
Comparison of Trapping Effects of Different Traps and Monitoring the Occurrence Dynamics of Spodoptera litura in Soybean Fields of Dangtu, Anhui Province, China


 ,
, 

Abstract
:1. Introduction
2. Materials and Methods
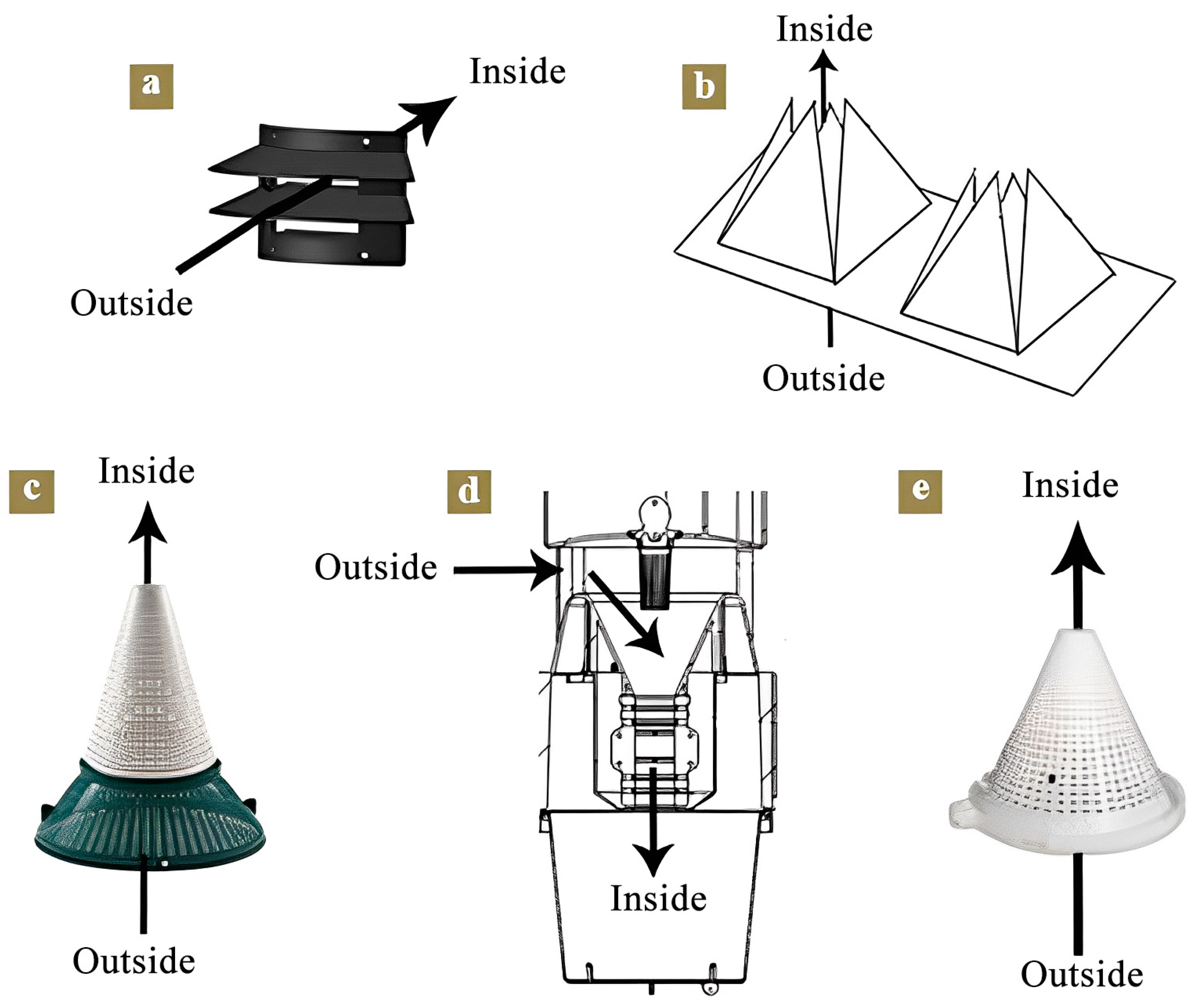
2.1. Trap Types and Pheromone Lure
2.2. Experimental Design and Investigation Method
2.2.1. Trapping Efficiency Comparison of S. litura with Different Traps
2.2.2. Effect of the Reuse of Trap YL-VT on Subsequent Traps of S. litura
2.2.3. Trapping of S. litura by AIM
2.2.4. Trapping of S. litura by AIM-lite-A
2.3. Data Analysis
3. Results
3.1. Comparison of Catches of Different Traps
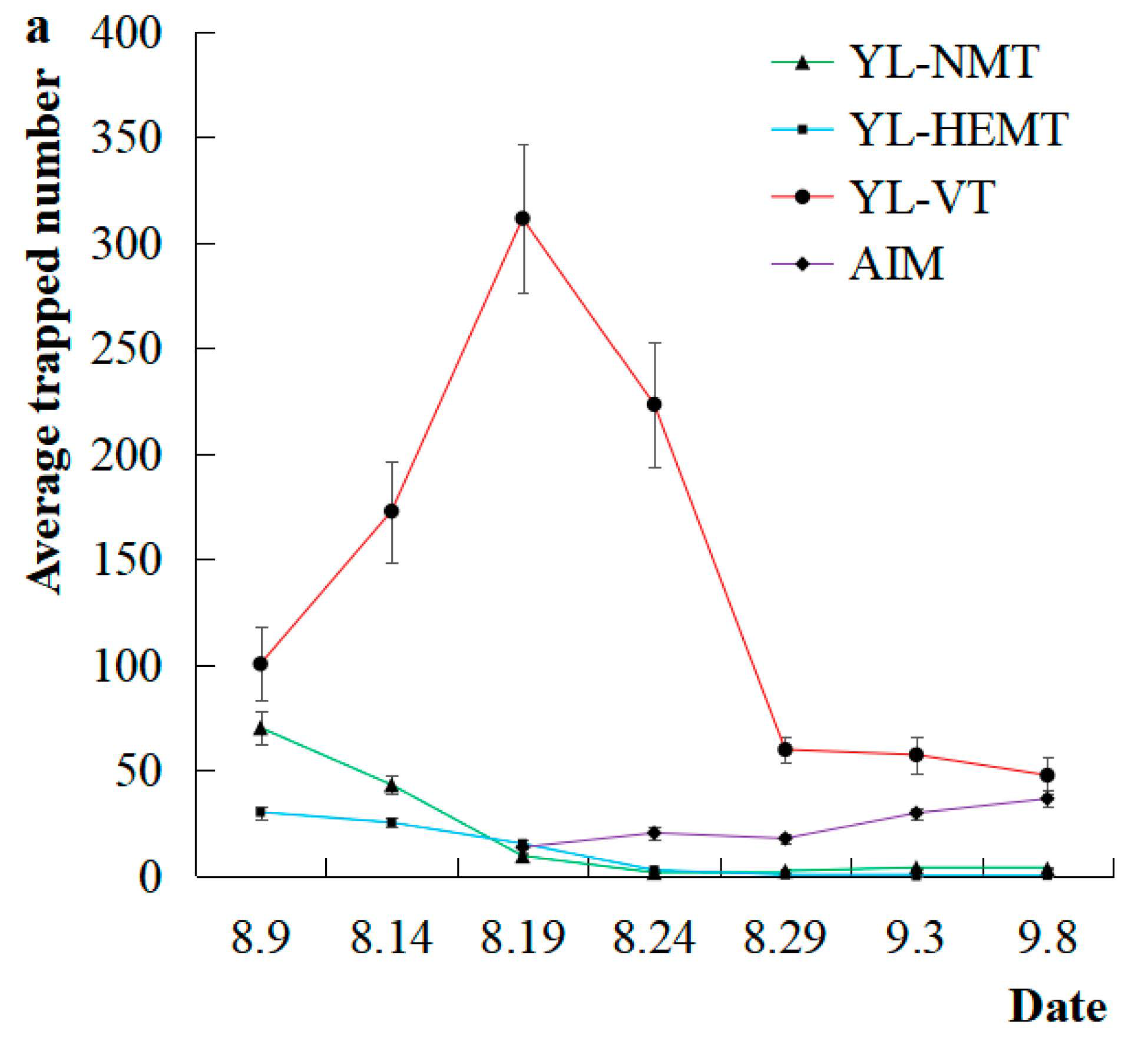
3.1.1. Trapping of S. litura with Different Traps
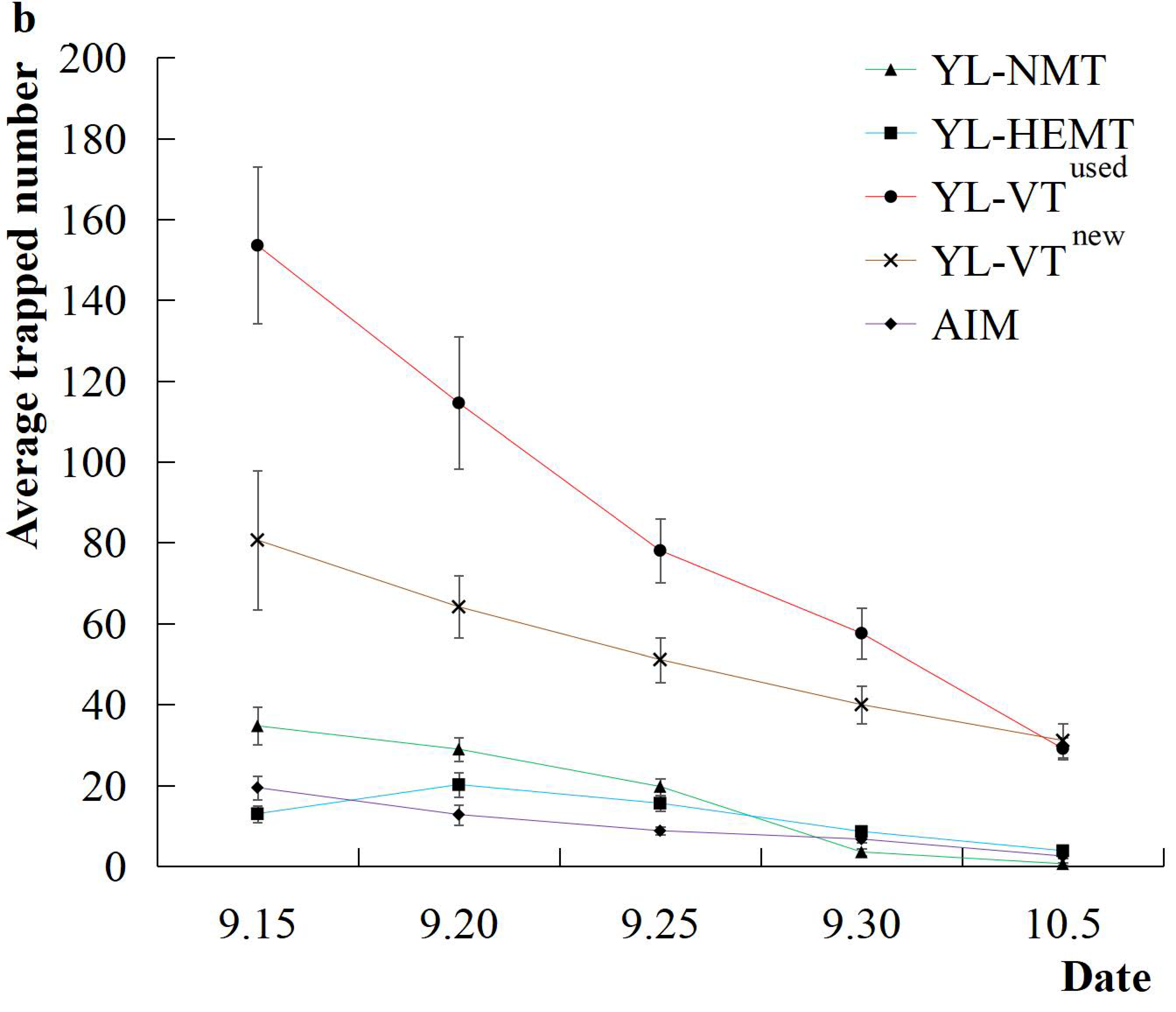
3.1.2. Effect on Trapping from Subsequently Reused YL-VT Traps
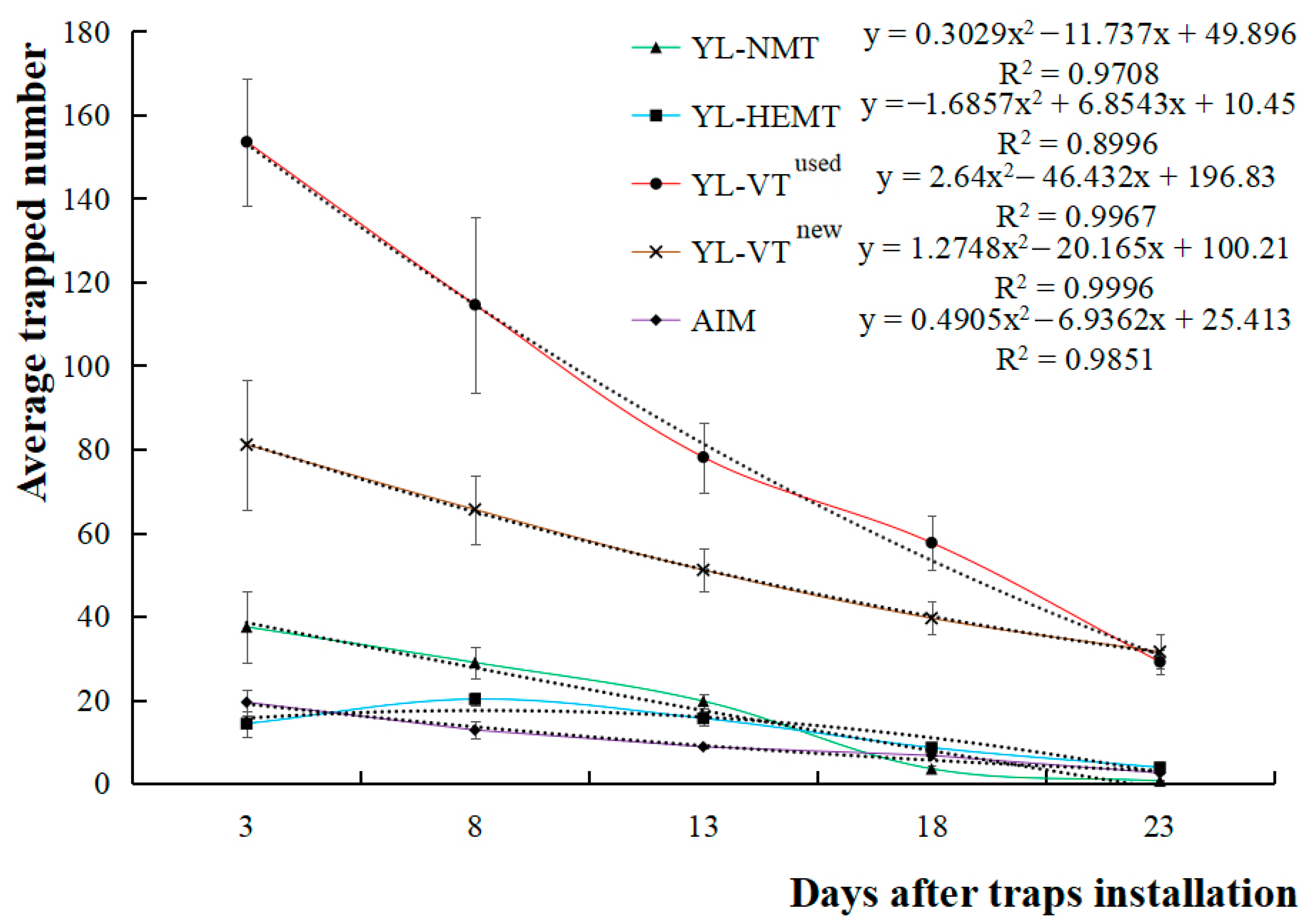
3.1.3. Half-Life of Pheromone Lure Core in Different Traps
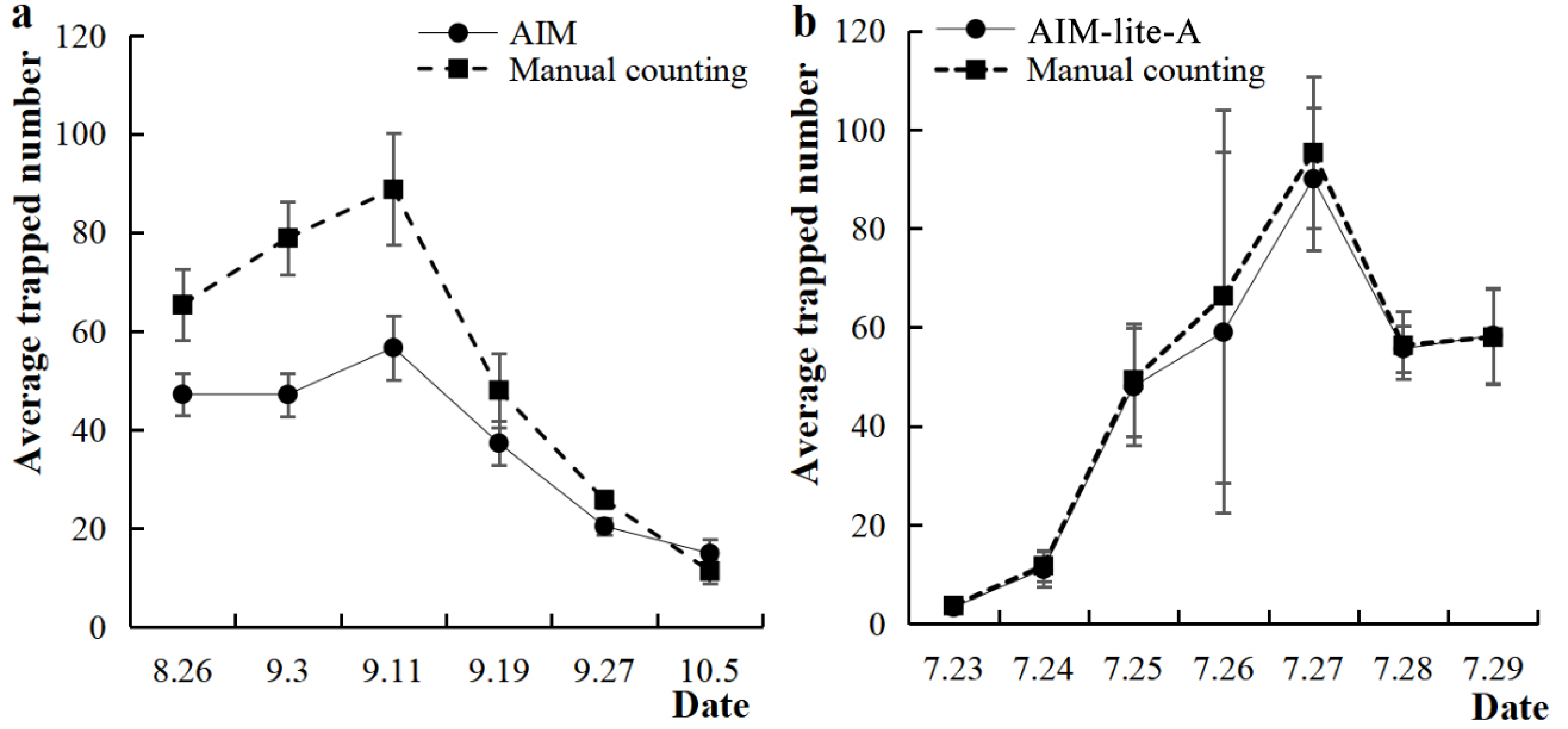
3.2. Accuracy of AIM and AIM-Lite-A in Monitoring S. litura
3.3. Application of AIM and AIM-Lite-A in Monitoring S. litura
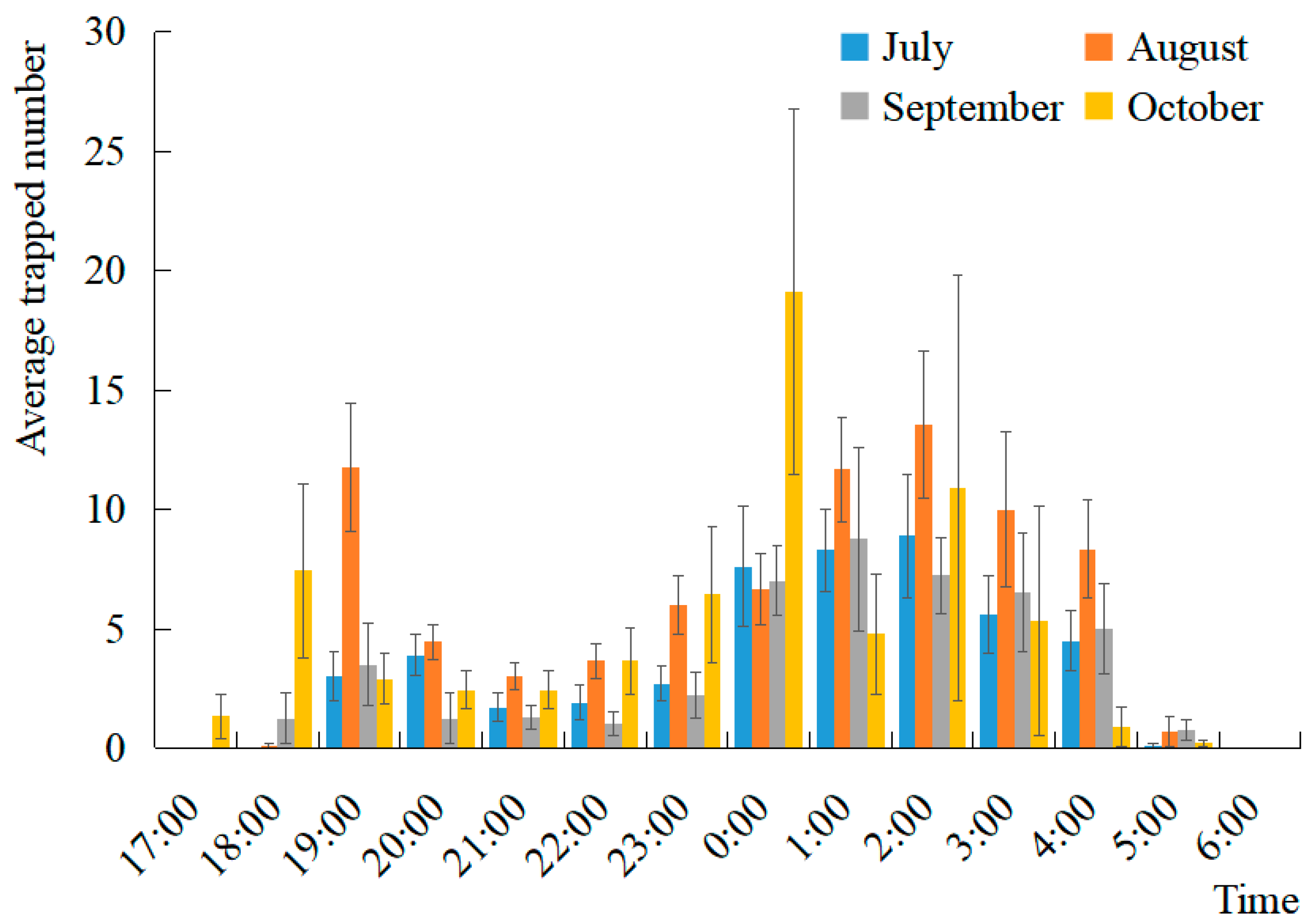
3.3.1. Diurnal Rhythm of Moth Activity among Main Months of S. litura Occurrence
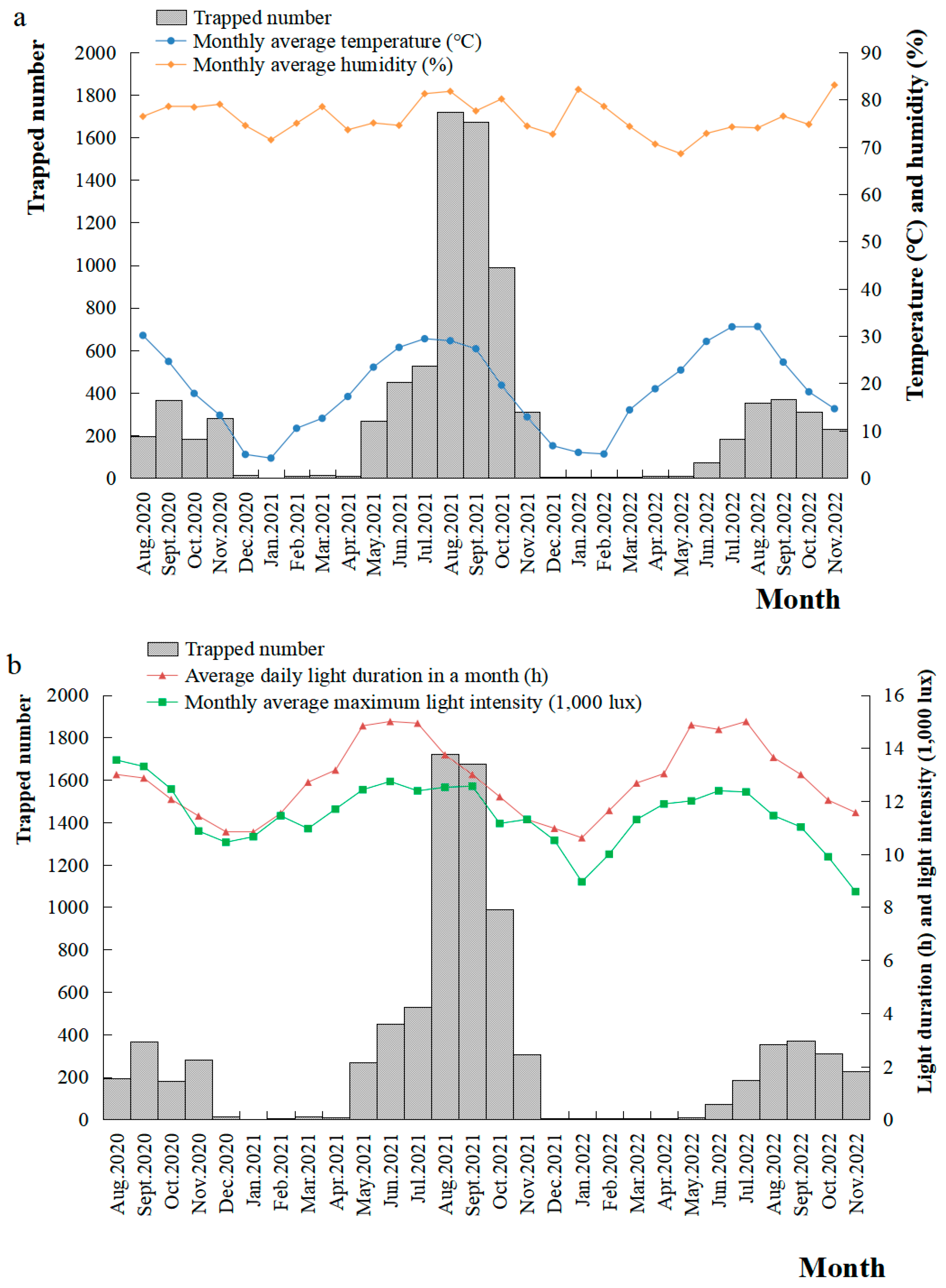
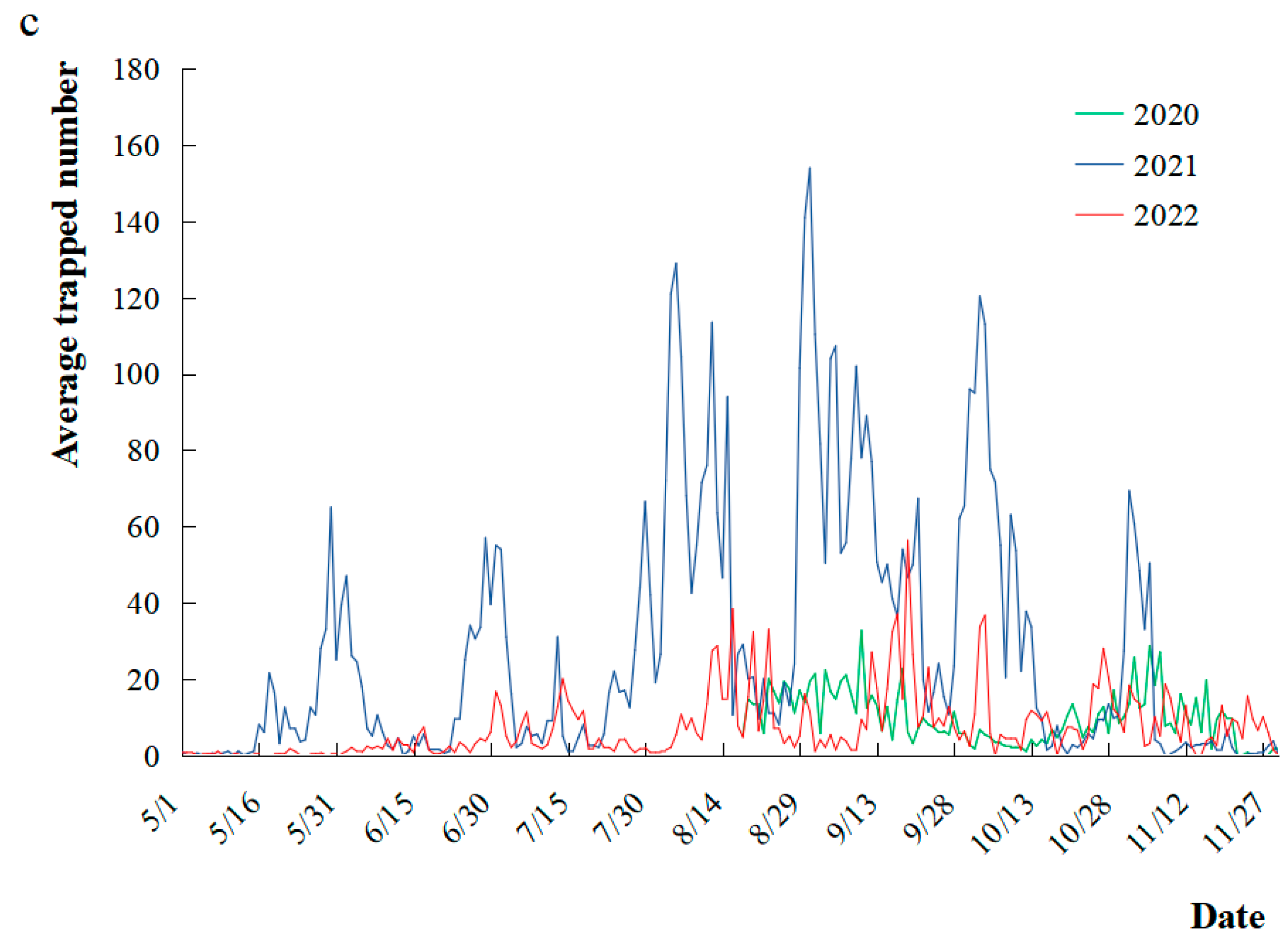
3.3.2. Seasonal Rhythm of S. litura Moth Occurrence and Its Relationship with Meteorological Factors
4. Discussion
4.1. Factors Affecting Trapping Efficiency of S. litura Traps
4.1.1. Configuration Parameters of Inlet Devices and Collecting Devices
4.1.2. Influence of Pheromone Lure and Its Installed Position on Trapping Effect
4.2. Future Application and Improvement of AIM-Lite-A and AIM
