
Wheat Response and Weed-Suppressive Ability in the Field Application of a Nanoencapsulated Disulfide (DiS-NH2) Bioherbicide Mimic

 ,
,  ,
,  , , ,
, , , 
 and
and 
Abstract
:1. Introduction
2. Materials and Methods
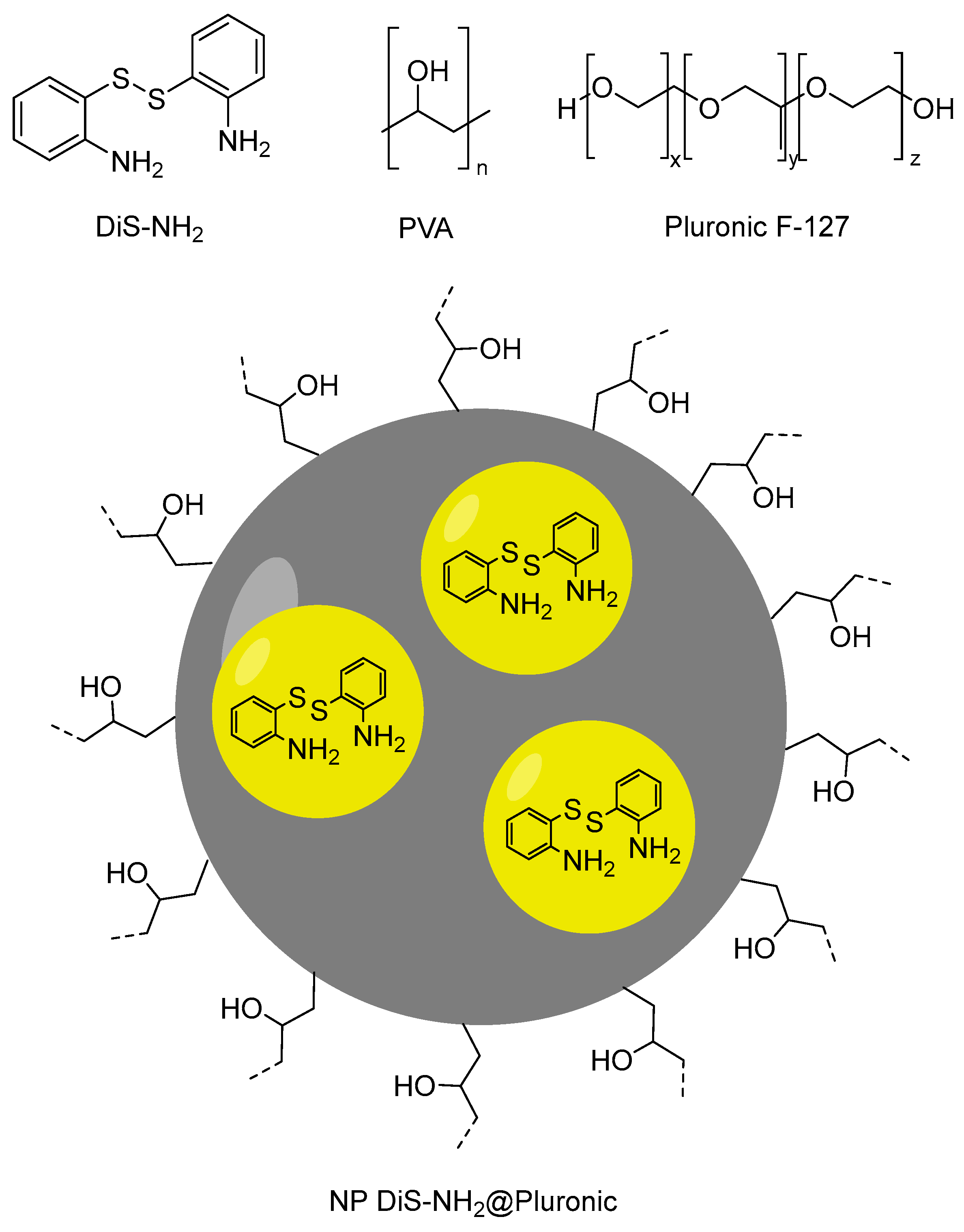
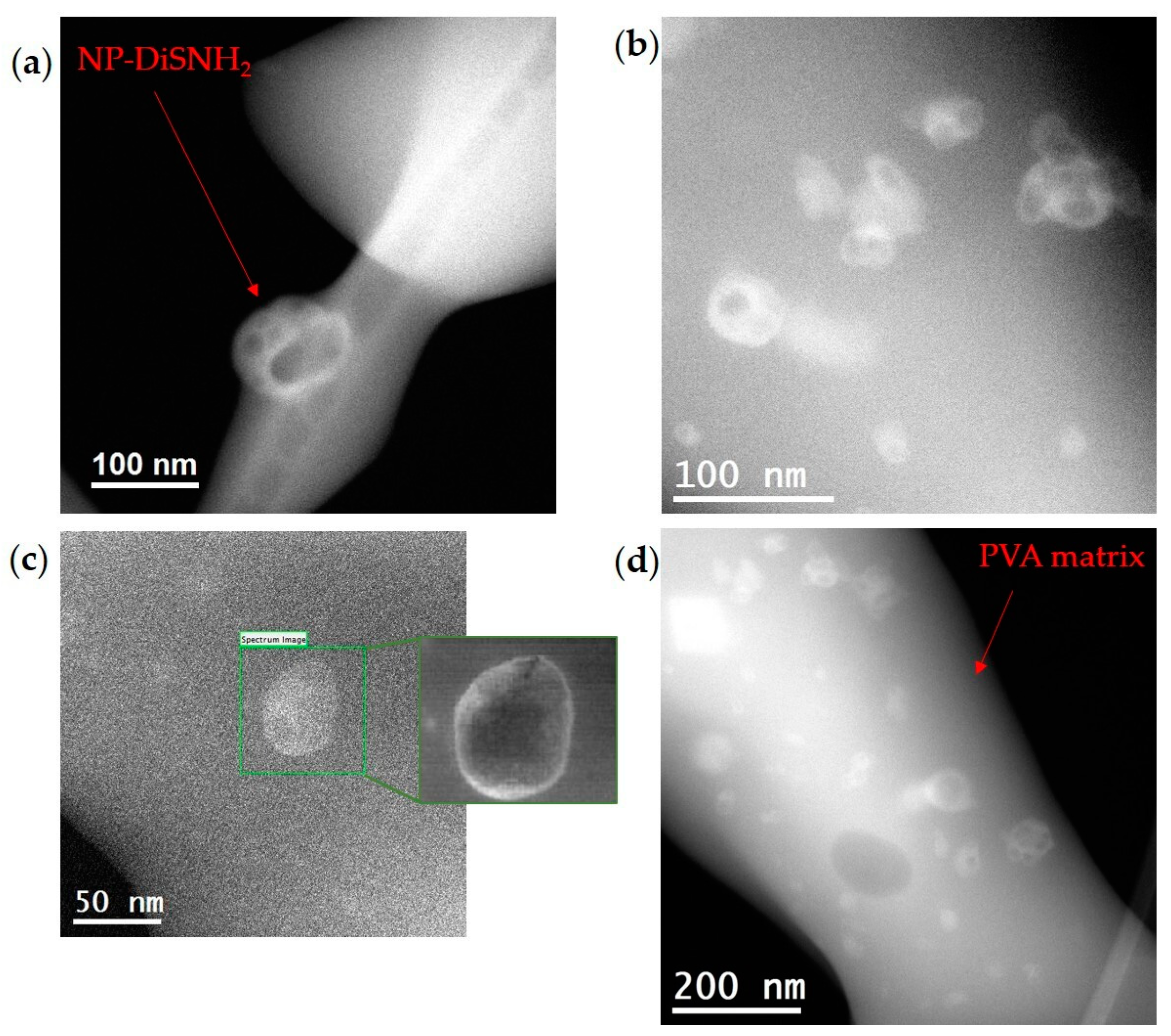
2.1. Formulation of Encapsulated DiS-NH2
2.2. Field Trials and Crop Management
2.3. Measurements
2.3.1. Weed-Suppressive Ability (WSA) of Weedy Treatments
