 1.00 d | 3.33 ± 0.58 e | 17.18 ± 0.84 f | 0.46 ± 0.006 e | 0.06 ± 0.31 g | 5.97 ± 0.31 d | 0.058 ± 0.01 f | 0.013 ± 0.001 d | |
| pot. Silicate (80 mM) | 17.07 ± 0.89 a | 27.67 ± 1.00 b | 6 ± 0.58 abcd | 93.94 ± 4.38 b | 0.77 ± 0.065 cde | 0.14 ± 0.64 d | 6.83 ± 0.64 abc | 0.061 ± 0.009 e | 0.015 ± 0.001 c | |
| Humic acid (50 mg−1 L) | 17.77 ± 1.58 a | 33.33 ± 1.00 a | 6.67 ± 0.58 ab | 107.8 ± 1.18 a | 1.17 ± 0.013 b | 0.19 ± 0.68 b | 7.1 ± 0.68 ab | 0.121 ± 0.004 a | 0.023 ± 0.002 a | |
| γ-rays (50 Gy) | 17.23 ± 0.83 a | 33 ± 1.00 a | 7 ± 0.58 a | 89.98 ± 1.92 b | 2.13 ± 0.085 a | 0.22 ± 0.71 a | 7.57 ± 0.71 a | 0.123 ± 0.076 a | 0.022 ± 0.002 a | |
| NaCl (150 mM) + Pot. silicate | 11.7 ± 0.25 cd | 21 ± 1.00 c | 5.67 ± 0.58 bcd | 54.16 ± 2.12 d | 0.85 ± 0.035 bcd | 0.13 ± 0.95 e | 6.47 ± 0.95 bcd | 0.091 ± 0.01 b | 0.019 ± 0.001 b | |
| NaCl (150 mM) + Humic acid | 12.83 ± 0.3 b | 26.67 ± 1.00 b | 5.33 ± 0.58 cd | 67.78 ± 1.17 c | 0.96 ± 0.44 bc | 0.12 ± 0.85 e | 6.53 ± 0.85 bcd | 0.077 ± 0.002 d | 0.02 ± 0.00 b | |
| NaCl (150 mM) + γ-rays | 12.73 ± 0.53 bc | 21.33 ± 1.00 c | 6.33 ± 0.58 abc | 60.99 ± 1.33 c | 1.03 ± 0.021 bc | 0.16 ± 0.40 c | 7.43 ± 0.40 a | 0.08 ± 0.002 c | 0.026 ± 0.01 a | |
| LSD at 0.05 | 1.062 | 4.315 | 1.303 | 6.62 | 0.313 | 0.002 | 0.834 | 0.002 | 0.002 | |
| Growth Parameters | Shoot Length (cm) | No. of Leaves/Plants | No. of Branches/ Plant | Leaves Area/Plant (cm)2 | Fresh Weight of Shoot (g) | Dry Weight of Shoot (g) | Root Length (cm) | Fresh Weight of Root (g) | Dry Weight of Root (g) | No. of Inflorescence/Plant | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Treatment | |||||||||||
| Control | 31.33 ± 3.79 de | 69 ± 3.60 c | 7 ± 1.00 d | 38.98 ± 1.27 e | 2.55 ± 0.29 d | 0.493 ± 0.06 de | 8.33 ± 1.53 b | 0.139 ± 0.017 d | 0.077 ± 0.015 d | 6.33 ± 1.53 d | |
| Salinity (150 mM NaCl) | 22.67 ± 2.08 f | 32.33 ± 3.79 e | 5 ± 1.00 e | 21.45 ± 1.25 f | 1.99 ± 0.37 d | 0.389 ± 0.04 e | 8.17 ± 0.76 b | 0.278 ± 0.058 c | 0.07 ± 0.016 e | 4 ± 1.00 e | |
| pot. Silicate (80 mM) | 36 b ± 2.65 c | 104 ± 5.29 a | 9 ± 1.00 bc | 93.94 ± 1.92 b | 4.77 ± 0.21 b | 0.913 ± 0.09 b | 11.73 ± 1.97 a | 0.32 ± 0.078 c | 0.11 ± 0.019 c | 10.33 ± 1.53 ab | |
| Humic acid (50 mg L−1) | 45 ± 3.60 a | 99 ± 3.60 a | 12.33 ± 0.58 a | 107.8 ± 6.37 a | 6.87 ± 1.48 a | 1.586 ± 0.33 a | 13.07 ± 1.10 a | 0.933 ± 0.153 a | 0.243 ± 0.017 a | 11 ± 1.00 a | |
