

Impact of 2,4-D and Glyphosate on Soil Enzyme Activities in a Resistant Maize Cropping System


Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description, Experimental Design, and Plot Management
2.2. Sample Collection
2.3. Soil Sample Analyses
2.4. Statistics
3. Results and Discussion
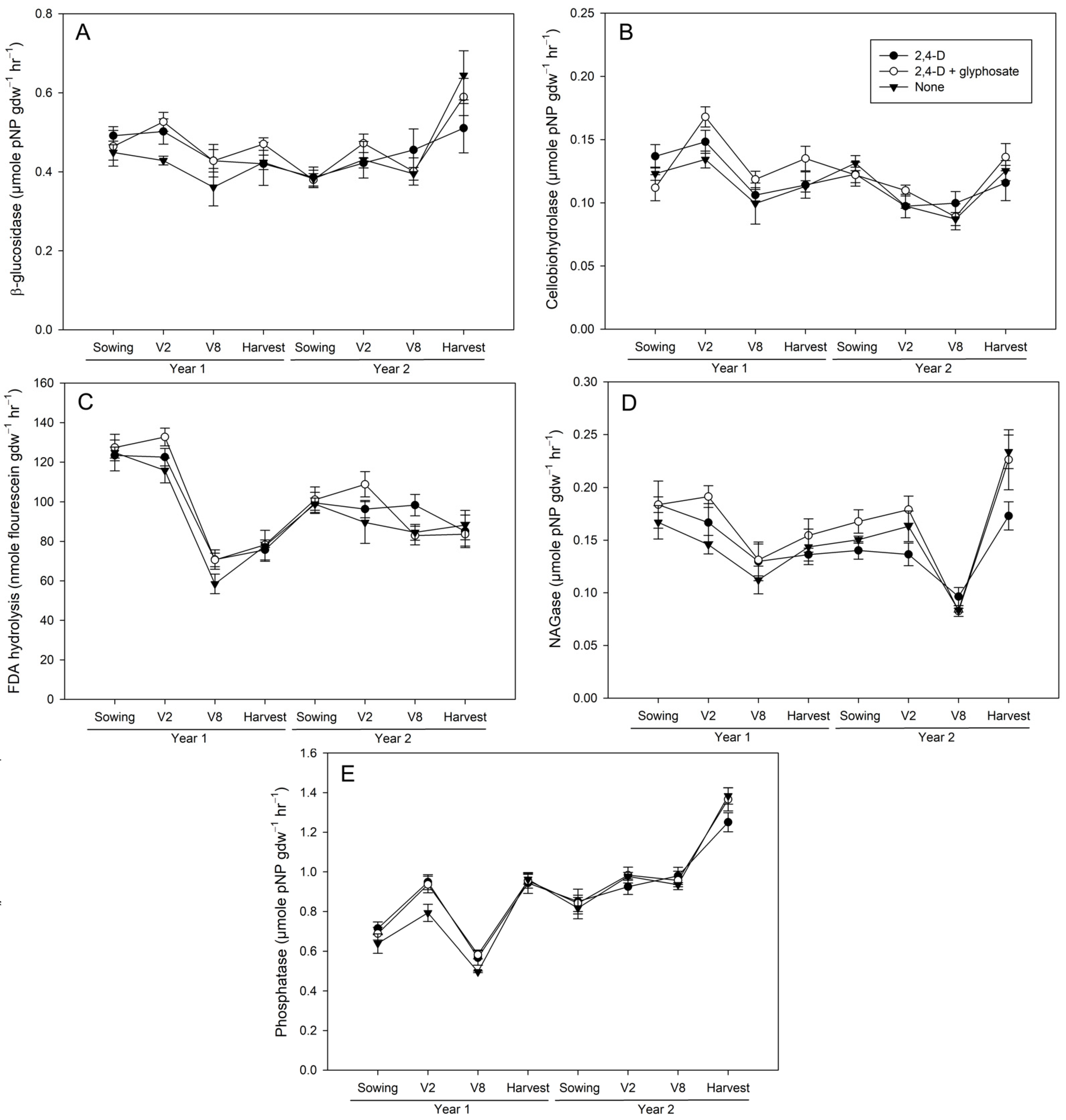
3.1. Effects of 2,4-D and Glyphosate on Bulk Soil
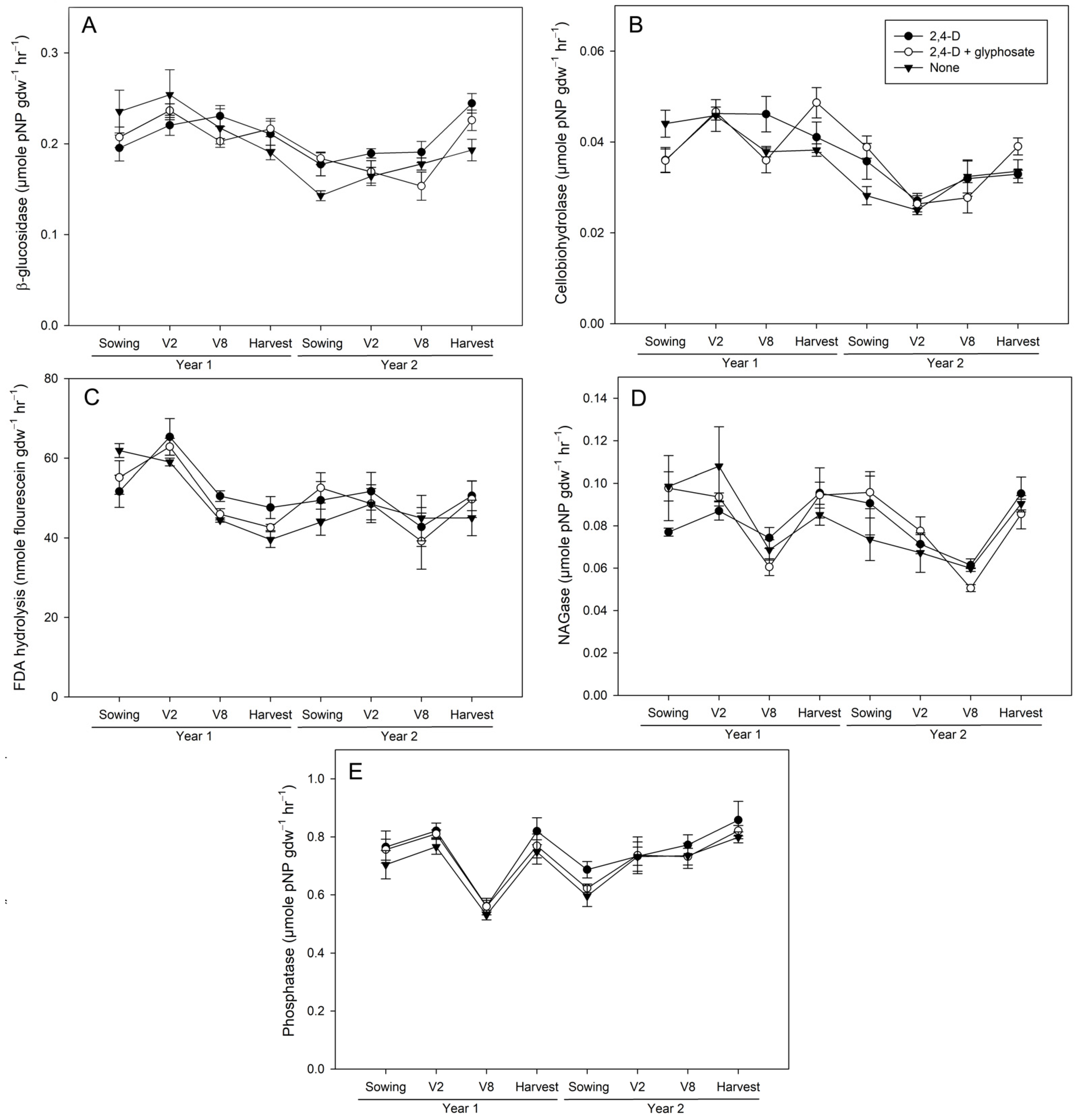
3.2. Activities in Rhizosphere Soil from 2,4-D and Glyphosate Treated Maize
4. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nandula, V.K. Herbicide Resistance Traits in Maize and Soybean: Current Status and Future Outlook. Plants 2019, 8, 337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sikorski, J.A.; Gruys, K.J. Understanding glyphosate’s molecular mode of action with EPSP Synthase: Evidence favoring an allosteric inhibitor model. Acc. Chem. Res. 1997, 30, 2–8. [Google Scholar] [CrossRef]
- Moorman, T.B.; Becerril, J.M.; Lydon, J.; Duke, S.O. Production of hydroxybenzoic acids by Bradyrhizobium japonicum strains after treatment with glyphosate. J. Agric. Food Chem. 1992, 40, 289–293. [Google Scholar] [CrossRef]
- Fischer, R.S.; Berry, A.; Gaines, C.G.; Jensen, R.A. Comparative action of glyphosate as a trigger of energy drain in Eubacteria. J. Bacteriol. 1986, 168, 1147–1154. [Google Scholar] [CrossRef] [Green Version]
- Gomez, E.; Ferreras, L.; Lovotti, L.; Fernandez, E. Impact of glyphosate application on microbial biomass and metabolic activity in a Vertic Argiudoll from Argentina. Eur. J. Soil Biol. 2009, 45, 163–167. [Google Scholar] [CrossRef]
- Dick, R.E.; Quinn, J.P. Glyphosate-degrading isolates from environmental samples: Occurrence and pathways of degradation. Appl. Microbiol. Biotechnol. 1995, 43, 545–550. [Google Scholar] [CrossRef] [PubMed]