2.3.2. Wheat Morphological, Productive, and Quality Traits
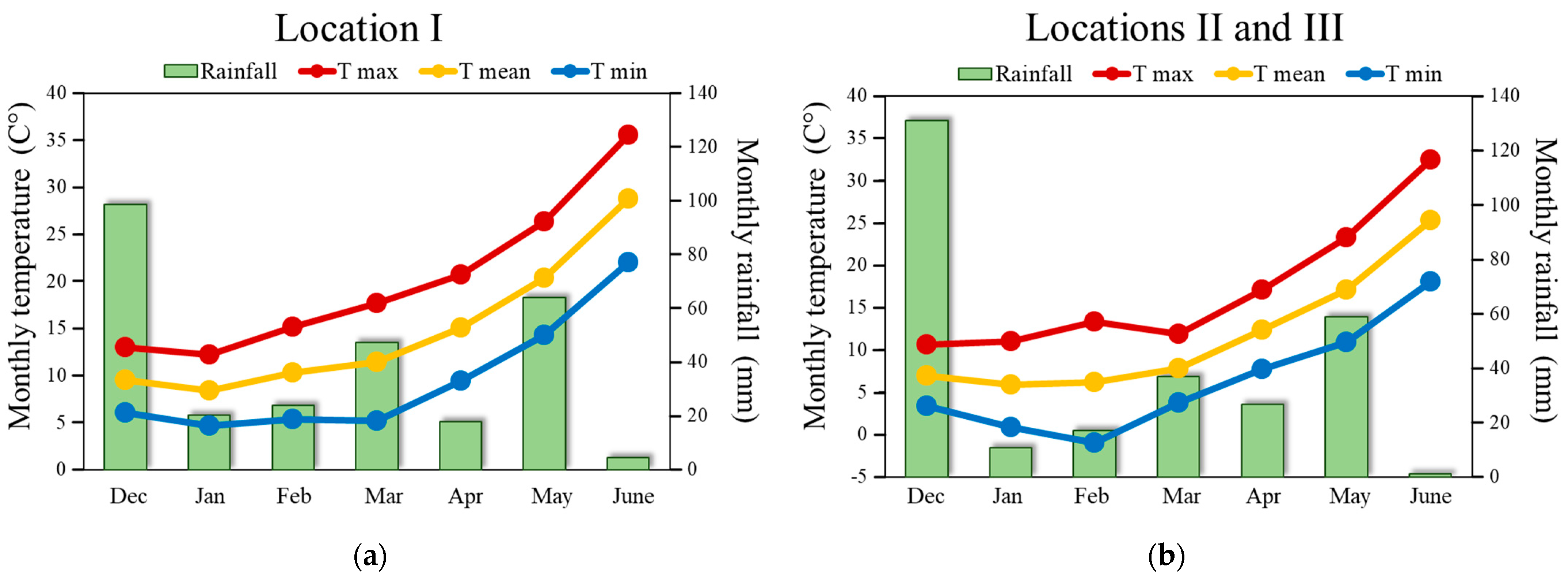
2.4. Climatic Data
2.5. Statistical Analysis
3. Results
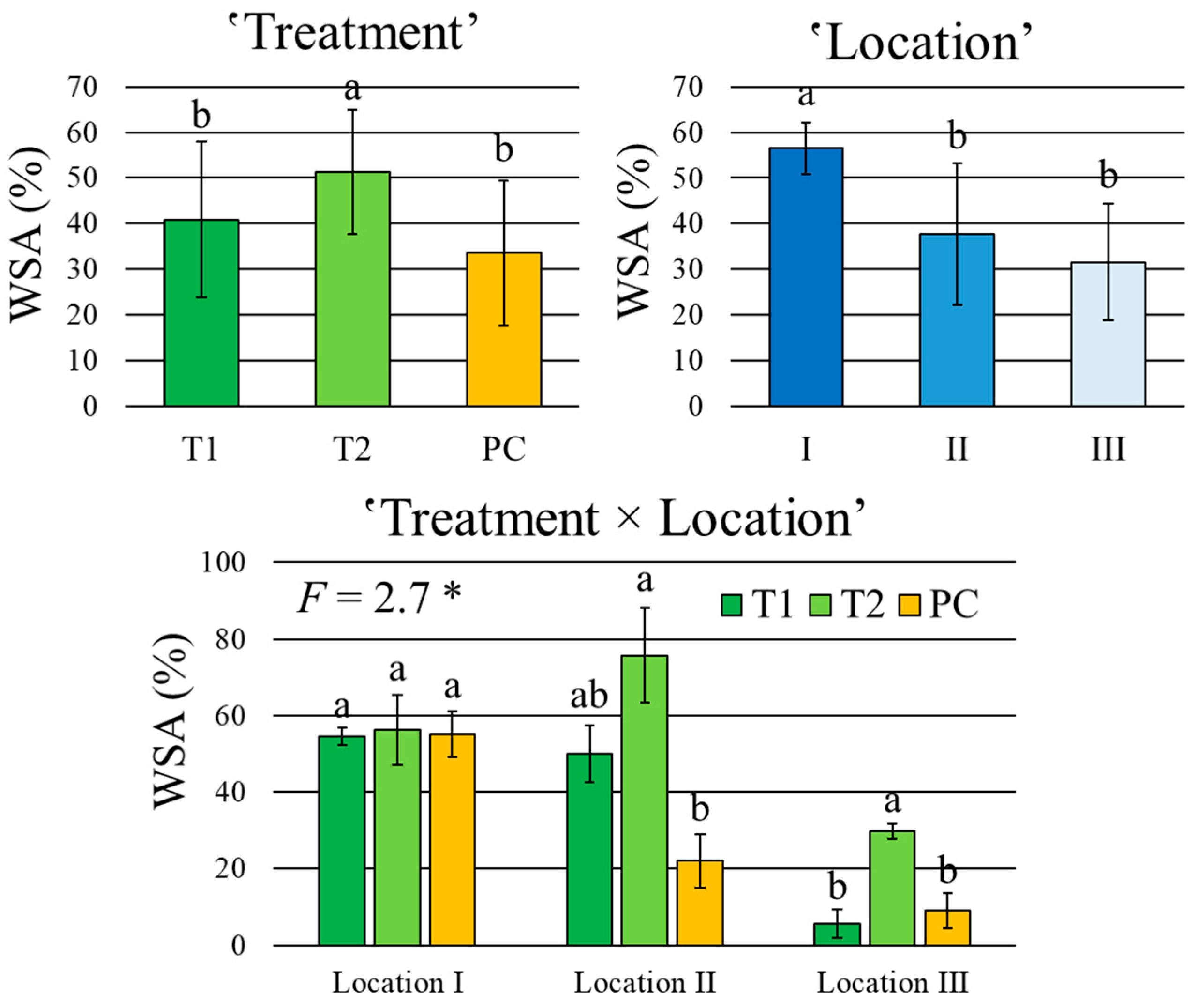
