
Transpiration Efficiency of Some Potato Genotypes under Drought



Abstract
:1. Introduction
2. Materials and Methods
2.1. Glasshouse Experiments, Climatic Conditions, and Genotypes
2.2. Initial Biomass Harvest and Preparation for Dry-Down Treatment
2.3. Transpiration Measurement and Plant Watering
2.4. Normalizing Transpiration
2.5. Transpiration Efficiency
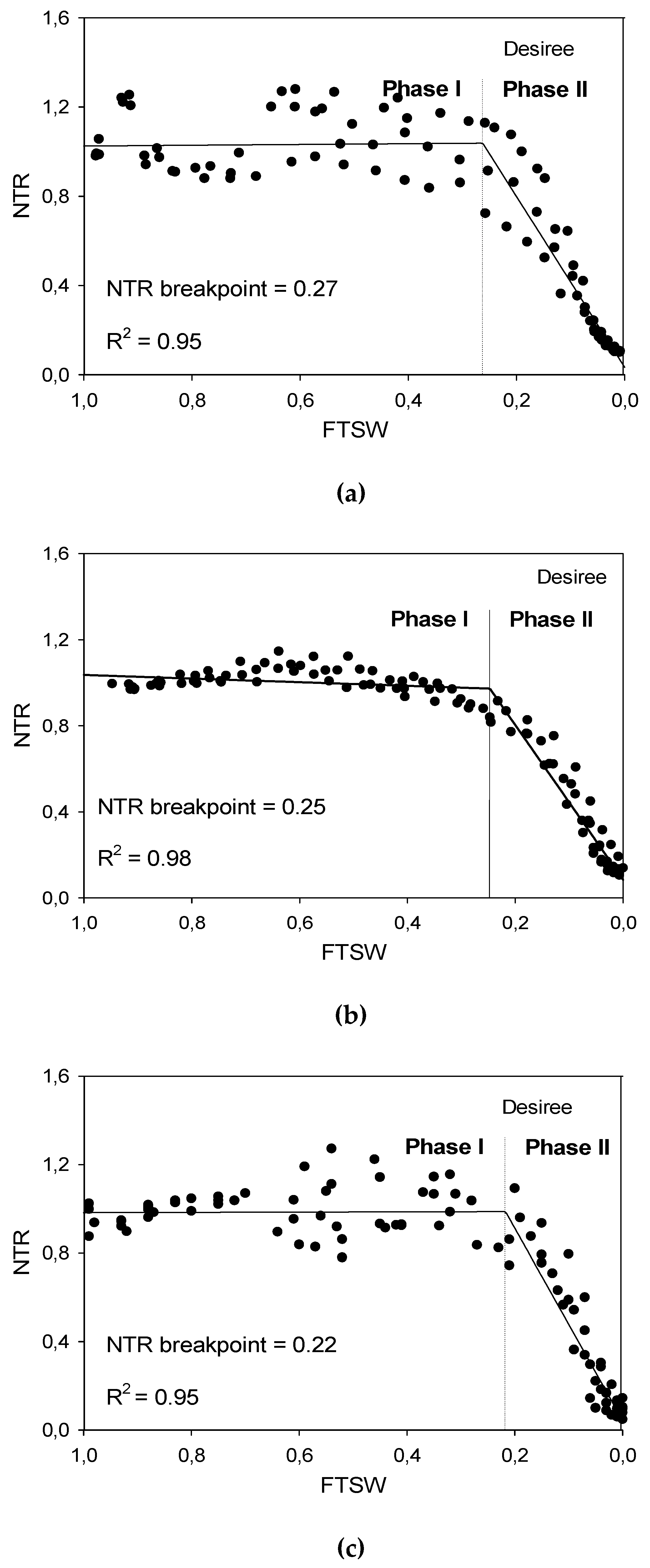
2.6. VPD, FTSW and FTSW Threshold
2.7. Dry Biomass Recording
