
Photosynthesis Alterations in Wheat Plants Induced by Herbicide, Soil Drought or Flooding





Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growing Conditions and Treatments
2.2. Leaf Pigment Content
2.3. Leaf Gas Exchange Parameters
2.4. Chlorophyll a Fluorescence Parameters
2.5. Assessment of Recovery and Resilience Indices of Photosynthesis Traits
2.6. Statistical Analysis
3. Results
3.1. Phenotypic Alterations
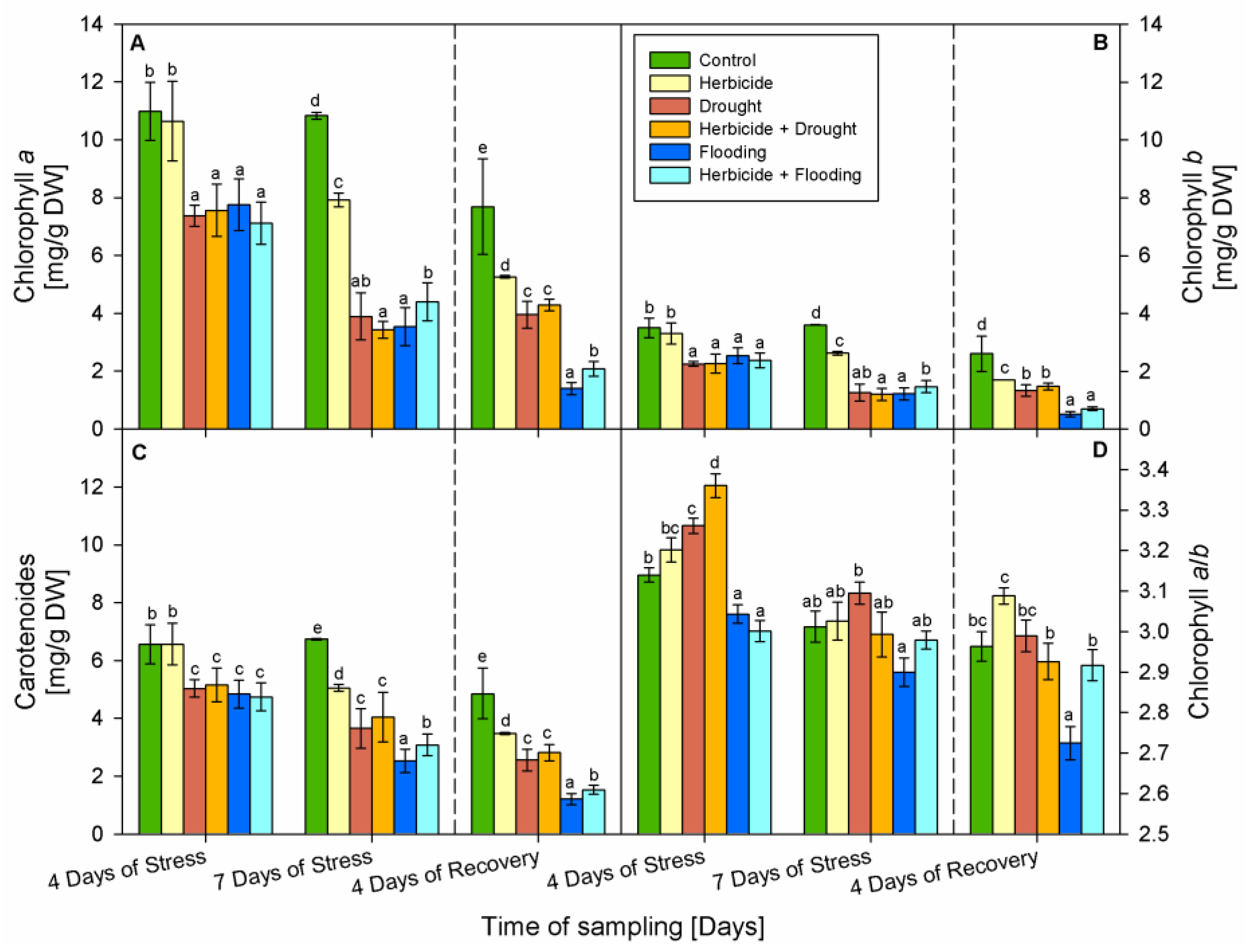
3.2. Leaf Pigments Content
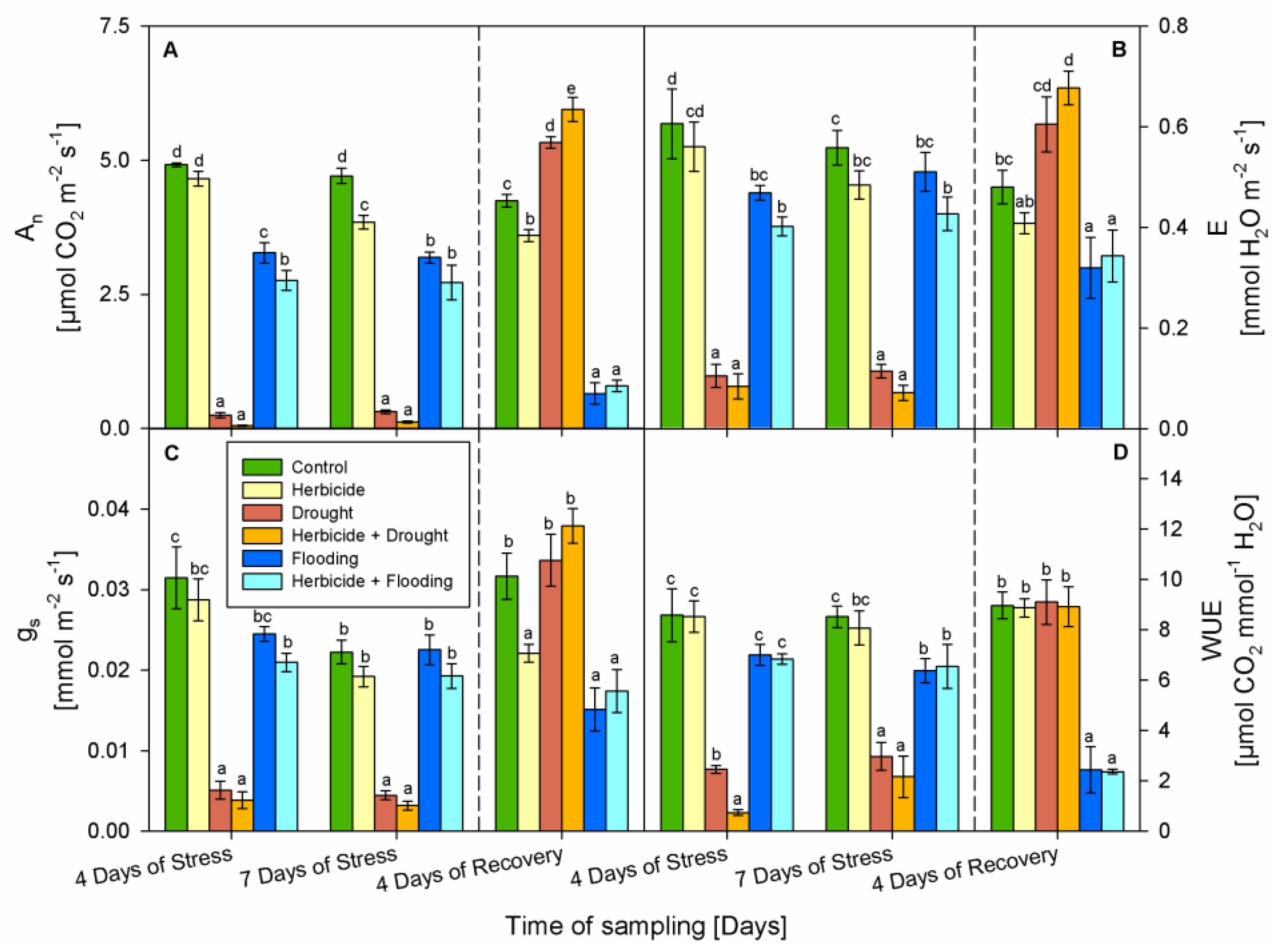
3.3. Gas Exchange Parameters
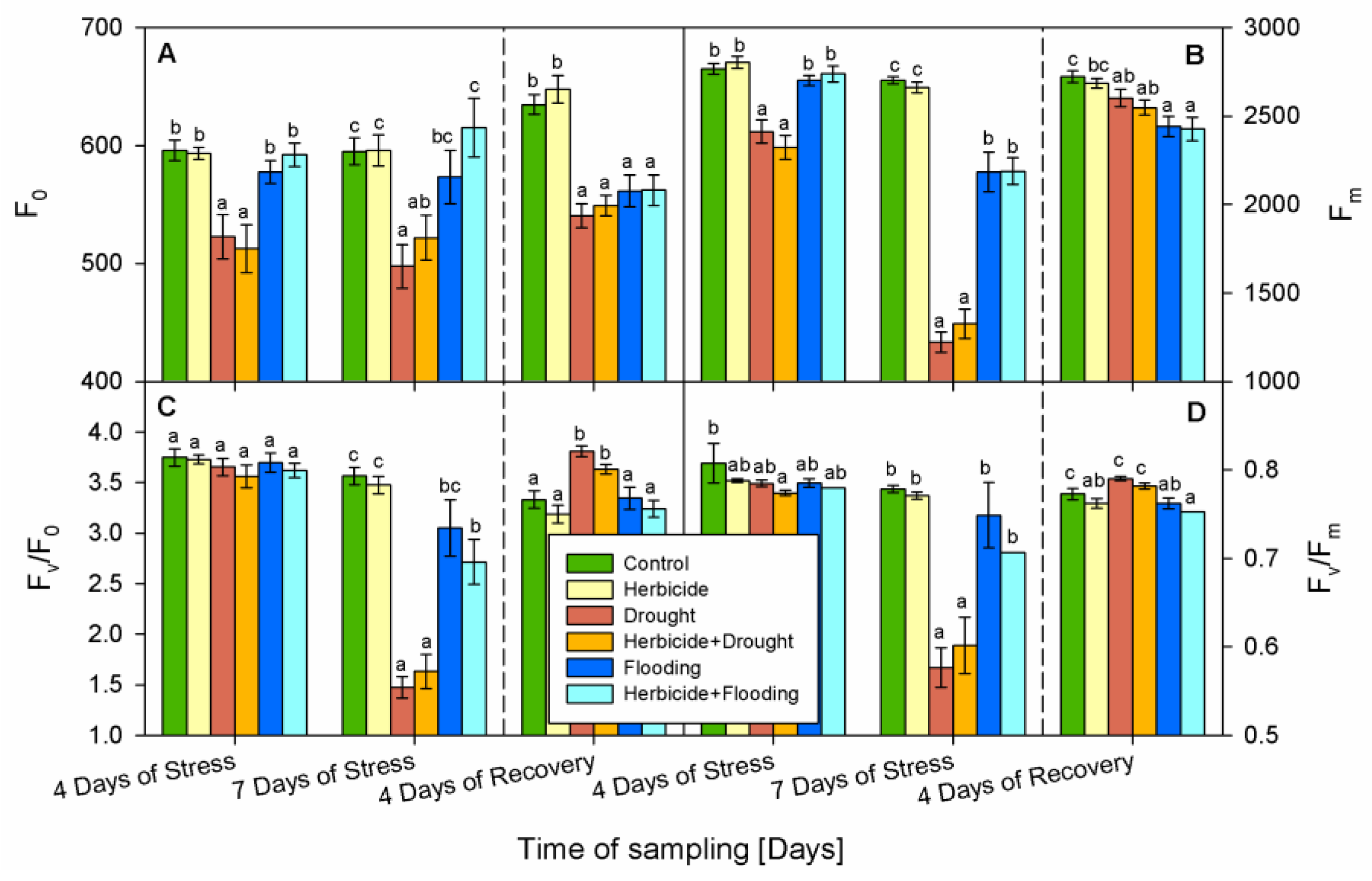
3.4. Chlorophyll Fluorescence Parameters
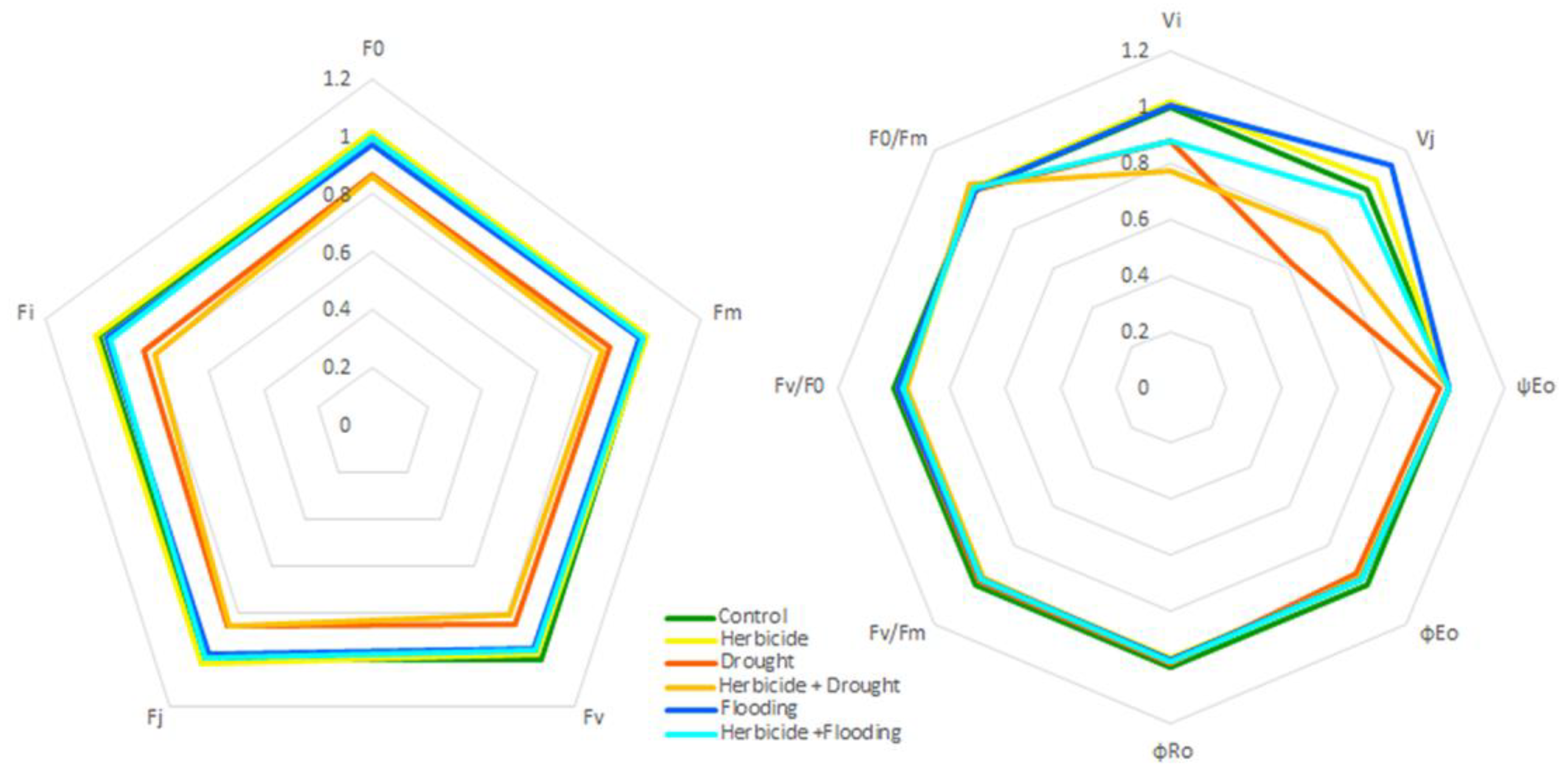
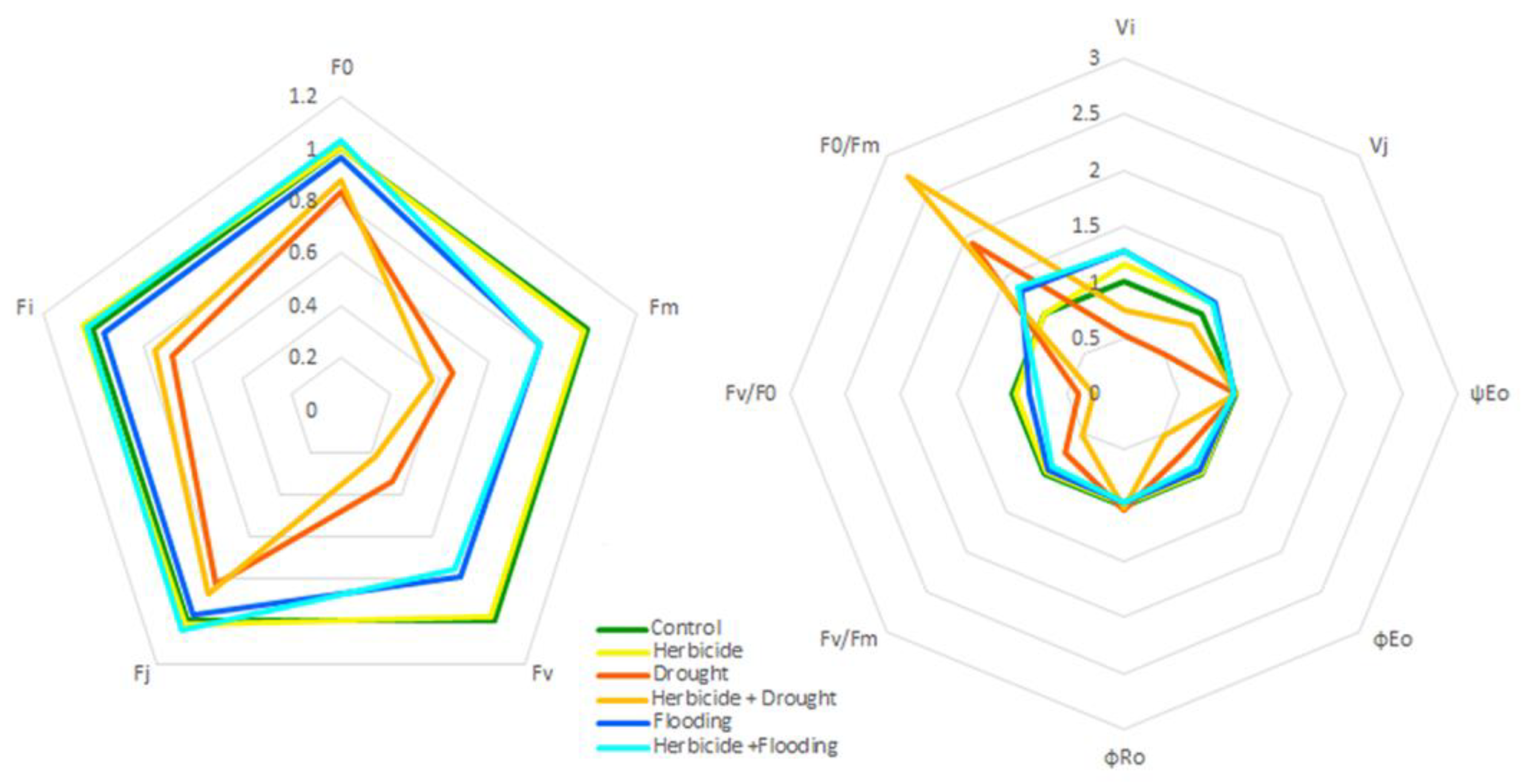
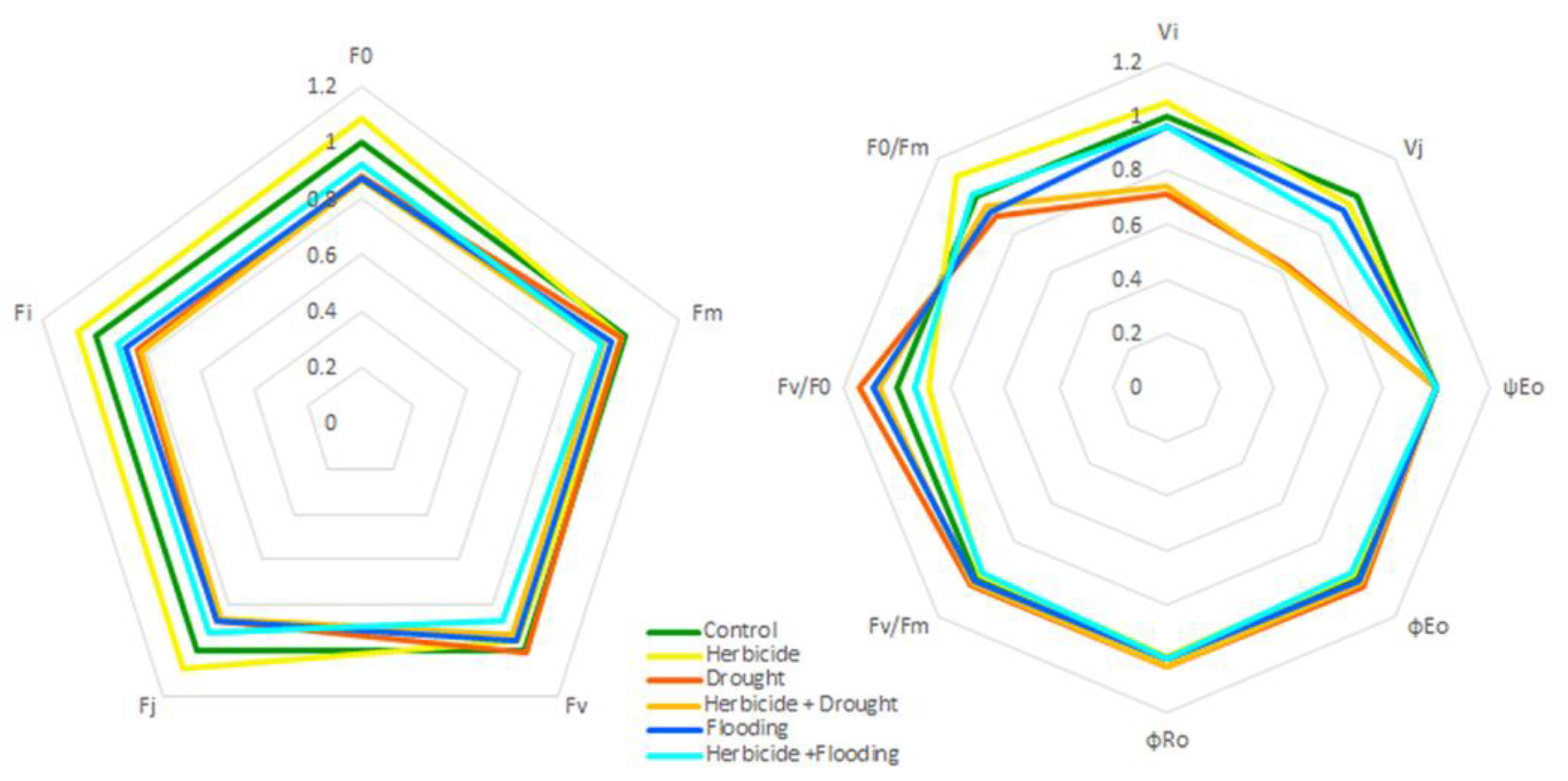
3.5. Spider Plot Analyses of Fluorescence Parameters
3.6. Assessment of the Recovery and Resilience Indices of Photosynthesis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nakka, S.; Jugulam, M.; Peterson, D.; Asif, M. Herbicide resistance: Development of wheat production systems and current status of resistant weeds in wheat cropping systems. Crop J. 2019, 7, 750–760. [Google Scholar] [CrossRef]
- Available online: https://sitem.herts.ac.uk/aeru/ppdb/en/atoz.htm (accessed on 12 January 2022).