| γ-rays (50 Gy) | 40 ± 1.00 b | 89 ± 3.60 b | 10 ± 1.00 b | 89.98 ± 3.03 b | 3.97 ± 0.72 c | 0.796 ± 0.06 bc | 11.07 ± 1.8 a | 0.474 ± 0.067 b | 0.19 ± 0.034 b | 8.67 ± 1.53 bc | |
| NaCl (150 mM)+Pot. Silicate | 27 ± 1.00 ef | 54.67 ± 6.43 d | 8 ± 1.00 cb | 54.16 ± 4.19 d | 2.67 ± 0.16 d | 0.431 ± 0.07 de | 11.57 ± 1.21 a | 0.284 ± 0.009 c | 0.085 ± 0.008 d | 7 ± 1.00 cd | |
| NaCl (150 mM)+Humic acid | 32.33 ± 2.51 cd | 69 ± 3.60 c | 7 ± 1.00 d | 67.18 ± 4.00 c | 2.9 ± 0.20 cd | 0.667 ± 0.08 bcd | 11.27 ± 0.68 a | 0.469 ± 0.03 b | 0.099 ± 0.019 cd | 9 ± 1.53 abc | |
| NaCl (150 mM) + γ-rays | 27.67 ± 0.76 e | 67 ± 8.19 c | 7 ± 1.00 d | 60.99 ± 5.43 c | 2.6 ± 0.10 d | 0.596 ± 0.15 cde | 9.73 ± 0.70 b | 0.358 ± 0.10 c | 0.08 ± 0.011 d | 7.67 ± 1.00 cd | |
| LSD at 0.05 | 4.56 | 9.27 | 1.644 | 6.35 | 1.134 | 0.2477 | 2.329 | 0.1238 | 0.0055 | 2.166 | |
| Yield Components | No of Fruits/Plant | No of Seeds/Plant | Weight Seeds/Plant (g) | Seed Index (Weight 1000 Seeds (g)) | |
|---|---|---|---|---|---|
| Treatment | |||||
| Control | 122.00 ± 10.00 g | 244 ± 14.00 e | 0.57 ± 0.14 f | 2.33 ± 0.14 f | |
| Salinity (150 mM NaCl) | 68.00 ± 2.00 h | 136 ± 17.00 f | 0.19 ± 0.02 g | 1.41 ± 0.02 g | |
| pot. Silicate (80 mM) | 526.65 ± 23.03 b | 1053.3 ± 142.00 b | 4.08 ± 0.35 b | 3.87 ± 0.35 b | |
| Humic acid (50 mg L−1) | 589.3 ± 10.00 a | 1178.6 ± 135.00 a | 5.78 ± 0.14 a | 4.90 ± 0.14 a | |
| γ-rays (50 Gy) | 425.00 ± 2.00 d | 850 ± 27.00 c | 2.20 ± 0.27 d | 2.59 ± 0.27 e | |
| NaCl (150 mM) + Pot. Silicate | 392.00 ± 23.00 e | 784 ± 13.00 c | 2.19 ± 0.13 d | 2.79 ± 0.13 de | |
| NaCl (150 mM) + Humic acid | 481.00 ± 23.00 c | 962 ± 127.00 b | 3.46 ± 0.33 c | 3.60 ± 0.33 c | |
| NaCl (150 mM) + γ-rays | 292.00 ± 10.00 f | 584 ± 29.00 d | 1.68 ± 0.29 e | 2.87 ± 0.29 d | |
| LSD at 0.05 | 27.62 | 104.6 | 0.2072 | 0.2072 | |
| Endogenous Phytohormones | IAA | GA3 | ABA | IAA+GA3/ABA | |
|---|---|---|---|---|---|
| Treatment | |||||
| Control | 7.12 ± 1.12 f | 389.7 ± 25.00 d | 1.90 ± 0.50 f | 208.847 | |
| Salinity (150 mM NaCl) | 2.34 ± 1.12 h | 266.2 ± 25.00 h | 11.51 ± 1.10 a | 23.329 | |
| pot. Silicate (80 mM) | 13.20 ± 1.10 b | 405.5 ± 25.00 b | 3.90 ± 1.10 e | 107.499 | |
| Humic acid (50 mg L−1) | 15.54 ± 1.12 a | 443.4 ± 25.00 a | 4.07 ± 1.10 e | 112.659 | |
| γ-rays (50 Gy) | 10.86 ± 1.12 c | 390.9 ± 25.00 c | 7.74 ± 1.10 c | 51.902 | |
| NaCl (150 mM) + Pot. Silicate | 8.66 ± 1.12 e | 299.9 ± 25.00 g | 9.26 ± 1.10 b | 33.32 | |
| NaCl (150 mM) + Humic acid | 9.37 ± 1.12 d | 367.9 ± 25.00 e | 7.98 ± 1.10 c | 47.28 | |