3. Results
3.1. Transpiration under Different VPD Condition
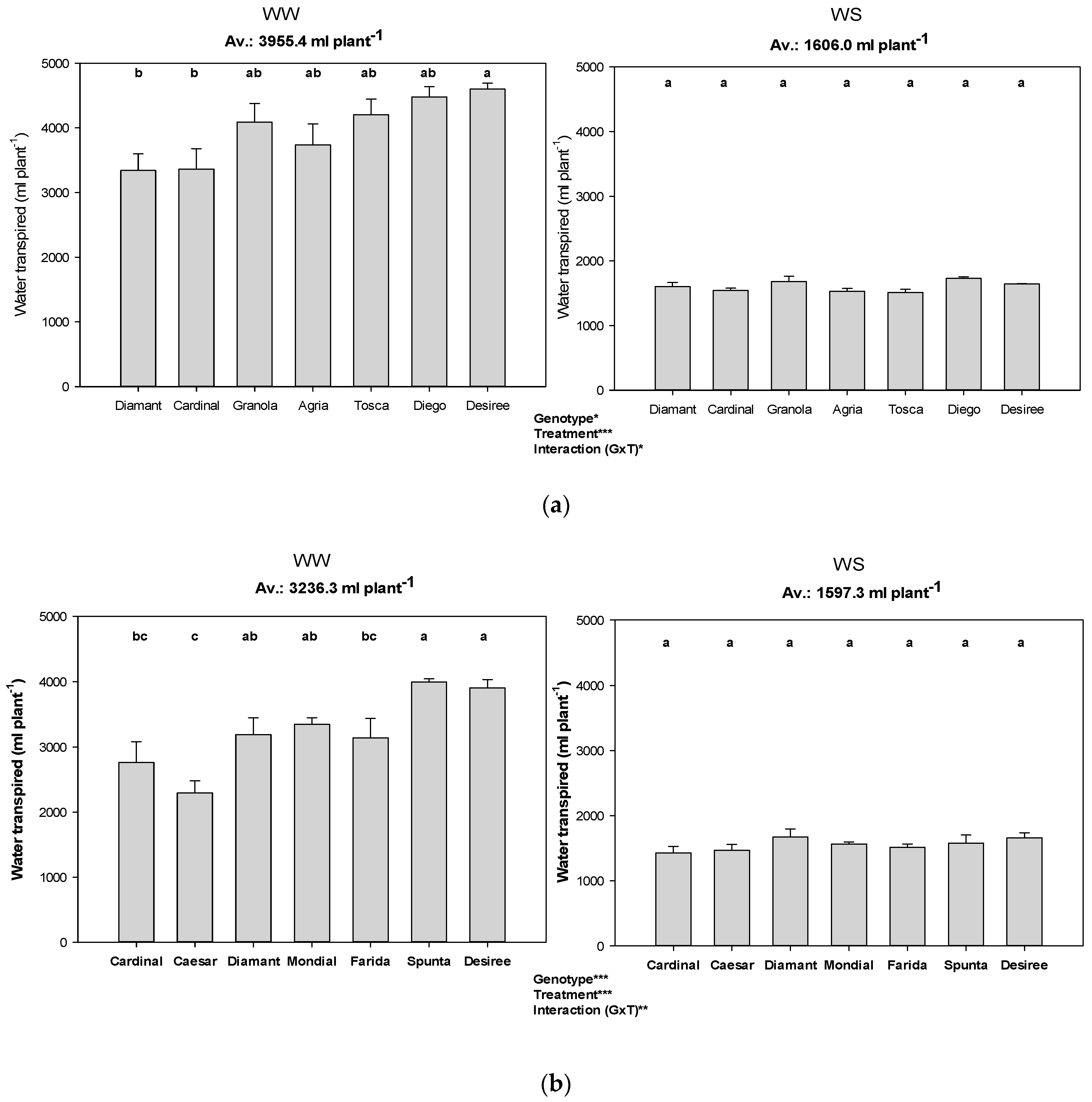
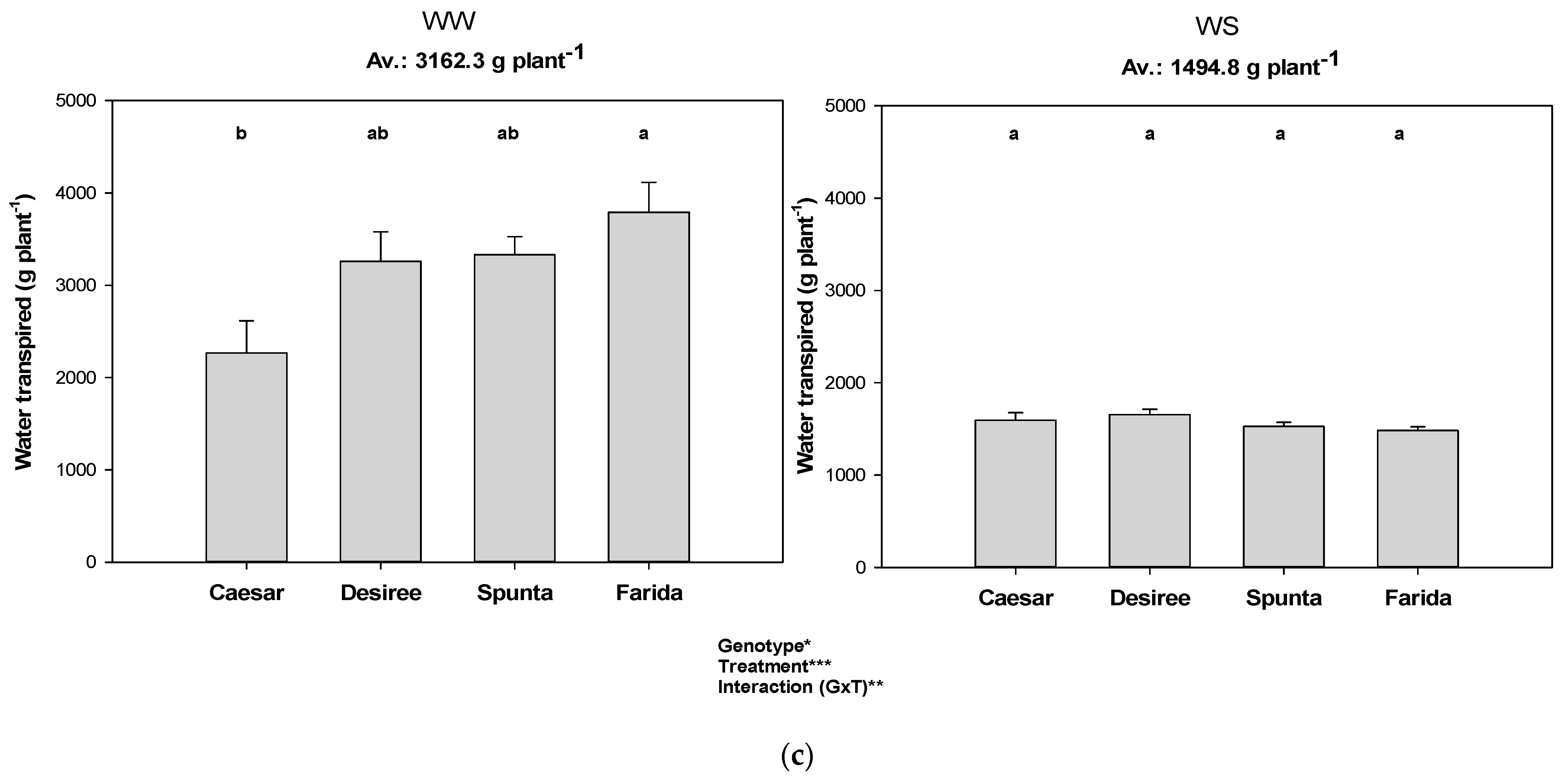
3.2. Transpiration during Soil Drying Cycles
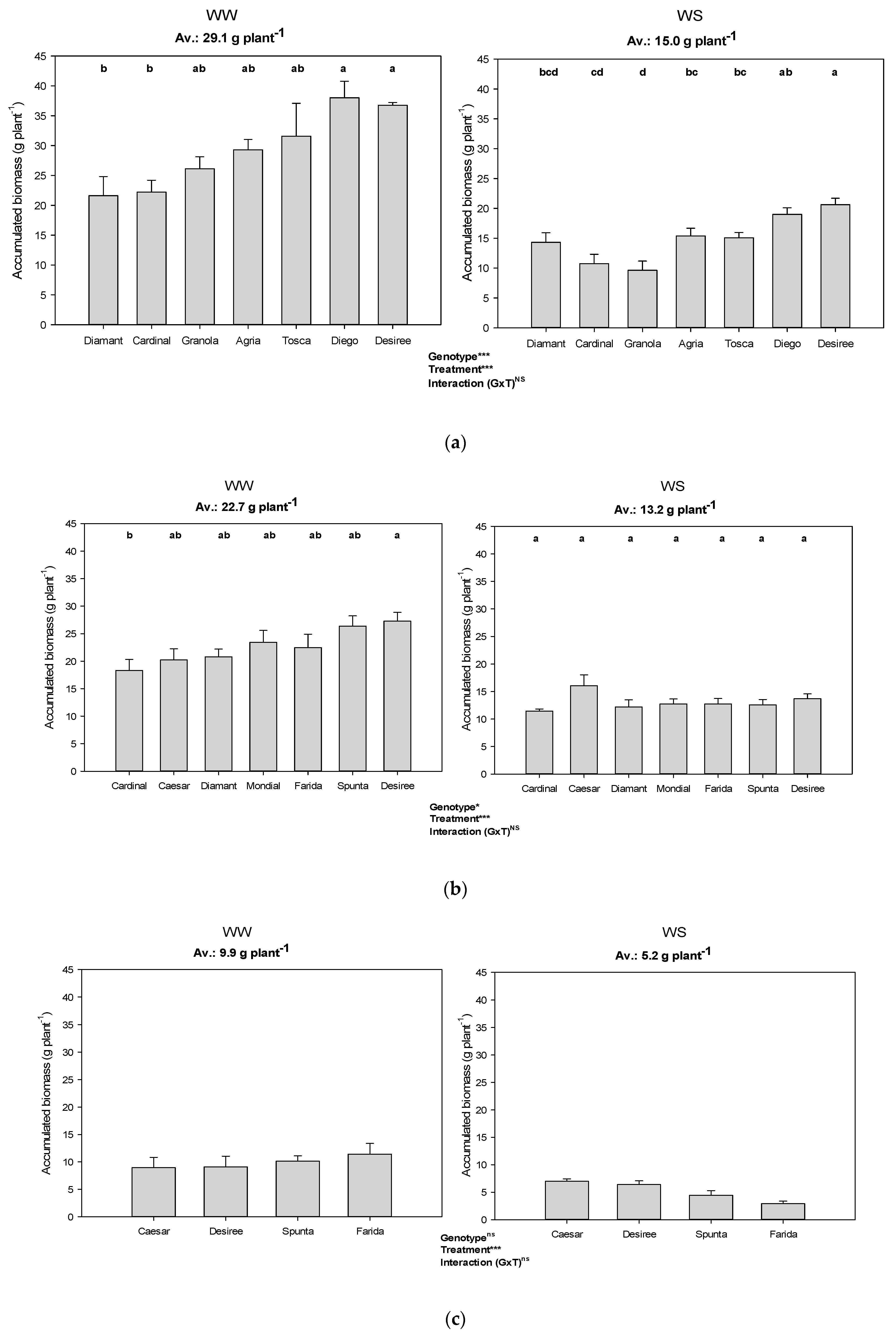
3.3. Biomass Production during the Dry-Down Cycle
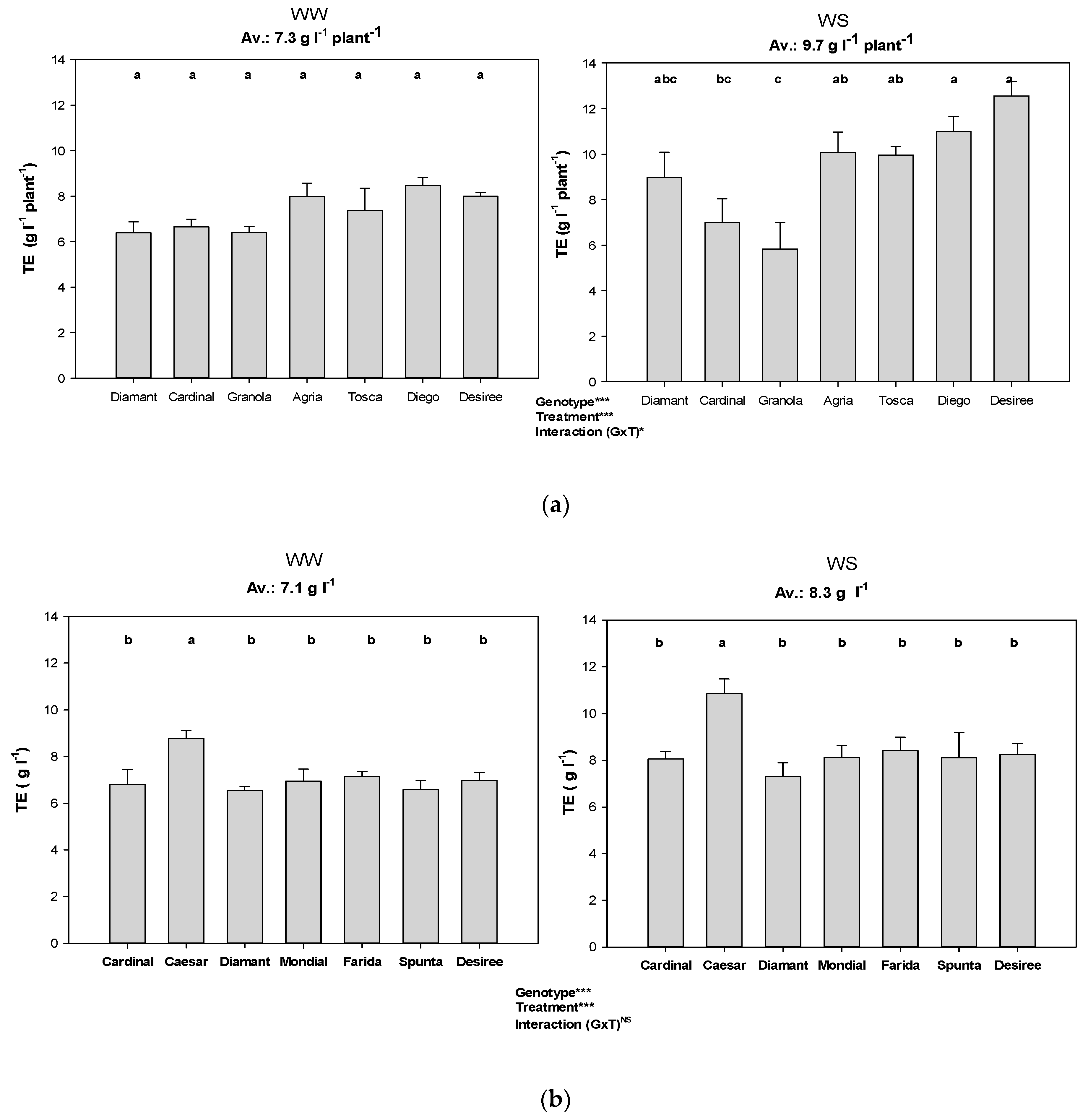
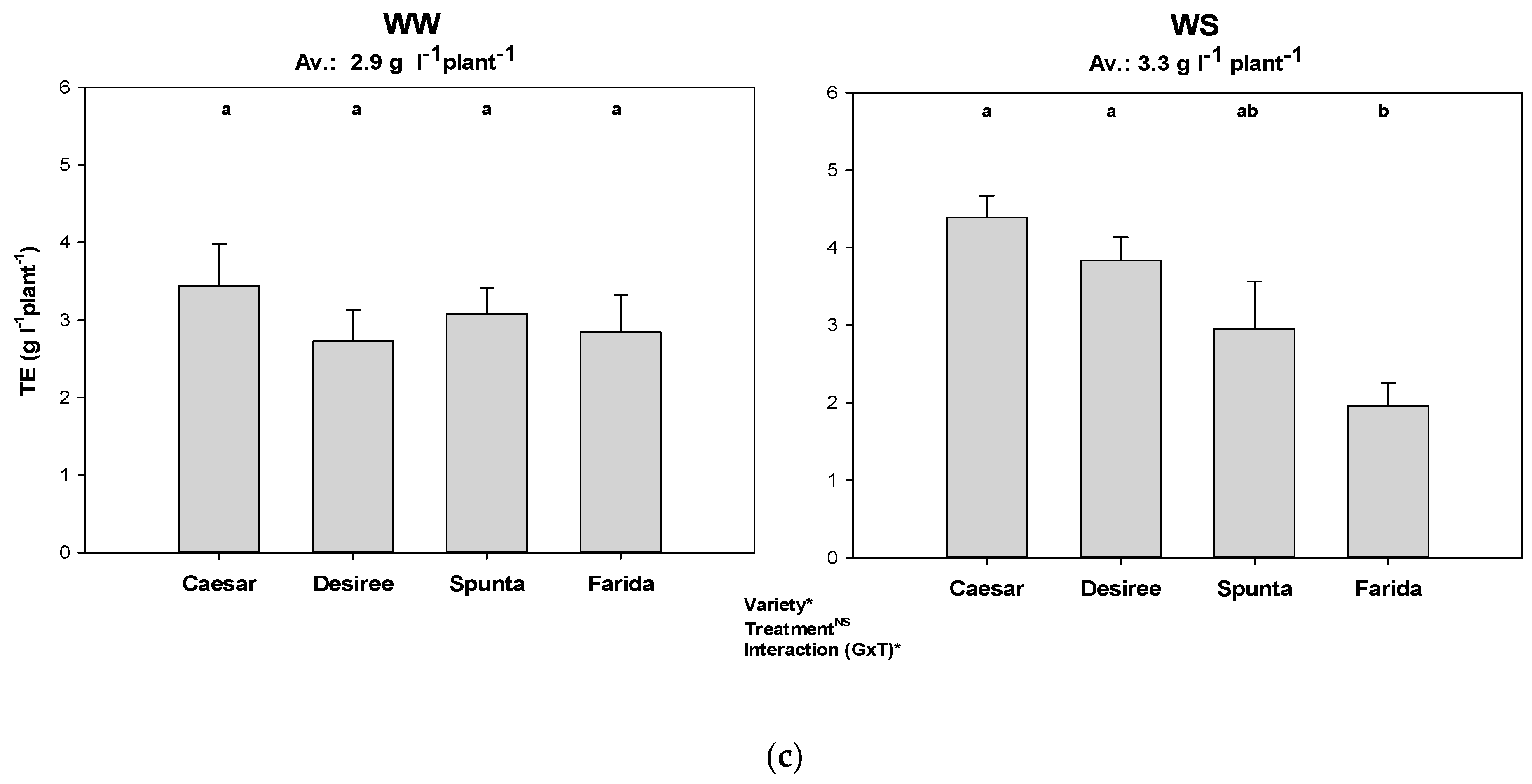
3.4. Transpiration Efficiency Based on Accumulated Biomass
3.5. Relationships between Transpiration and Water Saving Properties of WS Plants
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Lutaladio, N.; Castaldi, L. Potato: The Hidden Treasure. J. Food Compos. Anal. 2009, 22, 491–493. [Google Scholar] [CrossRef]
- Fuqiang, L.; Haoliang, D.; Yucai, W.; Xuan, L.; Xietian, C.; Lintao, L.; Hengjia, Z. Potato Growth, Photosynthesis, Yield, and Quality Response to Regulated Deficit Drip Irrigation under Film Mulching in A Cold and Arid Environment. Res. Sq. 2021, 11, 1–16. [Google Scholar] [CrossRef]
- Muhammad, W.N.; Zoltan, T. Effect of Drought Stress on Potato Production: A Review. Agronomy 2022, 12, 635. [Google Scholar]
- Reddy, B.J.; Mandal, R.; Chakroborty, M.; Hijam, L.; Dutta, P. A Review on Potato (Solanum tuberosum L.) and Its Genet. Diversity. Int. J. Genet. 2018, 10, 360–364. [Google Scholar]
- FAOSTAT. Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on 23 November 2021).