- Haney, R.L.; Senseman, S.A.; Hons, F.M. Effect of roundup ultra on microbial activity and biomass from selected soils. J. Environ. Qual. 2002, 31, 730–735. [Google Scholar] [CrossRef] [PubMed]
- Gupta, D.; Joshi, N. Changes in microbial biomass and phosphatase activity exposed to 2,4-D and glyphosate. J. Environ. Res. Dev. 2009, 3, 663–669. [Google Scholar]
- Haney, R.L.; Senseman, S.A.; Hons, F.M.; Zuberer, D.A. Effect of glyphosate on soil microbial activity and biomass. Weed Sci. 2000, 48, 89–93. [Google Scholar] [CrossRef]
- Liphadzi, K.B.; Al-Khatib, K.; Bensch, C.N.; Stahlman, P.W.; Dille, J.A.; Todd, T.; Rice, C.W.; Horak, M.J.; Head, G. Soil microbial and nematode communities as affected by glyphosate and tillage practices in a glyphosate-resistant cropping system. Weed Sci. 2005, 53, 536–545. [Google Scholar] [CrossRef]
- Lupwayi, N.Z.; Blackshaw, R.E.; Geddes, C.M.; Dunn, R.; Petri, R.M. Multi-year and multi-site effects of recurrent glyphosate applications on the wheat rhizosphere microbiome. Environ. Res. 2022, 215, 114363. [Google Scholar] [CrossRef]
- Araújo, A.S.F.; Monteiro, R.T.R.; Abarkeli, R.B. Effect of glyphosate on the microbial activity of two Brazilian soils. Chemosphere 2003, 52, 799–804. [Google Scholar] [CrossRef] [Green Version]
- Jenkins, M.B.; Locke, M.A.; Reddy, K.N.; McChesney, D.S.; Steinriede, R.W. Impact of glyphosate-resistant corn, glyphosate applications and tillage on soil nutrient ratios, exoenzyme activities and nutrient acquisition ratios. Pest Manag. Sci. 2017, 73, 78–86. [Google Scholar] [CrossRef] [PubMed]
- Sannino, F.; Gianfreda, L. Pesticide influence on soil enzymatic activities. Chemosphere 2001, 45, 417–425. [Google Scholar] [CrossRef]