- Mutava, R.N.; Prince, S.J.K.; Syed, N.H.; Song, L.; Valliyodan, B.; Chen, W.; Nguyen, H.T. Understanding abiotic stress tolerance mechanisms in soybean: A comparative evaluation of soybean response to drought and flooding stress. Plant Physiol. Biochem. 2015, 86, 109–120. [Google Scholar] [CrossRef]
- Qi, M.; Liu, X.; Li, Y.; Song, H.; Yin, Z.; Zhang, F.; He, Q.; Xu, Z.; Zhou, G. Photosynthetic resistance and resilience under drought, fooding and rewatering in maize plants. Photosynth. Res. 2021, 148, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Zhou, G.; Shimizu, H. Plant responses to drought and rewatering. Plant Signal. Behav. 2010, 5, 649–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basu, S.; Ramegowda, V.; Kumar, A.; Pereira, A. Plant adaptation to drought stress. F1000Research 2016, 5, 1554. [Google Scholar] [CrossRef] [PubMed]
- Georgieva, K.; Maslenkova, L.; Peeva, V.; Markovska, Y.; Stefanov, D.; Tuba, Z. Comparative study on the changes in photosynthetic activityof the homoiochlorophyllous desiccation-tolerant Haberlea rhodopensis and desiccation-sensitive spinach leaves during desiccation and rehydration. Photosynth. Res. 2005, 85, 191–203. [Google Scholar] [CrossRef] [PubMed]
- Abid, M.; Ali, S.; Qi, L.K.; Zahoor, R.; Tian, Z.; Jiang, D.; Snider, J.L.; Dai, T. Physiological and biochemical changes during drought and recovery periods at tillering and jointing stages in wheat (Triticum aestivum L.). Sci. Rep. 2018, 8, 4615. [Google Scholar] [CrossRef] [PubMed]