- Battisti, D.S.; Naylor, R.L. Historical Warnings of Future Food Insecurity with Unprecedented Seasonal Heat. Science 2009, 323, 240–244. [Google Scholar] [CrossRef] [Green Version]
- Camargo, D.C.; Montoya, F.; Córcoles, J.I.; Ortega, J.F. Modeling the Impacts of Irrigation Treatments on Potato Growth and Development. Agric. Water Manag. 2015, 150, 119–128. [Google Scholar] [CrossRef]
- Dettinger, M.; Cayan, D.R. Drought and the California Delta—A Matter of Extremes. San Franc. Estuary Watershed. Sci. 2014, 12, 2. [Google Scholar] [CrossRef] [Green Version]
- Wilhite, D.A. Drought-Management Policies and Preparedness Plans: Changing the Paradigm from Crisis to Risk Management in Land Restoration; Elsevier: Amsterdam, The Netherlands, 2016; pp. 443–462. [Google Scholar]
- Deblonde, P.M.K.; Haverkort, A.J.; Ledent, J.-F. Responses of Early and Late Potato Cultivars to Moderate Drought Conditions: Agronomic Parameters and Carbon Isotope Discrimination. Eur. J. Agron. 1999, 11, 91–105. [Google Scholar] [CrossRef]
- Fahad, S.; Bajwa, A.A.; Nazir, U.; Anjum, S.A.; Farooq, A.; Zohaib, A.; Sadia, S.; Nasim, W.; Adkins, S.; Saud, S. Crop Production under Drought, and Heat Stress: Plant Responses and Management Options. Front. Plant Sci. 2017, 8, 1147. [Google Scholar] [CrossRef] [Green Version]
- Hirut, B.; Shimelis, H.; Fentahun, M.; Bonierbale, M.; Gastelo, M.; Asfaw, A. Combining Ability of Highland Tropic Adapted Potato for Tuber Yield and Yield Components under Drought. PLoS ONE 2017, 12, e0181541. [Google Scholar] [CrossRef] [Green Version]
- Boguszewska-Mankowska, D.; Ruszczak, B.; Zarzynska, K. Classification of Potato Varieties Drought Stress Tolerance Using Supervised Learning. Appl. Sci. 2022, 12, 1939. [Google Scholar] [CrossRef]
- Van Loon, C.D. The Effect of Water Stress on Potato Growth, Development, and Yield. Am. Potato J. 1981, 58, 51–69. [Google Scholar] [CrossRef]
- Deblonde, P.M.K.; Ledent, J.F. Effects of Moderate Drought Conditions on Green Leaf Number, Stem Height, Leaf Length and Tuber Yield of Potato Cultivars. Eur. J. Agron. 2001, 14, 31–41. [Google Scholar] [CrossRef]
- Lahlou, O.; Ledent, J.-F. Root Mass and Depth, Stolons and Roots Formed on Stolons in Four Cultivars of Potato under Water Stress. Eur. J. Agron. 2005, 22, 159–173. [Google Scholar] [CrossRef]
- Lahlou, O.; Ouattar, S.; Ledent, J.-F. The Effect of Drought and Cultivar on Growth Parameters, Yieldand Yield Components of Potato. Agronomie 2005, 23, 257–268. [Google Scholar] [CrossRef]
- Kumar, S.; Asrey, R.A.M.; Mandal, G. Effect of Differential Irrigation Regimes on Potato (Solanum tuberosum) Yield and Postharvest Attributes. Indian J. Agric. Sci 2007, 77, 366–368. [Google Scholar]
- Schafleitner, R.; Gutierrez, R.; Legay, S.; Evers, D.; Bonierbale, M. Drought Stress Tolerance Traits of Potato. In Proceedings of the 15th International Symposium of the International Society for Tropical Root Crops (ISTRC), Lima, Peru, 2–7 November 2009; pp. 2–7. [Google Scholar]
- Eiasu, B.K.; Soundy, P.; Hammes, P.S. Response of Potato (Solanum tuberosum) Tuber Yield Components to Gel-Polymer Soil Amendments and Irrigation Regimes. N. Z. J. Crop Hortic. Sci 2007, 35, 25–31. [Google Scholar] [CrossRef] [Green Version]
- Ahmadi, S.H.; Andersen, M.N.; Plauborg, F.; Poulsen, R.T.; Jensen, C.R.; Sepaskhah, A.R.; Hansen, S. Effects of Irrigation Strategies and Soils on Field Grown Potatoes: Yield and Water Productivity. Agric. Water Manag. 2010, 97, 1923–1930. [Google Scholar] [CrossRef]
- Li, W.; Xiong, B.; Wang, S.; Deng, X.; Yin, L.; Li, H. Regulation Effects of Water and Nitrogen on the Source-Sink Relationship in Potato during the Tuber Bulking Stage. PLoS ONE 2016, 11, e0146877. [Google Scholar] [CrossRef] [Green Version]
- Sadeghipour, O.; Aghaei, P. Response of Common Bean (Phaseolus vulgaris L.) to Exogenous Application of Salicylic Acid (SA) under Water Stress Conditions. Adv. Environ. Biol. 2012, 6, 1160–1168. [Google Scholar]
- Passioura, J.B. Drought and drought tolerance. In Drought Tolerance in Higher Plants: Genetical, Physiological and Molecular Biological Analysis; Springer: Dordrecht, The Netherlands, 1996; pp. 1–5. [Google Scholar]