- Yu, Y.; Zhang, H.; Zhou, Q. Using soil available P and activities of soil dehydrogenase and phosphatase as indicators for biodegradation of organophosphorus pesticide methamidophos and glyphosate. Soil Sediment Contam. 2011, 20, 688–701. [Google Scholar] [CrossRef]
- Nakatani, A.S.; Fernandes, M.F.; De Souza, R.A.; Da Silva, A.P.; Dos Reis-Junior, F.B.; Mendes, I.C.; Hungria, M. Effects of the glyphosate-resistance gene and of herbicides applied to the soybean crop on soil microbial biomass and enzymes. Field Crops Res. 2014, 162, 20–29. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Zhang, M.; Tang, L.; Che, R.; Chen, H.; Blumfield, T.; Boyd, S.; Nouansyvong, M.; Xu, Z. Long-term harvest residue retention could decrease soil bacterial diversities probably due to favouring oligotrophic lineages. Microb. Ecol. 2018, 76, 771–781. [Google Scholar] [CrossRef]
- Fournier, J.C. Enumeration of the soil micro-organisms able to degrade 2,4-D by metabolism or co-metabolism. Chemosphere 1980, 9, 169–174. [Google Scholar] [CrossRef]
- Rivarola, V.; Fabra, A.; Mori, G.; Balegno, H. In vitro protein synthesis is affected by the herbicide 2,4-dichlorophenoxyacetic acid in Azospirillum brasilense. Toxicology 1992, 73, 71–79. [Google Scholar] [CrossRef]
- Fabra, A.; Duffard, R.; De Duffard, A.E. Toxicity of 2,4-dichlorophenoxyacetic acid to Rhizobium sp. in pure culture. Bull. Environ. Contam. Toxicol. 1997, 59, 645–652. [Google Scholar] [CrossRef]
- Durga Devi, K.M.; Beena, S.; Abraham, C.T. Effect of 2,4-D residues on soil microflora. J. Trop. Agric. 2008, 46, 64–66. [Google Scholar]