4.3. Application of Pheromone Traps in Pest Control
4.4. Insect Resistant Soybean Breeding to S. litura
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Sun, H.X.; Shu, Y.H.; Tang, W.C.; Wang, Q.; Zhou, Q.; Zhang, G.R. Nickel accumulation and its effects on the survival rate of Spodoptera litura Fabricius under continuous nickel stress. Chin. Sci. Bull. 2007, 14, 1957–1963. [Google Scholar] [CrossRef]
- Lin, X.D.; Zhang, L.; Jiang, Y.Y. Characterization of Spodoptera litura (Lepidoptera: Noctuidae) takeout genes and their differential responses to insecticides and sex pheromone. J. Insect Sci. 2017, 17, 81–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, H.; Qian, Y.; Peng, B.Z.; Jiang, X.L.; Hua, X.M. Environmental pesticide pollution and its countermeasures in China. AMBIO A J. Hum. Environ. 2003, 32, 78–80. [Google Scholar]
- Wilson, E.O.; Bossert, W.H. Chemical communication; among animals. Recent Prog. Horm. Res. 1963, 19, 673–716. [Google Scholar]
- Li, Y.; Zeng, X.N.; Wang, R.X.; Jiang, J.Q.; Ma, F.N. Influence of semiochemicals on insect behevior and their application in pest management. Guangdong Agric. Sci. 2008, 7, 85–89. [Google Scholar]
- Tamaki, Y.; Yushima, T. Biological activity of the synthesized sex pheromone and its geometrical isomers of Spodoptera litura (F.) (Lepidoptera: Noctuidae). Appl. Ent. Zool. 1974, 9, 73–79. [Google Scholar] [CrossRef] [Green Version]
- Yusunvm, T.; Noguchi, H.; Tamaki, Y.; Fukazawa, N.; Sugino, T. Mating and sex pheromone of Spodoptera litura F. (Lepidoptera: Noctuidae): An introductory report. Appl. Entomol. Zool. 1973, 8, 18–26. [Google Scholar]