- Pan, J.; Sharif, R.; Xu, X.; Chen, X. Mechanisms of waterlogging tolerance in plants: Research progress and prospects. Front. Plant Sci. 2021, 11, 627331. [Google Scholar] [CrossRef]
- Luan, H.; Shen, H.; Pan, Y.; Guo, B.; Lv, C.; Xu, R. Elucidating the hypoxic stress response in barley (Hordeum vulgare L.) during waterlogging: A proteomics approach. Sci. Rep. 2018, 8, 9655. [Google Scholar] [CrossRef]
- Malik, A.I.; Colmer, D.T.; Lambers, H.; Schortemeyer, M. Changes in physiological and morphological traits of roots and shoots of wheat in response to different depths of waterlogging. Aust. J. Plant Physiol. 2001, 28, 1121–1131. [Google Scholar] [CrossRef]
- Todorova, D.; Sergiev, I.; Katerova, Z.; Shopova, E.; Dimitrova, L.; Brankova, L. Assessment of the biochemical responses of wheat seedlings to soil drought after application of selective herbicide. Plants 2021, 10, 733. [Google Scholar] [CrossRef] [PubMed]
- Katerova, Z.; Sergiev, I.; Todorova, D.; Shopova, E.; Dimitrova, L.; Brankova, L. Physiological responses of wheat seedlings to soil waterlogging applied after treatment with selective herbicide. Plants 2021, 10, 1195. [Google Scholar] [CrossRef] [PubMed]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- LiCor. Using the LI-6400/LI-6400XT Portable Photosynthesis System; LI-COR Biosciences: Lincoln, NE, USA, 2012; p. 846. [Google Scholar]