- Haahtela, K.; Kilpi, S.; Kari, K. Effects of phenoxy acid herbicides and glyphosate on nitrogenase activity (acetylene reduction) in root-associated Azospirillum, Enterobacter and Klebsiella. FEMS Microbiol. Lett. 1988, 53, 123–127. [Google Scholar] [CrossRef]
- Sachu, M.; Kynshi, B.L.; Syiem, M.B. A biochemical, physiological and molecular evaluation of how the herbicide 2, 4-dichlorophenoxyacetic acid intercedes photosynthesis and diazotrophy in the cyanobacterium Nostoc muscorum Meg 1. Environ. Sci. Pollut. Res. 2022, 29, 36684–36698. [Google Scholar] [CrossRef] [PubMed]
- Ding, H.; Zhang, J.; Fang, Y.; Zheng, X.; Zhang, Y.; Chen, D. Impact of herbicide 2,4-dichlorophenoxyacetic acid butyl ester on soil nitrogen-transforming bacterial populations in two soils. Int. J. Agric. Biol. 2017, 19, 812–816. [Google Scholar] [CrossRef]
- De Oliveira, E.P.; Rovida, A.F.D.S.; Martins, J.G.; Pileggi, S.A.V.; Schemczssen-Graeff, Z.; Pileggi, M. Tolerance of Pseudomonas strain to the 2,4-D herbicide through a peroxidase system. PLoS ONE 2021, 16, e0257263. [Google Scholar] [CrossRef]
- Nandula, V.K.; Tyler, H.L. Effect of new auxin herbicide formulations on control of herbicide resistant weeds and on microbial activities in the rhizosphere. Am. J. Plant Sci. 2016, 7, 2429–2439. [Google Scholar] [CrossRef] [Green Version]
- Wei, F.; Hu, X.; Xu, X. Dispersal of Bacillus subtilis and its effect on strawberry phyllosphere microbiota under open field and protection conditions. Sci. Rep. 2016, 6, 22611. [Google Scholar] [CrossRef] [Green Version]
- Mississippi State University Delta Agricultural Weather Center. Available online: http://deltaweather.extension.msstate.edu/coop-stoneville (accessed on 24 May 2022).
- NRCS Web Soil Survey. Available online: https://websoilsurvey.sc.egov.usda.gov/App/WebSoilSurvey.aspx (accessed on 29 October 2020).