- Kawasaki, K. Electroantennogram responses of Spodoptera litura (F.) (Lepidoptera: Noctuidae) male moth to two female sex pheromone components. Appl. Ent. Zool. 1985, 20, 82–87. [Google Scholar] [CrossRef] [Green Version]
- Xu, G.Q.; Cai, Z.J.; Liu, P.B. Studies and application of sex pheromone production in beet armyworm, Spodoptera exigua. Chin. J. Appl. Entomol. 2008, 3, 357–361. [Google Scholar]
- Le, J.M.; Chen, Y.; Deng, R.J.; Ding, H.B.; Yang, H.; Zeng, X.H.; Cai, L. Study on dynamic regularity of Spodoptera litura Fabricius by insect sex pheromone monitoring in Guiyang area. Jiangsu Agric. Sci. 2019, 47, 75–78. [Google Scholar]
- Yi, L.; Tian, J.L.; Qiu, M.W.; Chen, Y.M.; Ye, Y.L.; Deng, H.B. Densities and types of traps against Spodoptera litura: Trapping and control effects in tobacco field. Chin. Agric. Sci. Bull. 2018, 34, 147–151. [Google Scholar]
- Hu, Z.Z.; Shi, Y.; Li, Y.H.; Xve, J.G.; Wang, W.B.; Qian, H.T.; Xing, G.N.; Gai, J.Y. Study on the trapping effect of different aggregation pheromone lure cores and trap devices in soybean field. Soybean Sci. 2020, 39, 288–296. [Google Scholar]
- Zhang, J.M.; Lin, W.C.; Lv, Y.B.; Bei, Y.W.; Li, W.D.; Zhang, Z.J.; Zhang, W.; Yao, H. Comparison of trapping effects of different types of traps on male Spodoptera litura. J. Zhejiang Agric. Sci. 2008, 4, 475–477. [Google Scholar]
- Robert, L.; Meagher, J.B.; Edward, H. Monitoring for exotic Spodoptera species (Lepidoptera: Noctuidae) in Florida. Fla. Entomol. 2008, 91, 517–522. [Google Scholar]
- Ranga, R.G.V.; Wightman, J.A.; Ranga, R.D.V. The development of a standard pheromone trapping procedure for Spodoptera Iitura (F) (Lepidoptera: Noctuidae) population in groundnut (Arachis hyopogaea L) crops. Trop. Pest Manag. 1991, 37, 37–40. [Google Scholar]