- Sinclair, T.R. Is transpiration efficiency a viable plant trait in breeding for crop improvement? Funct. Plant Biol. 2012, 39, 359–365. [Google Scholar] [CrossRef] [Green Version]
- Manschadi, A.M.; Christopher, J.T.; de Voil, P.; Hammer, G.L. The role of root architectural traits in adaptation of wheat to waterlimited environments. Funct. Plant Biol. 2006, 33, 823–837. [Google Scholar] [CrossRef] [Green Version]
- Choudhary, S.; Mutava, R.N.; Shekoofa, A.; Sinclair, T.R.; Vara Prasad, P.V. Is the stay-green trait in sorghum a result of transpiration sensitivity to either soil drying or vapor pressure deficit? Crop Sci. 2013, 53, 2129–2134. [Google Scholar] [CrossRef]
- Gholipoor, M.; Sinclair, T.R.; Prasad, P.V. Genotypic variation within sorghum for transpiration response to drying soil. Plant Soil 2012, 357, 35–40. [Google Scholar] [CrossRef]
- Sadras, V.O.; Milroy, S.P. Soil-water thresholds for the response of leaf expansion and gas exchange. A review. Field Crops Res. 1996, 47, 253–266. [Google Scholar] [CrossRef]
- Devi, M.J.; Sinclair, T.R.; Vadez, V.; Krishnamurthy, L. Peanut genotypic variation in transpiration efficiency and decreased transpiration during progressive soil drying. Field Crops Res. 2009, 114, 280–285. [Google Scholar] [CrossRef] [Green Version]
- Ray, J.D.; Sinclair, T.R. Stomatal closure of maize hybrids in response to drying soil. Crop Sci. 1997, 37, 803–807. [Google Scholar] [CrossRef]
- Zhang, P.; Yang, X.; Manevski, K.; Li, S.H.; Wei, Z.H.; Andersen, M.N.; Liu, F. Physiological and Growth Responses of Potato (Solanum tuberosum L.) to Air Temperature and Relative Humidity under Soil Water Deficits. Plants 2022, 11, 1126. [Google Scholar] [CrossRef]
- Juenger, T.E.; Verslues, P.E. Time for a drought experiment: Do you know your plants’ water status? Plant Cell 2023, 35, 10–23. [Google Scholar] [CrossRef]
- Fu, Z.; Ciais, P.; Feldman, A.F.; Gentine, P.; Makowski, D.; Prentice, I.C.; Stoy, P.C.; Bastos, A.; Wigneron, J.-P. Critical soil moisture thresholds of plant water stress in terrestrial ecosystems. Sci. Adv. 2022, 8, eabq7827. [Google Scholar] [CrossRef]
- Massmann, A.; Gentine1, P.; Lin, C. When Does Vapor Pressure Deficit Drive or Reduce Evapotranspiration? J. Adv. Model. Earth Syst. 2019, 11, 3305–3320. [Google Scholar] [CrossRef] [Green Version]
- Grossiord, C.; Buckley, T.N.; Cernusak, L.A.; Novick5, K.A.; Poulter, B.; Siegwolf, R.T.W.; Sperry, J.S.; McDowell8, N.J. Plant responses to rising vapor pressure deficit. New Phytol. 2020, 226, 1550–1566. [Google Scholar] [CrossRef] [Green Version]
- Krishnamurthy, L.; Vadez, V.; Devi, M.J.; Serraj, R.; Nigam, S.N.; Sheshshayee, M.S.; Chandra, S.; Aruna, R. Variation in transpiration efficiency and its related traits in a groundnut (Arachis hypogaea L.) mapping population. Field Crops Res. 2007, 103, 189–197. [Google Scholar] [CrossRef] [Green Version]
- Islam, M.S. Identification and Evaluation of Drought-Adaptive Traits in Potato. Ph.D. Thesis, University of Natural Resources and Life Sciences, Vienna, Austria, 2017. [Google Scholar]
- Ray, J.D.; Sinclair, T.R. The effect of pot size on growth and transpiration of maize and soybean during water deficit stress. J. Exp. Bot. 1998, 49, 1381–1386. [Google Scholar] [CrossRef]
- Sinclair, T.R.; Ludlow, M.M. Influence of soil water supply on the plant water balance of four tropical grain legumes. Funct. Plant Biol. 1986, 13, 329–341. [Google Scholar] [CrossRef]
- Trevisan de Souza, A.; Streck, N.A.; Heldwein, A.B.; Bisognin, D.A.; Minussi Winck, J.E.; Schmitz Marques da Rocha, T.; Zanon, A.J. Transpiration and leaf growth of potato clones in response to soil water deficit. Sci. Agric. 2014, 71, 96–104. [Google Scholar] [CrossRef] [Green Version]
- Cirelli, D.; Lieffers, V.J.; Tyree, M.T. Measuring whole-plant transpiration gravimetrically: A scalable automated system built from components. Trees 2012, 26, 1669–1676. [Google Scholar] [CrossRef]