| NaCl (150 mM) + γ-rays | 6.25 ± 1.12 g | 313.5 ± 25.00 f | 5.50 ± 1.10 d | 58.14 | |
| LSD at 0.05 | 0.0554 | 0.00175 | 0.367 | - | |
| Pigments/Carbohydrate | Photosynthetic Pigments | Carbohydrate Fractions | ||||||
|---|---|---|---|---|---|---|---|---|
| Treatment | Chl a | Chl b | Chl a/b | Total Chl | Soluble Sugars | Polysaccharides | Total Carbohydrate | |
| Control | 13.71 ± 0.42 d | 7.08 ± 0.01 bc | 1.94 ± 0.06 | 20.79 ± 0.41 bc | 2.24 ± 0.12 d | 14.39 ± 2.11 c | 16.63 ± 1.63 f | |
| Salinity (150 mM NaCl) | 9.25 ± 0.00 f | 3.25 ± 0.22 d | 2.85 ± 0.19 | 12.5 ± 0.22 d | 3.40 ± 0.28 b | 9.46 ± 0.44 e | 12.86 ± 1.63 h | |
| pot. Silicate (80 mM) | 15.62 ± 0.12 b | 7.55 ± 0.36 b | 2.07 ± 0.08 | 23.17 ± 0.48 b | 4.04 ± 0.13 a | 19.14 ± 0.87 a | 23.18 ± 1.63 b | |
| Humic acid (50 mg L−1) | 17.1 ± 0.32 a | 9.39 ± 0.17 a | 1.82 ± 0.00 | 26.49 ± 0.49 a | 3.59 ± 0.3 b | 20.89 ± 2.13 a | 24.48 ± 1.63 a | |
| Γ-rays (50 Gy) | 14.36 ± 0.13 c | 6.65 ± 0.26 c | 2.16 ± 0.07 | 21.01 ± 0.40 bc | 2.49 ± 0.41 cd | 16.64 ± 1.38 b | 19.13 ± 1.63 e | |
| NaCl (150 mM) + Pot. Silicate | 13.97 ± 0.48 cd | 7.01 ± 0.57 bc | 1.99 ± 0.10 | 20.98 ± 1.1 bc | 4.18 ± 0.13 a | 15.67 ± 0.50 bc | 19.88 ± 1.63 c | |
| NaCl (150 mM) + Humic acid | 14.05 ± 0.26 cd | 7.71 ± 0.01 b | 1.82 ± 0.03 | 21.76 ± 0.27 b | 3.79 ± 0.2 ab | 15.43 ± 1.17 bc | 19.22 ± 1.63 d | |
| NaCl (150 mM) + γ-rays | 12.26 ± 0.15 e | 6.52 ± 0.38 bc | 1.88 ± 0.06 | 18.78 ± 0.53 c | 2.84 ± 0.1 c | 12.42 ± 0.75 d | 15.26 ± 1.63 g | |
| LSD 0.05 | 0.56 | 0.76 | - | 1.22 | 0.411 | 1.966 | 0.00175 | |
| Antioxidant Compounds | Ascorbic Acid (g/100 g D. wt.) | Carotenoids (µg/g D. wt.) | Flavonoids (g/100 g D. wt.) | Total Phenolics (g/100 g D. wt.) | Proline (µg/100 g D. wt.) | |
|---|---|---|---|---|---|---|
| Treatment | ||||||
| Control | 0.49 ± 0.01 d | 4.63 ± 0.17 b | 0.484 ± 0.08 h | 0.968 ± 0.068 c | 166.3 ± 6 c | |
| Salinity (150 mM NaCl) | 0.22 ± 0.02 h | 2.64 ± 0.21 c | 0.695 ± 0.08 a | 1.084 ± 0.066 a | 698.9 ± 5.9 a | |
| Pot. silicate (80 mM) | 0.56 ± 0.02 c | 5.84 ± 0.68 a | 0.563 ± 0.084 e | 0.898 ± 0.068 g | 239.0 ± 5.8 b | |
| Humic acid (50 mg L−1) | 0.72 ± 0.015 a | 6.07 ± 0.2 a | 0.679 ± 0.084 b | 0.978 ± 0.066 d | 184.4 ± 6 c | |
| γ-rays (50 Gy) | 0.45 ± 0.03 f | 5.7 ± 0.42 a | 0.50 ± 0.08 g | 0.882 ± 0.068 h | 218.6 ± 6.1 b | |
| NaCl (150 mM) + Pot. silicate | 0.48 ± 0.015 e | 5.61 ± 0.14 a | 0.557 ± 0.08 f | 0.966 ± 0.07 e | 234.2 ± 6 b | |
| NaCl (150 mM) + Humic acid | 0.57 ± 0.02 b | 5.74 ± 0.16 a | 0.633 ± 0.084 c | 1.007 ± 0.068 b | 266.2 ± 6 b | |