- Wan, X.; Deng, J.Y.; Wang, Y.P. Effect of different trap types and fluorescent tubes at different bands on capture of Hyphantria cunea (Drury). Plant Prot. 2021, 47, 103–107. [Google Scholar]
- Guo, X.X.; Dai, K.S.; Li, J.; Wang, S.K.; Xu, M.W.; Wang, L. Trapping efficacy of three trapper against Spodoptera litura. J. Zhejiang Agric. Sci. 2021, 62, 1821–1823. [Google Scholar]
- Chen, B.H.; Wen, Y.H.; Chen, C.B.; Yang, Y.H.; Shen, J.M. Comparative test on trapping effects of different sex pheromone lures to Spodoptera litura in Guangzhou. Guangdong Agric. Sci. 2019, 46, 94–98. [Google Scholar]
- Shu, Y.N. Spodoptera litura (Fabricius) in China. Chin. J. Appl. Entomol. 1959, 3, 106–107. [Google Scholar]
- Karakasis, A.; Lampiri, E.; Rumbos, C.I.; Athanassiou, C.G. Factors affecting adult captures of the cotton bollworm, Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae) in pheromone-baited traps. Agronomy 2021, 11, 2539. [Google Scholar] [CrossRef]
- Bailey, J.B.; Mcdonough, L.M.; Hoffmann, M.P. Western avocado leaf roller, Amorbia cuneana (Walsingham), (Lepidoptera Tortricidae): Discovery of populations utilizing different ratios of sex pheromone components. J. Chem. Ecol. 1986, 12, 1239–1245. [Google Scholar] [CrossRef] [PubMed]
- Lofstedt, C.; Hansson, B.S.; Lofovist, J.; Vanderpers, J.N.C.; Hansson, B.S. Pheromone dialects in European turnip moths Agrotis segtum. Oikos 1986, 46, 250–257. [Google Scholar] [CrossRef]
- Wang, Y.C.; Li, L.Y.; Gong, X.J.; Cao, R.Y.; Xiong, Y.Y.; Huang, F.; Luo, F. Experiment on trapping effects of different sex-attractants on Prodenia litura in tea garden. Tianjin Agric. Sci. 2019, 25, 81–83. [Google Scholar]