3.1. Weed-Suppressive Ability of Encapsulated DiS-NH2
3.2. Wheat Height
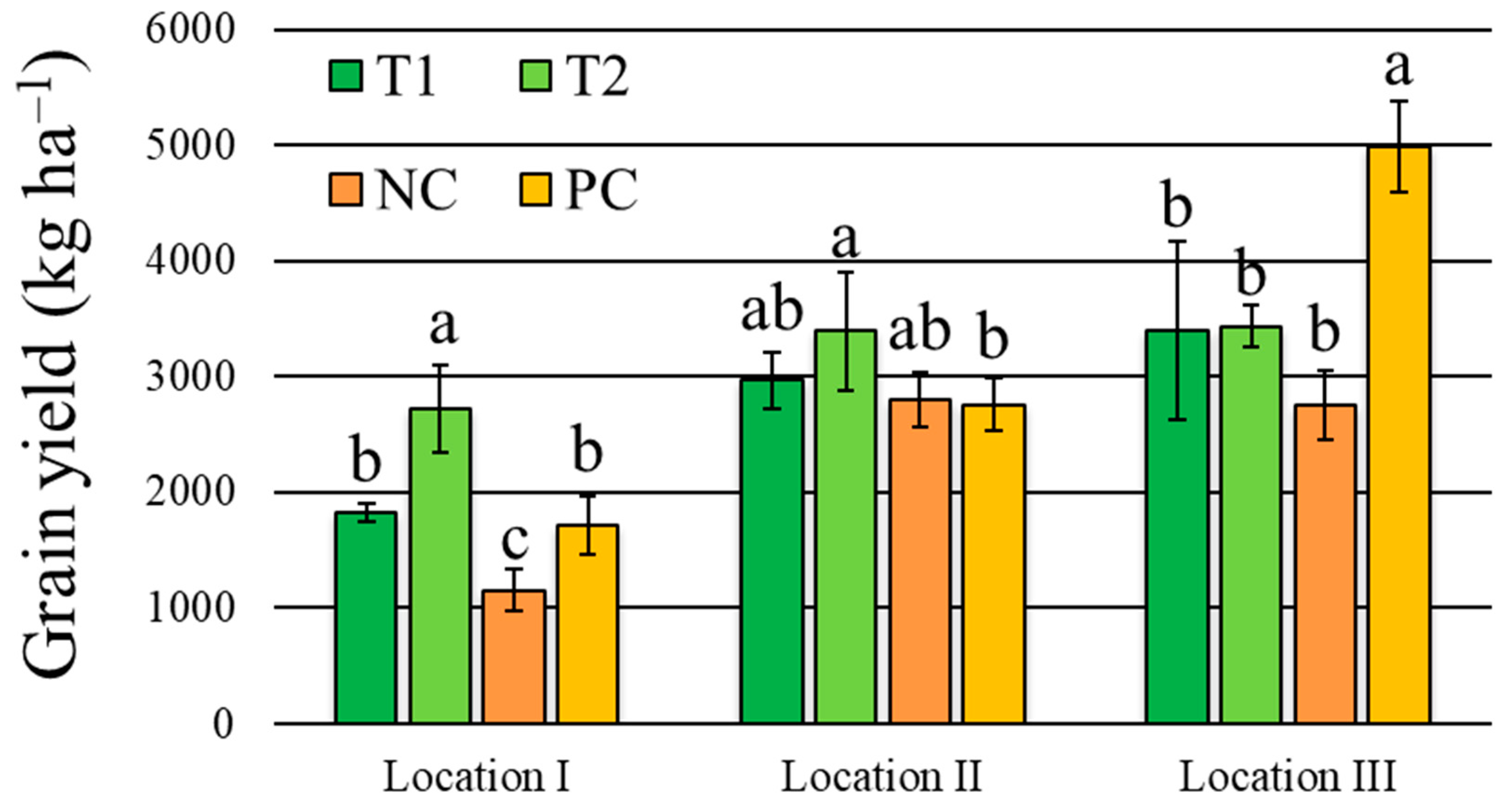
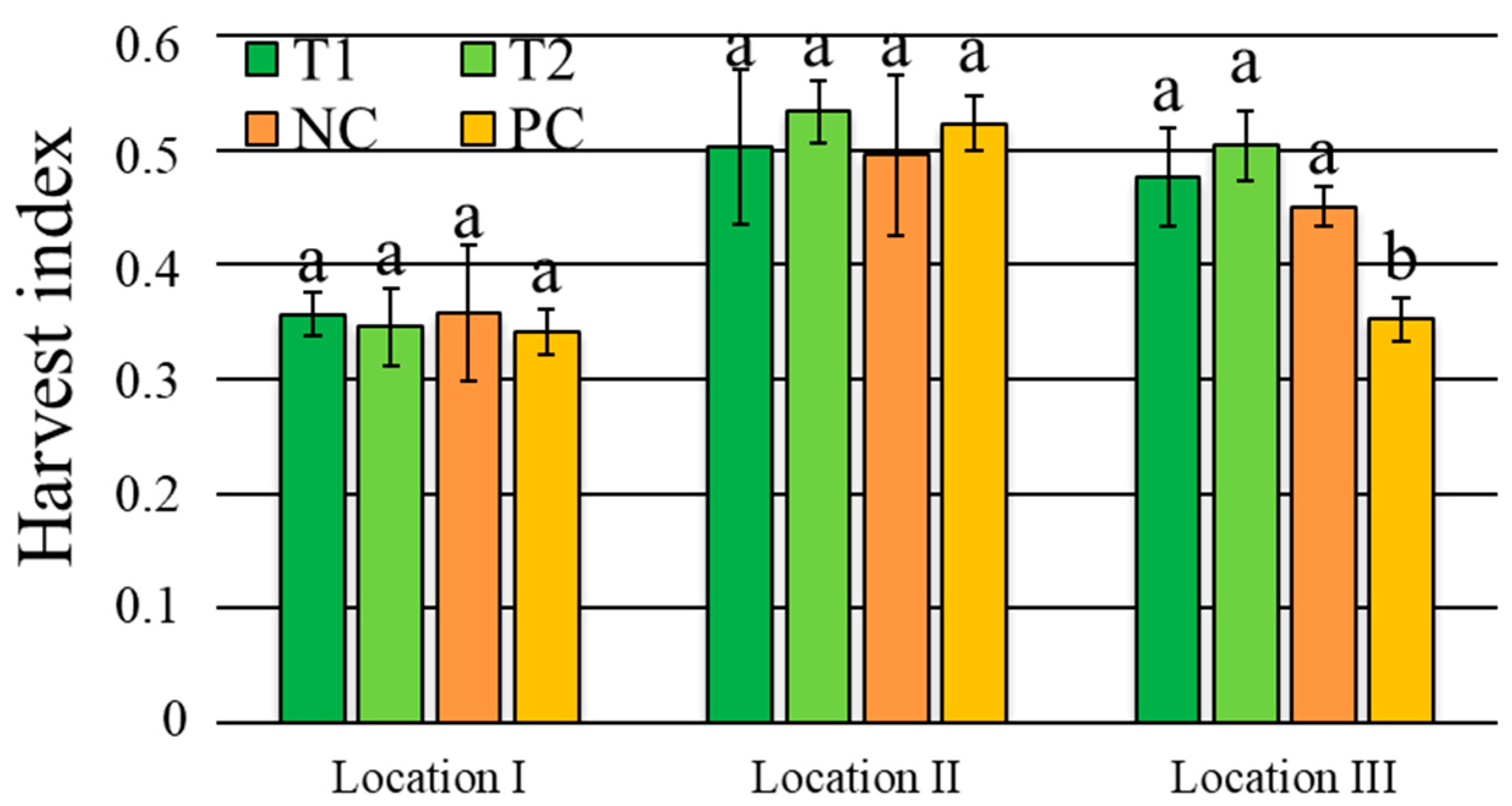
3.3. Wheat Yield and Productive Traits
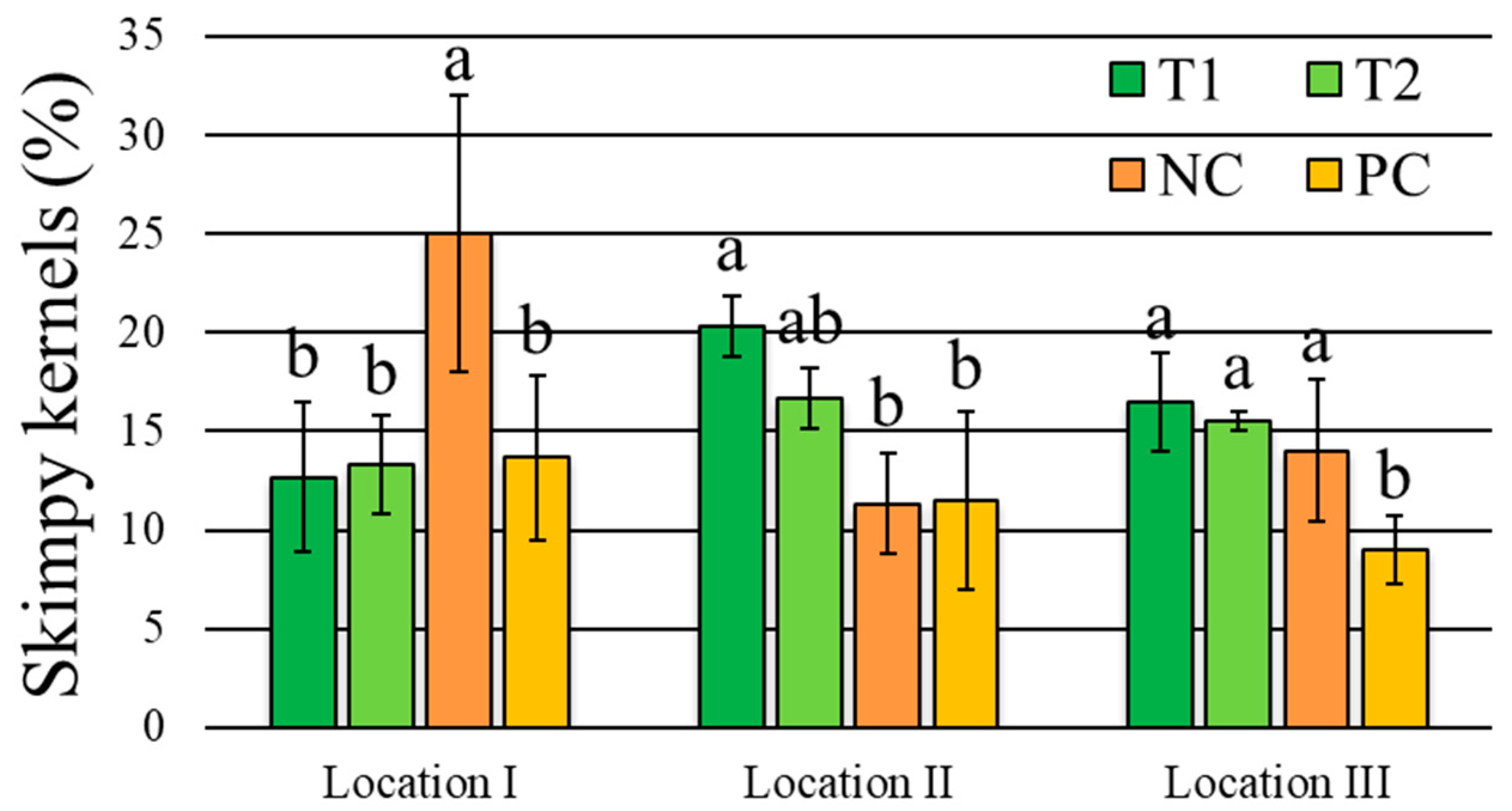
3.4. Wheat Grain Quality Traits
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Faostat. Food and Agriculture Organization of the United Nations 2022. Available online: http://www.fao.org/faostat/en (accessed on 18 February 2023).