- Strasser, R.J.; Tsimilli-Michael, M.; Srivastava, A. Analysis of the chlorophyll a fluorescence transient. In Chlorophyll a Fluorescence: A Signature of Photosynthesis; Papageorgiou, G.C., Govindjee, Eds.; Springer: Dordrecht, The Netherlands, 2004; pp. 321–362. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Oukarroum, A.; Alexandrov, V.; Kouzmanova, M.; Brestic, M.; Zivcak, M.; Samborska, I.A.; Cetner, M.D.; Allakhverdiev, S.I.; Goltsev, V. Identification of nutrient deficiency in maize and tomato plants by in vivo chlorophyll a fluorescence measurements. Plant Physiol. Biochem. 2014, 81, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Katam, R.; Shokri, S.; Murthy, N.; Singh, S.K.; Suravajhala, P.; Khan, M.N.; Bahmani, M.; Sakata, K.; Reddy, K.R. Proteomics, physiological, and biochemical analysis of cross tolerance mechanisms in response to heat and water stresses in soybean. PLoS ONE 2020, 15, e0233905. [Google Scholar] [CrossRef]
- Wu, X.; Tang, Y.; Li, C.; Wu, C.; Huang, G. Chlorophyll fluorescence and yield responses of winter wheat to waterlogging at different growth stages. Plant Prod. Sci. 2015, 18, 284–294. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Li, H.; Gerhards, R. Chlorophyll fluorescence response to herbicide stress in Alopecurus myosuroides. Julius-Kühn-Archiv. In Proceedings of the 27th German Conference on Weed Biology and Weed Control, Braunschweig, Germany, 23–25 February 2016; Volume 452, pp. 57–67. [Google Scholar] [CrossRef]