- Teng, H.Y.; Wang, D.S.; Zhang, Q.L. Evaluation of trapping effects of different sex pheromone products on Spodoptera exigua and Spodoptera litura in Shanghai. Acta Agric. Shanghai 2015, 31, 40–43. [Google Scholar]
- Li, X. The Pest Monitoring System Based on the Internet of Things. Master’s Thesis, Beijing Forestry University, Beijing, China, 2019. [Google Scholar]
- Duraimurugan, P. Effect of weather parameters on the seasonal dynamics of tobacco caterpillar, S. litura (Lepidoptera: Noctuidae) in castor in Telangana State. J. Agrometeorol. 2018, 20, 139–143. [Google Scholar] [CrossRef]
- Menéndez, R.; González-Megías, A.; Collingham, Y.; Fox, R.; Roy, D.B.; Ohlemuller, R.; Thomas, C.D. Direct and indirect effects of climate and habitat factors on butterfly diversity. Ecology 2007, 88, 605–611. [Google Scholar] [CrossRef]
- Wu, C.X. The Research on Behavior Rhythm and Sexual Selection of Spodoptera litura (Fabricius) (Lepidoptera: Noctuidae) Adults. Master’s Thesis, Guizhou University, Guiyang, China, 2015. [Google Scholar]
- Fu, X.W. Study on the Community Structure and Population Dynamics of Migratory Insects across the Bohai Strait. Ph.D. Thesis, Chinese Academy of Agricultural Sciences, Beijing, China, 2015. [Google Scholar]
- Liu, P.C.; Diao, Y.H.; Guo, W.J.; Gao, B.Y.; Hu, G. Insect migration behavior and its regulation. Chin. J. Appl. Entomol. 2021, 58, 520–529. [Google Scholar]
- Tojo, S.; Ryuda, M.; Fukuda, T.; Matsunaga, T.; Choi, D.R.; Otuka, A. Overseas migration of the common cutworm, S. litura (Lepidoptera: Noctuidae), from May to mid-July in East Asia. Appl. Entomol. Zool. 2013, 48, 131–140. [Google Scholar] [CrossRef]
- Wu, H.H.; Huang, M.S.; Lei, C.L. The spatial-temporal distribution of Spodoptera litura in China. J. Anhui Agric. Sci. 2016, 44, 142–144. [Google Scholar]