- Eurostat. European Commission 2023. Available online: https://ec.europa.eu/eurostat/statistics-explained/index.php?title=Organic_farming_statistics#Organic_production (accessed on 18 February 2023).
- Beres, B.L.; Rahmani, E.; Clarke, J.M.; Grassini, P.; Pozniak, C.J.; Geddes, C.M.; Porker, K.D.; May, W.E.; Ransom, J.K. A Systematic Review of Durum Wheat: Enhancing Production Systems by Exploring Genotype, Environment, and Management (G × E × M) Synergies. Front. Plant Sci. 2020, 11, 568657. [Google Scholar] [CrossRef] [PubMed]
- Scavo, A.; Restuccia, A.; Bannò, M.; Mauromicale, G. Differentiated Weed-Suppressive Ability of Modern and Old Durum Wheat Cultivars after Long-Term Cultivation under Semi-Arid Climate. Plants 2022, 11, 3368. [Google Scholar] [CrossRef] [PubMed]
- Scavo, A.; Mauromicale, G. Integrated Weed Management in Herbaceous Field Crops. Agronomy 2020, 10, 466. [Google Scholar] [CrossRef]
- Piñar Fuentes, J.C.; Leiva, F.; Cano-Ortiz, A.; Musarella, C.M.; Quinto-Canas, R.; Pinto-Gomes, C.J.; Cano, E. Impact of Grass Cover Management with Herbicides on Biodiversity, Soil Cover and Humidity in Olive Groves in the Southern Iberian. Agronomy 2021, 11, 412. [Google Scholar] [CrossRef]
- Hulme, P.E. Weed resistance to different herbicide modes of action is driven by agricultural intensification. Field Crops Res. 2023, 292, 108819. [Google Scholar] [CrossRef]
- Scavo, A.; Pandino, G.; Restuccia, A.; Mauromicale, G. Leaf extracts of cultivated cardoon as potential bioherbicide. Sci. Hortic. 2020, 261, 109024. [Google Scholar] [CrossRef]
- Hoagland, R.E.; Boyette, C.D.; Weaver, M.A.; Abbas, H.K. Bioherbicides: Research and risks. Toxin Rev. 2007, 26, 313–342. [Google Scholar] [CrossRef]
- Scavo, A.; Mauromicale, G. Crop Allelopathy for Sustainable Weed Management in Agroecosystems: Knowing the Present with a View to the Future. Agronomy 2021, 11, 2104. [Google Scholar] [CrossRef]
- Hussain, M.I.; Araniti, F.; Schulz, M.; Baerson, S.; Vieites-Álvarez, Y.; Rempelos, L.; Bilsborrow, P.; Durán, A.G.; Chinchilla, N.; Macías, F.A.; et al. Benzoxazinoids in wheat allelopathy—From discovery to application for sustainable weed management. Environ. Exp. Bot. 2022, 204, 105096. [Google Scholar] [CrossRef]
- Friebe, A. Role of benzoxazinones in cereals. J. Crop Prod. 2001, 4, 379–400. [Google Scholar] [CrossRef]
- Oliveira, S.C.C.; Andrade, C.K.Z.; Varela, R.M.; Molinillo, J.M.G.; Macías, F.A. Phytotoxicity Study of Ortho-Disubstituted Disulfides and Their Acyl Derivatives. ACS Omega 2019, 4, 2362–2368. [Google Scholar] [CrossRef]
- Mejías, F.J.R.; Carrasco, Á.; Durán, A.G.; Molinillo, J.M.G.; Macías, F.A.; Chinchilla, N. On the Formulation of Disulfide Herbicides Based on Aminophenoxazinones: Polymeric Nanoparticle Formulation and Cyclodextrin Complexation to Combat Crop Yield Losses. Pest Manag. Sci. 2022, 79, 1547–1556. [Google Scholar] [CrossRef] [PubMed]
- Macías, F.A.; Mejías, F.J.R.; Molinillo, J.M.G. Recent Advances in Allelopathy for Weed Control: From Knowledge to Applications. Pest Manag. Sci. 2019, 75, 2413–2436. [Google Scholar] [CrossRef] [PubMed]
- Mejías, F.J.R.; Fernández, I.P.; Rial, C.; Varela, R.M.; Molinillo, J.M.G.; Calvino, J.J.; Trasobares, S.; Macías, F.A. Encapsulation of Cynara Cardunculus Guaiane-Type Lactones in Fully Organic Nanotubes Enhances Their Phytotoxic Properties. J. Agric. Food Chem. 2022, 70, 3644–3653. [Google Scholar] [CrossRef] [PubMed]