- Tyler, H.L. Winter cover crops and no till management enhance enzyme activities in soybean field soils. Pedobiologia 2020, 81-82, 150666. [Google Scholar] [CrossRef]
- Jackson, C.R.; Tyler, H.L.; Millar, J.J. Determination of microbial extracellular enzyme activity in waters, soils, and sediments using high throughput microplate assays. J. Vis. Exp. 2013, 80, e50399. [Google Scholar] [CrossRef] [Green Version]
- Burns, R.G.; DeForest, J.L.; Marxsen, J.; Sinsabaugh, R.L.; Stromberger, M.E.; Wallenstein, M.D.; Weintraub, M.N.; Zoppini, A. Soil enzymes in a changing environment: Current knowledge and future directions. Soil Biol. Biochem. 2013, 58, 216–234. [Google Scholar] [CrossRef]
- Weintraub, S.R.; Wieder, W.R.; Cleveland, C.C.; Townsend, A.R. Organic matter inputs shift soil enzyme activity and allocation patterns in a wet tropical forest. Biogeochemistry 2013, 114, 313–326. [Google Scholar] [CrossRef]
- Štursová, M.; Baldrian, P. Effects of soil properties and management on the activity of soil organic matter transforming enzymes and the quantification of soil-bound and free activity. Plant Soil 2011, 338, 99–110. [Google Scholar] [CrossRef]
- Schnecker, J.; Wild, B.; Hofhansl, F.; Alves, R.J.E.; Bárta, J.; Čapek, P.; Fuchslueger, L.; Gentsch, N.; Gittel, A.; Guggenberger, G.; et al. Effects of Soil Organic Matter Properties and Microbial Community Composition on Enzyme Activities in Cryoturbated Arctic Soils. PLoS ONE 2014, 9, e94076. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Liu, X.; Dong, F.; Xu, J.; Zheng, Y.; Li, J. Soil microbial communities response to herbicide 2,4-dichlorophenoxyacetic acid butyl ester. Eur. J. Soil Biol. 2010, 46, 175–180. [Google Scholar] [CrossRef]
- Nowak, J.; Telesiński, A.; Szymczak, J. Comparison of herbicides containing isoproturon, 2.4-D and dicamba on phosphatase activity in the soil and in spring wheat (Triticum aestivum L.). Electron. J. Pol. Agric. Univ. 2006, 9, 17. [Google Scholar]
- Ljungdahl, L.G.; Eriksson, K.-E. Ecology of Microbial Cellulose Degradation. In Advances in Microbial Ecology; Marshall, K.C., Ed.; Springer: Boston, MA, USA, 1985; Volume 8, pp. 237–299. [Google Scholar]
- Tabatabai, M.A.; Ekenler, M.; Senwo, Z.N. Significance of enzyme activities in soil nitrogen mineralization. Commun. Soil Sci. Plant Anal. 2010, 41, 595–605. [Google Scholar] [CrossRef]
- Turner, B.L.; McKelvie, I.D.; Haygarth, P.M. Characterisation of water-extractable soil organic phosphorus by phosphatase hydrolysis. Soil Biol. Biochem. 2002, 34, 27–35. [Google Scholar] [CrossRef]
- Schnürer, J.; Rosswall, T. Fluorescein diacetate hydrolysis as a measure of total microbial activity in soil and litter. Appl. Environ. Microbiol. 1982, 43, 1256–1261. [Google Scholar] [CrossRef] [Green Version]
- Dennis, P.G.; Kukulies, T.; Forstner, C.; Orton, T.G.; Pattison, A.B. The effects of glyphosate, glufosinate, paraquat and paraquat-diquat on soil microbial activity and bacterial, archaeal and nematode diversity. Sci. Rep. 2018, 8, 2119. [Google Scholar] [CrossRef] [Green Version]
- Zablotowicz, R.M.; Reddy, K.N.; Weaver, M.A.; Mengistu, A.; Krutz, L.J.; Gordon, R.E.; Bellaloui, N. Cover crops, tillage, and glyphosate effects on chemical and biological properties of a lower Mississippi Delta soil and soybean yield. Environ. Res. J. 2010, 4, 227–251. [Google Scholar]