- Wu, H.H. Study on Trapped Dynamics and Genetic Structure among Different Geographic Populations of Spodoptera litura. Ph.D. Thesis, Huazhong Agricultural University, Wuhan, China, 2018. [Google Scholar]
- Zeng, A.P.; Chen, Y.N.; Zhang, Q.; Hu, R.S.; Zhou, Z.C.; Long, J.Z.; Li, X.Y.; Wu, C.E. Occurrence pattern of Spodoptera litura in Hunan and its prediction methods. Chin. Tob. Sci. 2010, 31, 9–13. [Google Scholar]
- Chen, H. Current situation and trend of utilization pheromones for control of bark beetles. J. Northwest For. Univ. 2002, 17, 60–63. [Google Scholar]
- Ross, D.W.; Niwa, C.G. Using aggregation and antiaggregation pheromones of the Douglas-fir beetle to produce snags for wildlife habitat. West. J. Appl. For. 1997, 12, 52–54. [Google Scholar] [CrossRef] [Green Version]
- Punithavalli, M.; Sharma, A.N.; Balaji, R.M. Seasonality of the common cutworm Spodoptera litura in a soybean ecosystem. Phytoparasitica 2014, 42, 213–222. [Google Scholar] [CrossRef]
- Juil, K.; Min, K.; Ki, J.P.; Maharjanm, R. Monitoring of four major lepidopteran pests in Korean cornfields and management of Helicoverpa armigera. Entomol. Res. 2018, 48, 308–316. [Google Scholar]
- Akira, O.; Masaya, M.; Makoto, T. Dispersal of the common cutworm, Spodoptera litura, monitored by searchlight trap and relationship with occurrence of soybean leaf damage. Insects 2020, 11, 427–443. [Google Scholar]
- Rao, M.S.; Manimanjari, D.; Rao, C.A.R.; Maheswari, M. Prediction of pest scenarios of Spodoptera litura Fab. in peanut growing areas of India during future climate change. Natl. Acad. Sci. Lett. 2015, 38, 465–468. [Google Scholar]