- Mejías, F.J.R.; López-Haro, M.; Gontard, L.C.; Cala, A.; Fernández-Aparicio, M.; Molinillo, J.M.G.; Calvino, J.J.; Macías, F.A. A Novel Electron Microscopic Characterization of Core/Shell Nanobiostimulator Against Parasitic Plants. ACS Appl. Mater. Interfaces 2018, 10, 2354–2359. [Google Scholar] [CrossRef] [PubMed]
- Soil Survey Staff. Soil Taxonomy: A Basic System of Soil Classification for Making and Interpreting Soil Surveys, 2nd ed.; U.S. Government Publishing Office: Washington, DC, USA, 1999. [Google Scholar]
- Lancashire, P.D.; Bleiholder, H.; van den Boom, T.; Langeluddeke, P.; Stauss, R.; Weber, E.; Witzenberger, A. A Uniform Decimal Code for Growth Stages of Crops and Weeds. Ann. Appl. Biol. 1991, 119, 561–601. [Google Scholar] [CrossRef]
- Scavo, A.; Pandino, G.; Restuccia, A.; Caruso, P.; Lombardo, S.; Mauromicale, G. Allelopathy in Durum Wheat Landraces as Affected by Genotype and Plant Part. Plants 2022, 11, 1021. [Google Scholar] [CrossRef] [PubMed]
- Restuccia, A.; Scavo, A.; Lombardo, S.; Pandino, G.; Fontanazza, S.; Anastasi, U.; Abbate, C.; Mauromicale, G. Long-Term Effect of Cover Crops on Species Abundance and Diversity of Weed Flora. Plants 2020, 9, 1506. [Google Scholar] [CrossRef]
- Saito, K.; Futakuchi, K. Improving estimation of weed suppressive ability of upland rice varieties using substitute weeds. Field Crops Res. 2014, 162, 1–5. [Google Scholar] [CrossRef]
- Sabella, E.; Aprile, A.; Negro, C.; Nicolì, F.; Nutricati, E.; Vergine, M.; Luvisi, A.; De Bellis, L. Impact of Climate Change on Durum Wheat Yield. Agronomy 2020, 10, 793. [Google Scholar] [CrossRef]
- Scavo, A.; Restuccia, A.; Abbate, C.; Lombardo, S.; Fontanazza, S.; Pandino, G.; Anastasi, U.; Mauromicale, G. Trifolium subterraneum cover cropping enhances soil fertility and weed seedbank dynamics in a Mediterranean apricot orchard. Agron. Sustain. Dev. 2021, 41, 70. [Google Scholar] [CrossRef]
- Díaz-Franco, C.; Rial, C.; Molinillo, J.M.G.; Varela, R.M.; Macías, F.A. Synthesis of Aminophenoxazinones and Evaluation of Their Phytotoxicity in the Search for New Natural Herbicides. Agronomy 2023, 13, 568. [Google Scholar] [CrossRef]
- Venturelli, S.; Belz, R.G.; Kämper, A.; Berger, A.; von Horn, K.; Wegner, A.; Böcker, A.; Zabulon, G.; Langenecker, T.; Kohlbacher, O.; et al. Plants Release Precursors of Histone Deacetylase Inhibitors to Suppress Growth of Competitors. Plant Cell 2015, 27, 3175–3189. [Google Scholar] [CrossRef]
- De Vita, P.; Di Paolo, E.; Fecondo, G.; Di Fonzo, N.; Pisante, M. No-tillage and conventional tillage effects on durum wheat yield, grain quality and soil moisture content in southern Italy. Soil Till. Res. 2007, 92, 69–78. [Google Scholar] [CrossRef]
- Quaranta, F.; Belocchi, A.; Fornara, M.; Ripa, C.; D’Egidio, M.G. Le Varietà di Frumento Duro in Italia: Risultati Della Rete Nazionale di Sperimentazione 1999–2012; Council for Agricultural Research and Economics: Rome, Italy, 2013. [Google Scholar]
- Jannink, J.L.; Orf, J.H.; Jordan, N.R.; Shaw, R.G. Index selection for weed suppressive ability in soybean. Crop Sci. 2000, 40, 1087–1094. [Google Scholar] [CrossRef]
- Zhao, D.L.; Atlin, G.N.; Bastiaans, L.; Spiertz, J.H.J. Cultivar weed-competitiveness in aerobic rice: Heritability, correlated traits, and the potential for indirect selection. Crop Sci. 2006, 46, 372–380. [Google Scholar] [CrossRef]
- Prystupa, P.; Savin, R.; Slafer, G.A. Grain number and its relationship with dry matter, N and P in the spikes at heading in response to N×P fertilization in barley. Field Crops Res. 2004, 90, 245–254. [Google Scholar] [CrossRef]