- Devi, M.J.; Sinclair, T.R. Diversity in drought traits among commercial southeastern US peanut cultivars. Int. J. Agron. 2011, 2011, 754658. [Google Scholar] [CrossRef]
- Ray, J.D.; Gesch, R.W.; Sinclair, T.R.; Hartwell Allen, L. The effect of vapor pressure deficit on maize transpiration response to a drying soil. Plant Soil 2002, 239, 113–121. [Google Scholar] [CrossRef]
- Zaman-Allah, M.; Jenkinson, D.M.; Vadez, V. Chickpea genotypes contrasting for seed yield under terminal drought stress in the field differ for traits related to the control of water use. Funct. Plant Biol. 2011, 38, 270–281. [Google Scholar] [CrossRef] [Green Version]
- Howell, T.A.; Dusek, D.A. Comparison of vapor-pressure-deficit calculation methods—Southern high plains. J. Irrig. Drain. Eng. 1995, 121, 191–198. [Google Scholar] [CrossRef]
- Ritchie, J.T. Water dynamics in the soil-plant-atmosphere system. In Soil Water and Nitrogen in Mediterranean-Type Environments; Plant and Soil; Springer: Hague, The Netherlands, 1981; pp. 81–96. [Google Scholar]
- Devi, M.J.; Redy, V. Transpiration Response of Cotton to Vapor Pressure Deficit and Its Relationship with Stomatal Traits. Front. Plant Sci. 2018, 9, 1572. [Google Scholar] [CrossRef] [Green Version]
- Devi, M.J.; Redy, V. Cotton Genotypic Variability for Transpiration Decrease with Progressive Soil Drying. Agronomy 2020, 10, 1290. [Google Scholar] [CrossRef]
- Purdom, S.; Shekoofa, A.; McClure, A.; Pantalone, V.; Arelli, P. Genotype identification for a water saving trait: Exploring early stomatal closure under soil drying among mid-South soybean. Agron. J. 2021, 114, 545–554. [Google Scholar] [CrossRef]
- Fletcher, A.L.; Sinclair, T.R.; Allen, H.L. Transpiration responses to vapor pressure deficit in well-watered ‘slow-wilting’ and commercial soybean. Environ. Exp. Bot. 2007, 6, 145–151. [Google Scholar] [CrossRef]
- Sinclair, T.R.; Hammer, G.L.; van Osterom, E.J. Potential yield and water-use efficiency benefits in sorghum from limited maximum transpiration rate. Funct. Plant Biol. 2005, 32, 945–952. [Google Scholar] [CrossRef]
- Vadez, V.; Choudhary, S.; Kholová, J.; Hash, C.T.; Srivastava, R.; Kumar, A.A.; Prandavada, A.; Anjaiah, M. Transpiration efficiency: Insights from comparisons of C4 cereal species. J. Exp. Bot. 2021, 72, 5221–5234. [Google Scholar] [CrossRef]
- Sinclair, T.R.; Bennett, J.M. Principles of Ecology in Plant Production; CAB International: Wallingford, UK, 1998; pp. 103–120. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotypes | Exp 1 7 Genotypes | Exp 2 7 Genotypes | Exp 3 4 Genotypes |
|---|---|---|---|
| Granola | Planting date: 15 December 2014 Harvest date: 12 February 2015 Plant age (d) at onset of drought stress: 25 Plant age (d) at the end: 59 Dry down days: 34 Avg. Temp (°C) at onset of drought stress: 16.9 Avg. Temp (°C) at the end: 16.8 Avg. RH (%) at onset of drought stress: 42.0 Avg. RH (%) at the end: 42.2 Avg. VPD (KPa) at onset of drought stress: 1.19 Avg. VPD (KPa) at the end: 1.16 Thermal time (GDD) at onset of drought stress: 253.4 Thermal time (GDD) at the end: 597.6 | ||
| Agria | |||
| Tosca | |||
| Diego | |||
| Cardinal | Planting date: 14 February 2015 Harvest date: 10 April 2015 Plant age (d) at onset of drought stress: 23 Plant age (d) at the end: 55 Dry down days: 32 Avg. Temp (°C) at onset of drought stress: 18.4 Avg. Temp (°C) at the end: 17.5 Avg. RH (%) at onset of drought stress: 49.9 Avg. RH (%) at the end: 43.8 Avg. VPD (KPa) at onset of drought stress: 1.35 Avg. VPD (KPa) at the end: 1.30 Thermal time (GDD) at onset of drought stress: 251.7 Thermal time (GDD) at the end: 585.6 | ||
| Diamant | |||
| Desiree | Planting date: 14 April 2016 Harvest date: 14 June 2016 Plant age (d) at onset of drought stress: 33 Plant age (d) at the end: 61 Dry down days: 28 Avg. Temp (°C) at onset of drought stress: 18.1 Avg. Temp (°C) at the end: 20.4 Avg. RH (%) at onset of drought stress: 37.4 Avg. RH (%) at the end: 51.3 Avg. VPD (KPa) at onset of drought stress: 1.52 Avg. VPD (KPa) at the end: 1.59 Thermal time (GDD) at onset of drought stress: 406.4 Thermal time (GDD) at the end: 878.1 | ||