- Hassannejad, S.; Porheidar Ghafarbi, S. Assessment of some chlorophyll a fluorescence parameters of different corn cultivars in response to clodinafop-propagrgyl herbicide and salicylic acid. J. Plant. Physiol. Breed. 2018, 8, 47–57. [Google Scholar]
- Simova-Stoilova, L.; Pecheva, D.; Kirova, E. Drought stress response in winter wheat varieties –changes in leaf proteins and proteolytic activities. Acta Bot. Croat. 2020, 79, 121–130. [Google Scholar] [CrossRef]
- Marcińska, I.; Czyczyło-Mysza, I.; Skrzypek, E.; Filek, M.; Grzesiak, S.; Grzesiak, M.T.; Janowiak, F.; Hura, T.; Dziurka, M.; Dziurka, K.; et al. Impact of osmotic stress on physiological and biochemical characteristics in drought-susceptible and drought-resistant wheat genotypes. Acta Physiol. Plant. 2012, 35, 451–461. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Liu, X.; Zhang, Z.; Liu, N.; Li, D.; Hu, L. Melatonin improved waterlogging tolerance in alfalfa (Medicago sativa) by reprogramming polyamine and ethylene metabolism. Front. Plant Sci. 2019, 10, 44. [Google Scholar] [CrossRef]
- Yordanova, R.Y.; Alexieva, V.S.; Popova, L.P. Influence of root oxygen deficiency on photosynthesis and antioxidant status in barley plants. Russ. J. Plant Physiol. 2003, 50, 163–167. [Google Scholar] [CrossRef]