- García del Moral, L.F.; Rharrabti, Y.; Villegas, D.; Royo, C. Evaluation of Grain Yield and Its Components in Durum Wheat under Mediterranean Conditions: An Ontogenic Approach. Agron. J. 2003, 95, 266–274. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | Source of Variation | CV | |||
|---|---|---|---|---|---|
| Blocks | Location (L) | Treatment (T) | (L) × (T) | ||
| Degrees of freedom | 2 | 2 | 3 | 6 | |
| Weed-suppressive ability | 0.3 ns | 9.4 *** | 4.5 * | 2.7 * | 42.8% |
| Plant height | 0.1 ns | 219.6 *** | 7.2 ** | 2.4 ns | 3.6% |
| Grain yield | 0.9 ns | 75.9 *** | 13.6 *** | 9.3 *** | 12.6% |
| Spikes m−2 | 0.9 ns | 7.6 ** | 4.8 * | 2.0 ns | 14.4% |
| Kernel spikes−1 | 0.3 ns | 7.6 ** | 10.3 *** | 1.4 ns | 10.0% |
| Thousand kernel weight | 2.0 ns | 13.0 *** | 2.1 ns | 1.5 ns | 7.5% |
| Harvest index | 1.1 ns | 50.4 *** | 3.1 * | 2.9 * | 9.2% |
| Non-vitreous kernels | 0.6 ns | 34.2 *** | 10.8 *** | 2.0 ns | 10.7% |
| Skimpy kernels | 1.6 ns | 1.2 ns | 5.1 ** | 5.7 *** | 11.7% |
| Ph | GY | nS | KS | Tkw | HI | n-VK | SK | |
|---|---|---|---|---|---|---|---|---|
| cm | kg ha−1 | no. m−2 | no. spike−1 | g | ‒‒‒‒‒‒‒‒‒ % ‒‒‒‒‒‒‒‒‒ | |||
| Treatment | ||||||||
| T1 | 66.1 (3.7) a | 2732.2 (361.7) b | 230.7 (50.4) bc | 40.0 (4.1) a | 48.3 (5.4) a | 0.45 (0.1) a | 12.4 (5.3) c | 16.5 (2.6) a |
| T2 | 67.7 (3.8) a | 3185.0 (357.4) a | 249.6 (38.6) ab | 40.6 (4.3) a | 45.2 (3.9) ab | 0.46 (0.1) a | 19.1 (2.9) ab | 15.2 (1.5) a |
| NC | 63.1 (3.8) b | 2236.0 (239.4) c | 209.5 (45.5) c | 32.1 (4.7) c | 46.0 (5.6) ab | 0.43 (0.1) ab | 20.8 (4.1) a | 16.8 (4.4) a |
| PC | 67.7 (4.6) a | 3153.3 (290.2) a | 268.3 (39.6) a | 35.7 (4.8) b | 44.3 (4.4) b | 0.41 (0.1) b | 17.1 (5.6) b | 11.4 (3.5) b |
| Location | ||||||||
| I | 55.4 (2.3) c | 1853.6 (219.8) c | 210.3 (38.6) b | 38.9 (5.4) a | 41.7 (4.4) b | 0.35 (0.0) c | 11.2 (3.9) b | 16.2 (5.7) a |
| II | 67.3 (3.6) b | 2981.2 (306.1) b | 264.9 (49.2) a | 38.7 (3.8) a | 48.6 (4.4) a | 0.51 (0.0) a | 20.5 (4.1) a | 15.0 (4.6) a |
| III | 75.9 (3.5) a | 3645.1 (410.7) a | 243.4 (39.7) a | 33.7 (5.8) b | 47.5 (2.9) a | 0.45 (0.1) b | 20.4 (5.3) a | 13.8 (3.6) a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scavo, A.; Mejías, F.J.R.; Chinchilla, N.; Molinillo, J.M.G.; Schwaiger, S.; Lombardo, S.; Macías, F.A.; Mauromicale, G. Wheat Response and Weed-Suppressive Ability in the Field Application of a Nanoencapsulated Disulfide (DiS-NH2) Bioherbicide Mimic. Agronomy 2023, 13, 1132. https://doi.org/10.3390/agronomy13041132
Scavo A, Mejías FJR, Chinchilla N, Molinillo JMG, Schwaiger S, Lombardo S, Macías FA, Mauromicale G. Wheat Response and Weed-Suppressive Ability in the Field Application of a Nanoencapsulated Disulfide (DiS-NH2) Bioherbicide Mimic. Agronomy. 2023; 13(4):1132. https://doi.org/10.3390/agronomy13041132
Chicago/Turabian StyleScavo, Aurelio, Francisco J. R. Mejías, Nuria Chinchilla, José M. G. Molinillo, Stefan Schwaiger, Sara Lombardo, Francisco A. Macías, and Giovanni Mauromicale. 2023. "Wheat Response and Weed-Suppressive Ability in the Field Application of a Nanoencapsulated Disulfide (DiS-NH2) Bioherbicide Mimic" Agronomy 13, no. 4: 1132. https://doi.org/10.3390/agronomy13041132
APA StyleScavo, A., Mejías, F. J. R., Chinchilla, N., Molinillo, J. M. G., Schwaiger, S., Lombardo, S., Macías, F. A., & Mauromicale, G. (2023). Wheat Response and Weed-Suppressive Ability in the Field Application of a Nanoencapsulated Disulfide (DiS-NH2) Bioherbicide Mimic. Agronomy, 13(4), 1132. https://doi.org/10.3390/agronomy13041132