| Caesar | |||
| Spunta | |||
| Farida | |||
| Mondial |
| Parameters | Formula | Explanation |
|---|---|---|
| Daily transpiration | Ti = Wi − Wi−1 | i = measurement day (i = 1, 2, 3, …, n), Ti = daily transpiration Wi = daily pot weight |
| Water requirements for WW plants | Wati = [(W0 − 240) − Wi] | W0 = initial day pot weight Wati = daily water requirement if Wi ≤ W0 − 240; otherwise, Wati = 0 |
| Water requirements for WS plants | Wati = Ti − 80 | if Ti ≥ 80; otherwise, Wati = 0 |
| Transpiration normalization (first step) | TRi = [Ti/Tw of (Rep1 + Rep2, …, + Rep5)/5] | TRi = daily transpiration ratio Ti = daily transpiration of WS plants Tw = daily transpiration of WW plants Rep1, …, Rep5 = replications |
| Transpiration normalization (second step) | NTRn = [TRn/(TR2 + TR3 + TR4 + TR5)/4] | NTRn = normalized transpiration rates of WS plants on day n TRn = transpiration rate of WS plants on day n |
| Transpiration efficiency | M = accumulated biomass (g), T = accumulated water transpired (l) | |
| VPD | VPD = e*(T) × (1 − ) e*(T) = 0.611 × exp | e*(T) = saturated vapour pressure in kPa at a given temperature T = temperature (°C) RH = relative humidity (%) |
| FTSW | FTSW = (Wi − Wend)/W0 − Wend) | Wi = daily pot weight Wend = final day pot weight W0 = initial day pot weight |
| FTSW threshold | Region 1 (t) = , t1 ≤ t ≤ T1 Region 2 (t) = , T1 ≤ t ≤ t2 | Region 1 = regression line starting from the maximum FTSW to the breakpoint Region 2 = regression line starting from the break point to minimum FTSW y = transpiration of Phase I to Phase II x = FTSW T1 = transition or the breakpoint t1 = min(t), the minimum FTSW = 0 t2 = max(t), the maximum FTSW = 1.0 |
| Experimental Condition | Genotype | Soil Water Consumed until FTSW Threshold (mL) | Water Saved Compared to Highest Consuming Genotype (%) | Water Saved Compared to Highest Consuming Genotype (mL) | FTSW Threshold of WS Plants | Relative Biomass Production (%) of Genotypes under Water Stress (WS) |
|---|---|---|---|---|---|---|
| Low VPD (Exp 1) | Granola | 1378.4 a | 0.0 | 0.0 | 0.19 | 36.9 b |
| Diamant | 1262.5 ab | 9.2 | 115.9 | 0.22 | 66.2 a | |
| Cardinal | 1241.8 ab | 11.0 | 136.6 | 0.20 | 48.3 ab | |
| Desiree | 1217.6 ab | 13.2 | 160.8 | 0.27 | 56.1 ab | |
| Agria | 1129.9 ab | 22.0 | 248.5 | 0.28 | 52.5 ab | |
| Diego | 1097.0 ab | 25.7 | 281.4 | 0.36 | 58.1 ab | |
| Tosca | 993.1 b | 38.8 | 385.3 | 0.36 | 47.7 ab | |
| Average | 1188.6 | 17.1 | 189.8 | 0.27 | 52.3 | |
| Moderate VPD (Exp 2) | Diamant | 928.4 a | 0.0 | 0.0 | 0.24 | 58.6 ab |
| Desiree | 766.8 b | 21.1 | 161.6 | 0.24 | 50.2 b | |
| Cardinal | 755.1 b | 22.9 | 173.2 | 0.28 | 62.4 ab | |
| Caesar | 749.7 b | 23.8 | 178.7 | 0.31 | 79.5 a | |
| Mondial | 702.9 b | 32.1 | 225.5 | 0.24 | 54.4 b | |
| Farida | 650.0 b | 42.8 | 278.4 | 0.29 | 52.7 b | |
| Spunta | 645.9 b | 43.7 | 282.4 | 0.31 | 52.3 b | |
| Average | 742.7 | 26.6 | 185.7 | 0.27 | 58.6 | |
| High VPD (Exp 3) | Desiree | 1298.0 a | 0.0 | 0.0 | 0.19 | 70.3 b |
| Caesar | 1290.5 a | 0.6 | 7.6 | 0.21 | 91.0 a | |
| Spunta | 1234.9 a | 5.1 | 63.2 | 0.25 | 43.9 c | |
| Farida | 1001.6 b | 29.6 | 296.4 | 0.29 | 28.1 c | |
| Average | 1206.3 | 8.8 | 91.8 | 0.24 | 58.3 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salehi-Soghadi, Z.; Islam, M.S.; Manschadi, A.M.; Kaul, H.-P. Transpiration Efficiency of Some Potato Genotypes under Drought. Agronomy 2023, 13, 996. https://doi.org/10.3390/agronomy13040996
Salehi-Soghadi Z, Islam MS, Manschadi AM, Kaul H-P. Transpiration Efficiency of Some Potato Genotypes under Drought. Agronomy. 2023; 13(4):996. https://doi.org/10.3390/agronomy13040996
Chicago/Turabian StyleSalehi-Soghadi, Zohreh, Md. Saiful Islam, Ahmad M. Manschadi, and Hans-Peter Kaul. 2023. "Transpiration Efficiency of Some Potato Genotypes under Drought" Agronomy 13, no. 4: 996. https://doi.org/10.3390/agronomy13040996
APA StyleSalehi-Soghadi, Z., Islam, M. S., Manschadi, A. M., & Kaul, H. -P. (2023). Transpiration Efficiency of Some Potato Genotypes under Drought. Agronomy, 13(4), 996. https://doi.org/10.3390/agronomy13040996