- Yordanova, R.Y.; Uzunova, A.N.; Popova, L.P. Effects of short-term soil flooding on stomata behaviour and leaf gas exchange in barley plants. Biol. Plant. 2005, 49, 317–319. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Carpentier, R.; Allakhverdiev, S.I.; Bosa, K. Fluorescence parameters as early indicators of light stress in barley. J. Photochem. Photobiol. B Biol. 2012, 112, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Zivcak, M.; Brestic, M.; Balatova, Z.; Drevenakova, P.; Olsovska, K.; Kalaji, H.M.; Yang, X.; Allakhverdiev, S.I. Photosynthetic electron transport and specific photoprotective responses in wheat leaves under drought stress. Photosynth. Res. 2013, 117, 529–546. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Jajoo, A.; Oukarroum, A.; Brestic, M.; Zivcak, M.; Samborska, I.A.; Cetner, M.D.Ł.; Goltsev, I.V.; Ladle, R.J. Chlorophyll a fluorescence as a tool to monitor physiological status of plants under abiotic stress conditions. Acta Physiol. Plant. 2016, 38, 102. [Google Scholar] [CrossRef] [Green Version]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence—A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef]
- Borawska-Jarmułowicz, B.; Mastalerczuk, G.; Pietkiewicz, S.; Kalaji, M.H. Low temperature and hardening effects on photosynthetic apparatus efficiency and survival of forage grass varieties. Plant Soil Environ. 2014, 60, 177–183. [Google Scholar] [CrossRef] [Green Version]