- Zabaloy, M.C.; Carné, I.; Viassolo, R.; Gómez, M.A.; Gomez, E. Soil ecotoxicity assessment of glyphosate use under field conditions: Microbial activity and community structure of Eubacteria and ammonia-oxidising bacteria. Pest Manag. Sci. 2016, 72, 684–691. [Google Scholar] [CrossRef] [PubMed]
- Zabaloy, M.C.; Garland, J.L.; Gómez, M.A. An integrated approach to evaluate the impacts of the herbicides glyphosate, 2,4-D and metsulfuron-methyl on soil microbial communities in the Pampas region, Argentina. Appl. Soil Ecol. 2008, 40, 1–12. [Google Scholar] [CrossRef]
- Lupwayi, N.Z.; Harker, K.N.; Clayton, G.W.; O’Donovan, J.T.; Blackshaw, R.E. Soil microbial response to herbicides applied to glyphosate-resistant canola. Agric. Ecosyst. Environ. 2009, 129, 171–176. [Google Scholar] [CrossRef]
- Zhang, M.; Wang, W.; Tang, L.; Heenan, M.; Xu, Z. Effects of nitrification inhibitor and herbicides on nitrification, nitrite and nitrate consumptions and nitrous oxide emission in an Australian sugarcane soil. Biol. Fert. Soils 2018, 54, 697–706. [Google Scholar] [CrossRef]
- Martens, D.A.; Bremner, J.M. Influence of herbicides on transformations of urea nitrogen in soil. J. Environ. Sci. Health Part B Pestic. Food Contam. Agric. Wastes 1993, 28, 377–395. [Google Scholar] [CrossRef]
- Newman, M.M.; Lorenz, N.; Hoilett, N.; Lee, N.R.; Dick, R.P.; Liles, M.R.; Ramsier, C.; Kloepper, J.W. Changes in rhizosphere bacterial gene expression following glyphosate treatment. Sci. Total Environ. 2016, 553, 32–41. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Activity | Year 1 | Year 2 | Field Site |
|---|---|---|---|
| Sowing | 20 March 2019 | 5 April 2021 | All plots |
| Herbicide application 1 | 17 April 2019 | 7 May 2021 | Treatment-specific plots |
| Herbicide application 2 | 17 May 2019 | 26 May 2021 | Treatment-specific plots |
| Harvest | 11 September 2019 | 15 September 2019 | All plots |
| Moisture Content (%) | pH | Organic Matter (%) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Depth | Treatment | Sowing | V2 | V8 | Harvest | Sowing | V2 | V8 | Harvest | Sowing | V2 | V8 | Harvest |
| Year 1 | |||||||||||||
| 0–5 cm | 24D | 15.22 | 15.43 | 14.76 | 6.4 | 7.11 | 7.39 | 7.02 | 6.76 | 3.95 | 3.97 | 4 | 3.72 |
| glyphosate+24D | 14.55 | 15.74 | 15.2 | 5.37 | 6.9 | 7.07 | 7 | 6.7 | 3.86 | 4.02 | 3.99 | 3.78 | |
| none | 15.1 | 14.53 | 14.56 | 6.58 | 7.02 | 7.07 | 6.98 | 6.72 | 3.9 | 3.82 | 3.84 | 3.7 | |
| 5–15 cm | 24D | 14.49 | 19.4 | 17.1 | 9.65 | 7.24 | 7.13 | 6.53 | 6.86 | 3.52 | 3.29 | 3.54 | 3.13 |
| glyphosate+24D | 15.12 | 20.89 | 15.61 | 9.19 | 7.02 | 7.14 | 6.29 | 6.81 | 3.4 | 3.17 | 3.41 | 3.11 | |
| none | 15.11 | 15.98 | 16.07 | 9.69 | 7.12 | 6.97 | 6.81 | 6.74 | 3.4 | 3.2 | 3.42 | 3.13 | |
| 2-way ANOVA | |||||||||||||
| Depth | ns | ns | 0.005 | <0.0001 | ns | ns | 0.0027 | ns | 0.001 | <0.0001 | <0.0001 | <0.0001 | |
| Treatment | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | |
| Depth × Treatment | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | |
| Year 2 | |||||||||||||