| NaCl (150 mM) + γ-rays | 0.40 ± 0.015 g | 4.89 ± 0.27 b | 0.586 ± 0.08 d | 0.955 ± 0.066 f | 283.0 ± 6.1 b | |
| LSD at 0.05 | 0.0139 | 0.27 | 0.068 | 0.052 | 4.9 | |
| Antioxidant Enzymes/MDA | Antioxidant Enzymes (Unit/mg Protein) | Lipid Peroxidation (nmol/g F.wt.) MDA | |||
|---|---|---|---|---|---|
| Treatment | PPO | POD | CAT | ||
| Control | 5622.5 ± 152.85 de | 1.49 ± 0.16 e | 0.5707 ± 0.05 d | 0.478 ± 0.015 e | |
| Salinity (150 mM NaCl) | 11688.9 ± 212.10 a | 6.21 ± 0.74 a | 1.0913 ± 0.25 ab | 1.166 ± 0.16 a | |
| pot. Silicate (80 mM) | 6754.8 ± 57.77 cd | 3.08 ± 0.21 cd | 0.831 ± 0.19 c | 0.57 d ± 0.038 e | |
| Humic acid (50 mg L−1) | 4641.8 ± 103.12 e | 3.88 ± 0.07 c | 0.451 ± 0.12 de | 0.642 ± 0.036 cd | |
| γ-rays (50 Gy) | 4092.7 ± 367.56 e | 2.95 ± 0.21 d | 0.211 ± 0.00 e | 0.553 ± 0.042 de | |
| NaCl (150 mM) + Pot. Silicate | 9215.3 ± 301.10 b | 5.23 ± 0.12 b | 1.236 ± 0.03 a | 0.64 ± 0.037 cd | |
| NaCl (150 mM) + Humic acid | 7935.4 ± 331.02 bc | 2.72 ± 0.21 d | 0.889 ± 0.17 bc | 0.699 ± 0.037 bc | |
| NaCl (150 mM) + γ-rays | 1083.8 ± 125.00 f | 2.45 ± 0.74 d | 0.423 ± 0.17 de | 0.768 ± 0.070 b | |
| LSD at 0.05 | 2085 | 0.92 | 0.2597 | 0.111 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hassanein, R.A.; Hussein, O.S.; Farag, I.A.; Hassan, Y.E.; Abdelkader, A.F.; Ibrahim, M. Salt-Stressed Coriander (Coriandrum sativum L.) Responses to Potassium Silicate, Humic Acid and Gamma Irradiation Pretreatments. Agronomy 2022, 12, 2268. https://doi.org/10.3390/agronomy12102268
Hassanein RA, Hussein OS, Farag IA, Hassan YE, Abdelkader AF, Ibrahim M. Salt-Stressed Coriander (Coriandrum sativum L.) Responses to Potassium Silicate, Humic Acid and Gamma Irradiation Pretreatments. Agronomy. 2022; 12(10):2268. https://doi.org/10.3390/agronomy12102268
Chicago/Turabian StyleHassanein, Raifa A., Omaima S. Hussein, Iman A. Farag, Yousra E. Hassan, Amal F. Abdelkader, and Mohamed Ibrahim. 2022. "Salt-Stressed Coriander (Coriandrum sativum L.) Responses to Potassium Silicate, Humic Acid and Gamma Irradiation Pretreatments" Agronomy 12, no. 10: 2268. https://doi.org/10.3390/agronomy12102268
APA StyleHassanein, R. A., Hussein, O. S., Farag, I. A., Hassan, Y. E., Abdelkader, A. F., & Ibrahim, M. (2022). Salt-Stressed Coriander (Coriandrum sativum L.) Responses to Potassium Silicate, Humic Acid and Gamma Irradiation Pretreatments. Agronomy, 12(10), 2268. https://doi.org/10.3390/agronomy12102268