- Ikeda, Y.; Shinpei, O.; Kazuya, K.; Akira, T.; Hiroyuki, W.; Hitoshi, K.; van Rensem, J.J.S.; Boger, P.; Waksbayashi, K. Binding site of novel 2-benzylamino-4-methyl-6-trifluromethyl-1, 3, 5- triazine herbicides in the D1 protein of photosystem II. Photosynth. Res. 2003, 77, 35–43. [Google Scholar] [CrossRef]
- Van Ruijven, J.; Berendse, F. Diversity enhances community recovery, but not resistance, after drought. J. Ecol. 2010, 98, 81–86. [Google Scholar] [CrossRef]
- Mariotte, P.; Vandenberghe, C.; Kardol, P.; Hagedorn, F.; Buttler, A. Subordinate plant species enhance community resistance against drought in semi-natural grasslands. J. Ecol. 2013, 101, 763–773. [Google Scholar] [CrossRef]
- Pinheriro, C.; Passarinho, J.A.; Ricardo, C.P. Effect of drought and rewatering on the metabolism of Lupinus albus organs. J. Plant Physiol. 2004, 161, 1203–1210. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Traits | Treatments | Recovery | Resilience |
|---|---|---|---|
| Herbicide | −6 ▬ | −15 ▼ | |
| Drought | 1612 ▲ ▲ ▲ ▲ | 26 ▲ | |
| An | Herbicide + Drought | 4851 ▲ ▲ ▲ ▲ | 40 ▲ |
| Flooding | −79 ▼ ▼ | −85 ▼ ▼ | |
| Herbicide + Flooding | −71 ▼ ▼ | −81 ▼ ▼ | |
| Herbicide | −16 ▼ | −15 ▼ | |
| Drought | 428 ▲ ▲ ▲ ▲ | 26 ▲ | |
| E | Herbicide + Drought | 854 ▲ ▲ ▲ ▲ | 41 ▲ ▲ |
| Flooding | −37 ▼ | −33 ▼ | |