| 0–5 cm | 24D | 12.92 | 14.35 | 11.16 | 11.93 | 6.48 | 6.79 ab | 7.29 | 7.59 | 3.92 | 4.15 | 3.94 | 3.91 |
| glyphosate+24D | 12.54 | 14.96 | 11.7 | 12.86 | 6.45 | 6.83 a | 7.38 | 7.51 | 3.86 | 3.98 | 3.54 | 3.82 | |
| none | 13.46 | 14.44 | 9.85 | 13.36 | 6.5 | 6.69 b | 7.31 | 7.47 | 3.72 | 3.92 | 3.77 | 3.83 | |
| 5–15 cm | 24D | 14.31 | 16.18 | 13.29 | 14.08 | 6.47 | 7.44 cd | 7.87 | 7.59 | 3.45 | 3.75 | 3.4 | 3.45 |
| glyphosate+24D | 14.83 | 15.85 | 12.49 | 13.76 | 6.61 | 7.57 c | 7.91 | 7.59 | 3.37 | 3.65 | 3.34 | 3.281 | |
| none | 16.18 | 15.99 | 12.36 | 14.41 | 6.6 | 7.26 d | 7.92 | 7.63 | 3.28 | 3.56 | 3.37 | 3.37 | |
| 2-way ANOVA | |||||||||||||
| Depth | 0.0008 | 0.001 | 0.0324 | 0.0226 | ns | <0.0001 | <0.0001 | ns | 0.0011 | 0.0002 | 0.0215 | <0.0001 | |
| Treatment | ns | ns | ns | ns | ns | 0.0493 | ns | ns | ns | ns | ns | ns | |
| Depth × Treatment | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | |
| SM | pH | SOM | Phos | BG | NAGase | Cello | FDA | |
|---|---|---|---|---|---|---|---|---|
| SM | 1 | 0.0976 | 0.0536 | −0.322 | −0.1889 | −0.0849 | −0.1996 | 0.0546 |
| pH | 1 | 0.0293 | 0.2219 | −0.0343 | −0.1118 | −0.1551 | −0.1179 | |
| SOM | 1 | 0.2048 | 0.6585 | 0.508 | 0.6598 | 0.6381 | ||
| Phos | 1 | 0.5895 | 0.5492 | 0.4467 | 0.3753 | |||
| BG | 1 | 0.8343 | 0.9197 | 0.7793 | ||||
| NAGase | 1 | 0.8223 | 0.7262 | |||||
| Cello | 1 | 0.8469 | ||||||
| FDA | 1 |
| Year 1 | Year 2 | |||
|---|---|---|---|---|
| Treatment | V2 | V8 | V2 | V8 |
| Beta-glucosidase | ||||
| 2,4-D | 1.74 ± 0.23 | 1.32 ± 0.17 | 1.33 ± 0.09 | 2.62 ± 0.42 |
| 2,4-D+Glyphosate | 2.42 ± 0.39 | 1.66 ± 0.36 | 1.21 ± 0.09 | 2.24 ± 0.1 |
| No herbicide | 1.98 ± 0.47 | 2.06 ± 0.3 | 1.76 ± 0.31 | 2.36 ± 0.33 |
| Cellobiohydrolase | ||||
| 2,4-D | 0.53 ± 0.09 | 0.22 ± 0.01 | 0.23 ± 0 | 0.26 ± 0.01 |
| 2,4-D+Glyphosate | 0.61 ± 0.07 | 0.27 ± 0.05 | 0.2 ± 0.01 | 0.26 ± 0.01 |
| No herbicide | 0.52 ± 0.08 | 0.31 ± 0.02 | 0.27 ± 0.03 | 0.27 ± 0.02 |
| NAGase | ||||
| 2,4-D | 0.72 ± 0.05 | 0.38 ± 0.05 | 0.34 ± 0.02 | 0.53 ± 0.05 |
| 2,4-D+Glyphosate | 0.56 ± 0.06 | 0.38 ± 0.07 | 0.33 ± 0.02 | 0.46 ± 0.05 |
| No herbicide | 0.53 ± 0.09 | 0.43 ± 0.04 | 0.35 ± 0.07 | 0.49 ± 0.05 |
| Phosphatase | ||||
| 2,4-D | 3.53 ± 0.17 | 2.03 ± 0.19 | 4.11 ± 0.22 | 4.98 ± 0.69 |
| 2,4-D+Glyphosate | 3.98 ± 0.59 | 2.24 ± 0.42 | 3.74 ± 0.25 | 4.13 ± 0.31 |
| No herbicide | 3.49 ± 0.59 | 2.66 ± 0.24 | 4.6 ± 0.54 | 4.17 ± 0.52 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tyler, H.L. Impact of 2,4-D and Glyphosate on Soil Enzyme Activities in a Resistant Maize Cropping System. Agronomy 2022, 12, 2747. https://doi.org/10.3390/agronomy12112747
Tyler HL. Impact of 2,4-D and Glyphosate on Soil Enzyme Activities in a Resistant Maize Cropping System. Agronomy. 2022; 12(11):2747. https://doi.org/10.3390/agronomy12112747
Chicago/Turabian StyleTyler, Heather L. 2022. "Impact of 2,4-D and Glyphosate on Soil Enzyme Activities in a Resistant Maize Cropping System" Agronomy 12, no. 11: 2747. https://doi.org/10.3390/agronomy12112747
APA StyleTyler, H. L. (2022). Impact of 2,4-D and Glyphosate on Soil Enzyme Activities in a Resistant Maize Cropping System. Agronomy, 12(11), 2747. https://doi.org/10.3390/agronomy12112747