- Cui, Z.L.; Gai, J.Y.; Ji, D.F.; Ren, Z.J. Investigation and analysis of soybean leaf eating pests in Nanjing. Soybean Sci. 1997, 16, 12–20. [Google Scholar]
- Hu, Z.Z.; Xu, X.C.; Pan, L.; Li, M.; Zeng, J.; Muhammad, K.R.; Xing, G.N.; Gai, J.Y. Resistance analyses of soybean organs to common cutworm (Spodoptera litura) at different reproductive stages. Soybean Sci. 2020, 39, 932–939. [Google Scholar]
- Xing, G.N.; Liu, K.; Gai, J.Y. A high-throughput phenotyping procedure for evaluation of antixenosis against common cutworm at early seedling stage in soybean. Plant Methods 2017, 13, 66. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type | Outer Entrance Diameter (cm) | Inner Entrance Diameter (cm) | Length (cm) | Distance (cm) * | Number of Entrances |
|---|---|---|---|---|---|
| YL-HEMT | 29.5 | 4.0 | 29.00 | 29.0 | 1 |
| YL-NMT | 19.5 | 2.0 | 15.00 | 15.0 | 1 |
| YL-VT | 2.0 | 1.0 | 2.00 | −6.0 | 16 |
| AIM | 2.5*8.0 | 2.5*8.0 | 0.15 | −5.0 | 6 |
| AIM-lite-A | 12.0 | 6.0 | 10.00 | 10.0 | 1 |
| Light Dark Change Time | July | August | September | October |
|---|---|---|---|---|
| Sunrise Time | 4:36→4:55 | 4:55→5:16 | 5:17→5:35 | 5:35→5:56 |
| Sunset Time | 19:43→19:29 | 19:28→18:56 | 18:54→18:16 | 18:15→17:43 |
| Source of Variation | Date Range 1 | Date Range 2 | Date Range 3 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| DF | MS | F | P | DF | MS | F | P | DF | MS | F | P | |
| Trap | 2 | 896,807.6 | 281.6 ** | <0.0001 | 3 | 261,098.2 | 91.2 ** | <0.0001 | 4 | 125,582.6 | 103.9 ** | <0.0001 |
| Block | 4 | 18,227.2 | 5.7 ** | 0.0002 | 4 | 9465.9 | 3.3 * | 0.01 | 4 | 20,735.0 | 17.2 ** | <0.0001 |
| Date | 34 | 24,929.0 | 7.8 ** | <0.0001 | 20 | 9734.5 | 3.4 ** | <0.0001 | 25 | 8496.2 | 7.0 ** | <0.0001 |