| Herbicide + Flooding | −20 ▼ | −29 ▼ | |
| Herbicide | 10 ▬ | −1 ▬ | |
| Drought | 207 ▲ ▲ ▲ ▲ | 2 ▬ | |
| WUE | Herbicide + Drought | 312 ▲ ▲ ▲ ▲ | −1 ▬ |
| Flooding | −62 ▼ ▼ | −73 ▼ ▼ ▼ | |
| Herbicide + Flooding | −64 ▼ ▼ | −74 ▼ ▼ ▼ | |
| Herbicide | 15 ▲ | −30 ▼ | |
| Drought | 654 ▲ ▲ ▲ ▲ | 6 ▬ | |
| Gs | Herbicide + Drought | 1095 ▲ ▲ ▲ ▲ | 20 ▲ |
| Flooding | −33 ▼ | −52 ▼ ▼ | |
| Herbicide + Flooding | −10 ▬ | −45 ▼ ▼ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Todorova, D.; Aleksandrov, V.; Anev, S.; Sergiev, I. Photosynthesis Alterations in Wheat Plants Induced by Herbicide, Soil Drought or Flooding. Agronomy 2022, 12, 390. https://doi.org/10.3390/agronomy12020390
Todorova D, Aleksandrov V, Anev S, Sergiev I. Photosynthesis Alterations in Wheat Plants Induced by Herbicide, Soil Drought or Flooding. Agronomy. 2022; 12(2):390. https://doi.org/10.3390/agronomy12020390
Chicago/Turabian StyleTodorova, Dessislava, Vladimir Aleksandrov, Svetoslav Anev, and Iskren Sergiev. 2022. "Photosynthesis Alterations in Wheat Plants Induced by Herbicide, Soil Drought or Flooding" Agronomy 12, no. 2: 390. https://doi.org/10.3390/agronomy12020390
APA StyleTodorova, D., Aleksandrov, V., Anev, S., & Sergiev, I. (2022). Photosynthesis Alterations in Wheat Plants Induced by Herbicide, Soil Drought or Flooding. Agronomy, 12(2), 390. https://doi.org/10.3390/agronomy12020390