| Error | 411 | 3184.9 | 287 | 2862.6 | 428 | 1209.3 | ||||||
| Trap | Date Range 1 | Date Range 2 | Date Range 3 |
|---|---|---|---|
| YL-HEMT | 10.8 b | 1.4 c | 11.8 c |
| YL-NMT | 19.0 b | 3.0 c | 16.5 c |
| YL-VTused | 84.4 a | ||
| YL-VTnew | 139.1 a | 107.8 a | 52.0 b |
| AIM | 25.6 b | 9.7 c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, Y.; Ye, Z.; Xie, Z.; Zhang, D.; Liu, X.; Yan, Y.; Sun, L.; Chen, F.; Gai, J.; Xing, G. Comparison of Trapping Effects of Different Traps and Monitoring the Occurrence Dynamics of Spodoptera litura in Soybean Fields of Dangtu, Anhui Province, China. Agronomy 2023, 13, 47. https://doi.org/10.3390/agronomy13010047
Xu Y, Ye Z, Xie Z, Zhang D, Liu X, Yan Y, Sun L, Chen F, Gai J, Xing G. Comparison of Trapping Effects of Different Traps and Monitoring the Occurrence Dynamics of Spodoptera litura in Soybean Fields of Dangtu, Anhui Province, China. Agronomy. 2023; 13(1):47. https://doi.org/10.3390/agronomy13010047
Chicago/Turabian StyleXu, Yufei, Zhihao Ye, Zhiyuan Xie, Di Zhang, Xiaofeng Liu, Yulu Yan, Lei Sun, Fajun Chen, Junyi Gai, and Guangnan Xing. 2023. "Comparison of Trapping Effects of Different Traps and Monitoring the Occurrence Dynamics of Spodoptera litura in Soybean Fields of Dangtu, Anhui Province, China" Agronomy 13, no. 1: 47. https://doi.org/10.3390/agronomy13010047
APA StyleXu, Y., Ye, Z., Xie, Z., Zhang, D., Liu, X., Yan, Y., Sun, L., Chen, F., Gai, J., & Xing, G. (2023). Comparison of Trapping Effects of Different Traps and Monitoring the Occurrence Dynamics of Spodoptera litura in Soybean Fields of Dangtu, Anhui Province, China. Agronomy, 13(1), 47. https://doi.org/10.3390/agronomy13010047