
Study on the Mechanism of Grafting to Improve the Tolerance of Pepper to Low Temperature



Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Grafting and Low Temperature Treatment
2.3. Comparison of Transcriptome Analysis of Self-Rooted and Grafted Seedlings
2.3.1. RNA Extraction, Library Preparation and Sequencing
2.3.2. Processing of Sequencing Data
2.3.3. Functional Annotation
2.3.4. Real-Time Fluorescence Quantitative PCR
2.4. Determination of Physiological Parameters
2.5. Data Processing
3. Results
3.1. Transcriptome Analysis of Pepper Seedlings at Low Temperature
3.1.1. Raw Data Collation, Filtering and Analysis
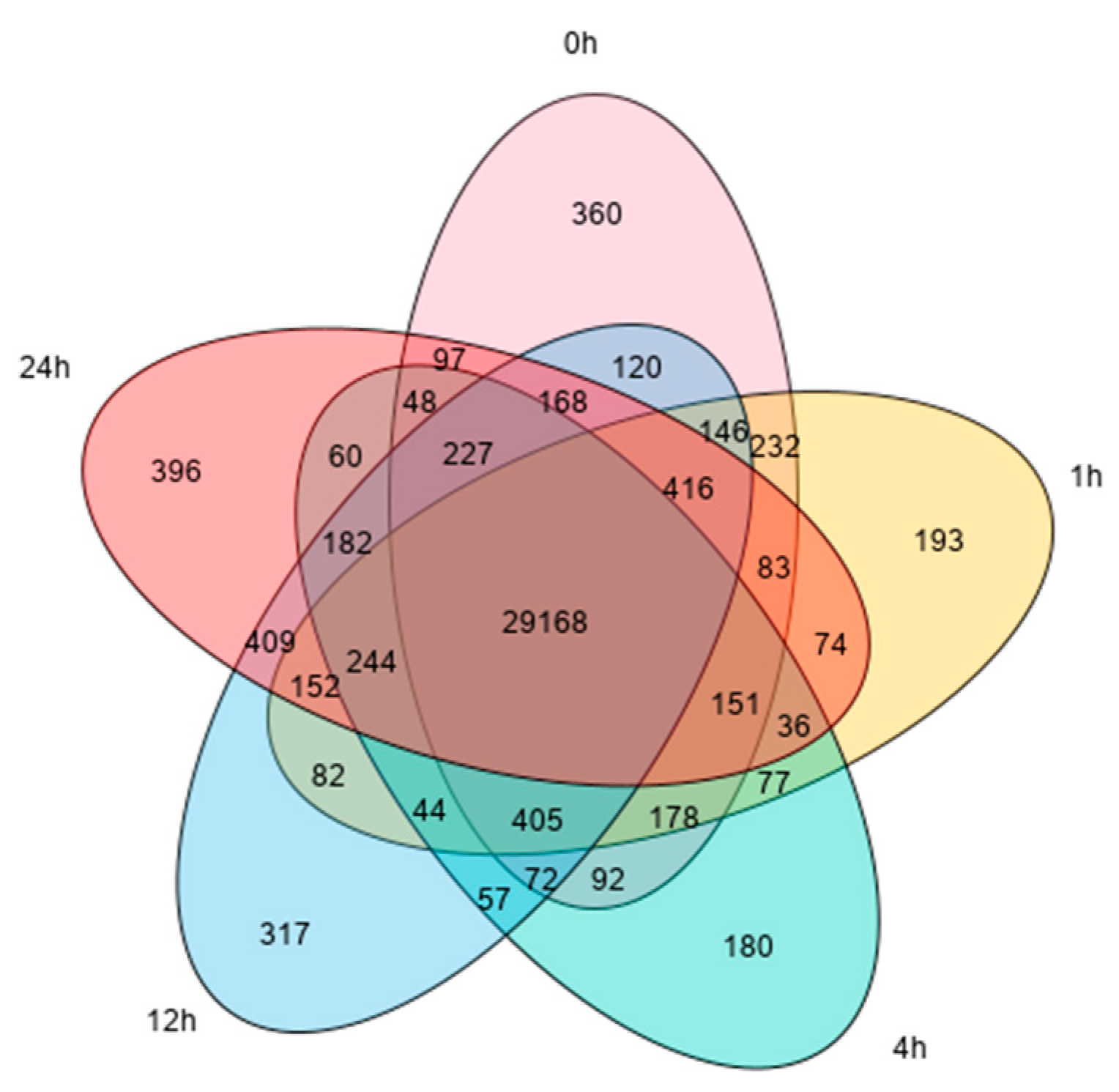
3.1.2. Analysis of the Differentially Expressed Genes
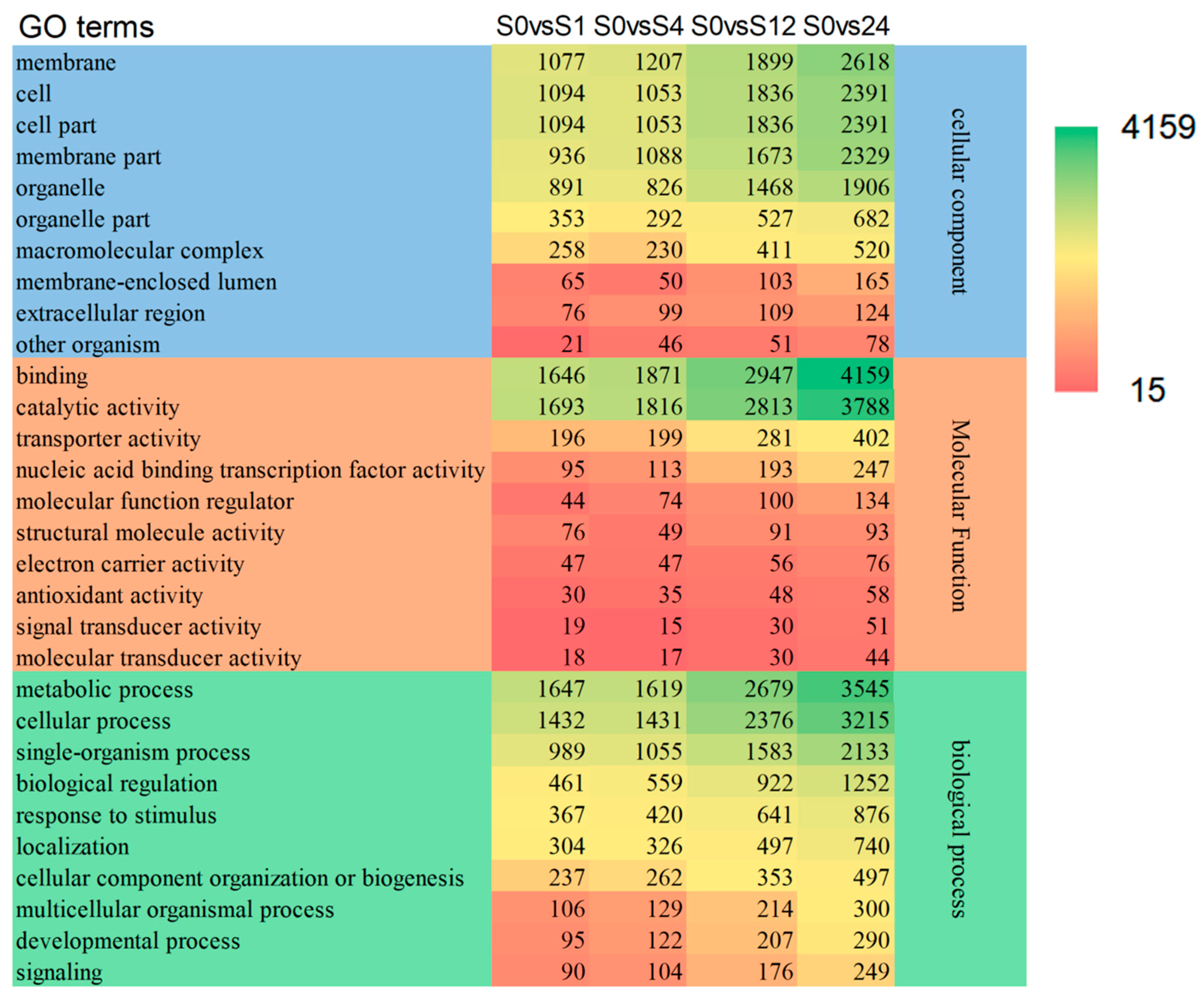
3.1.3. GO Analysis
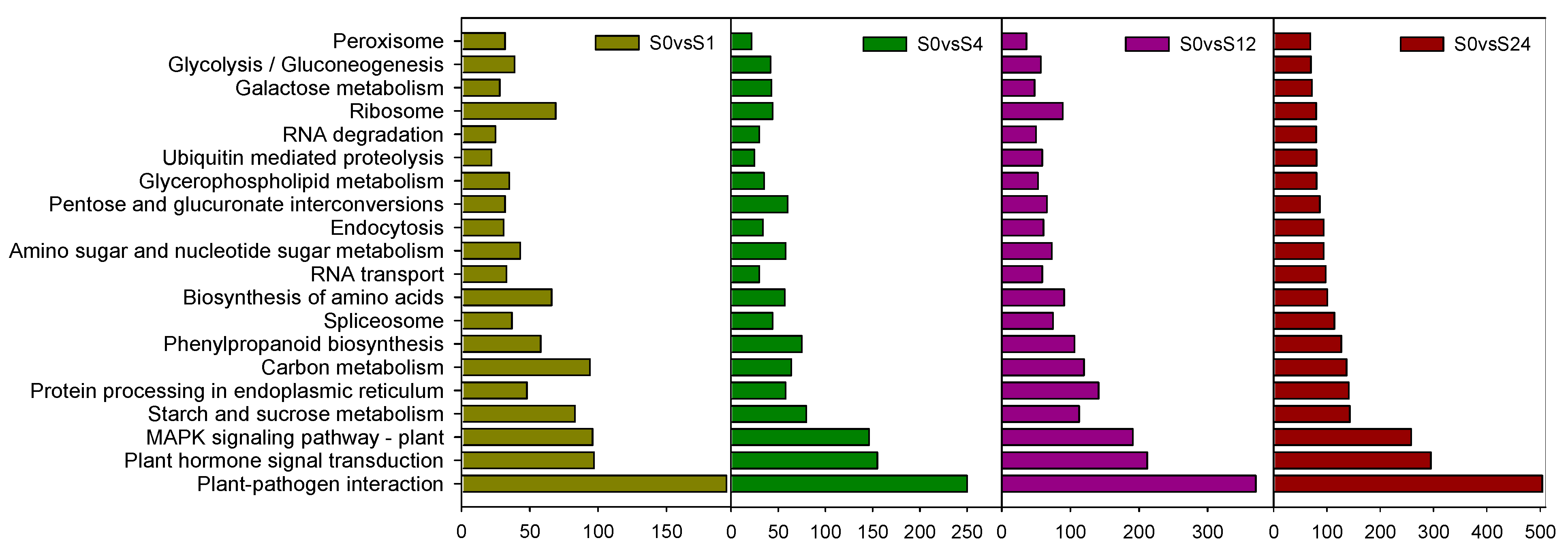
3.1.4. KEGG Analysis
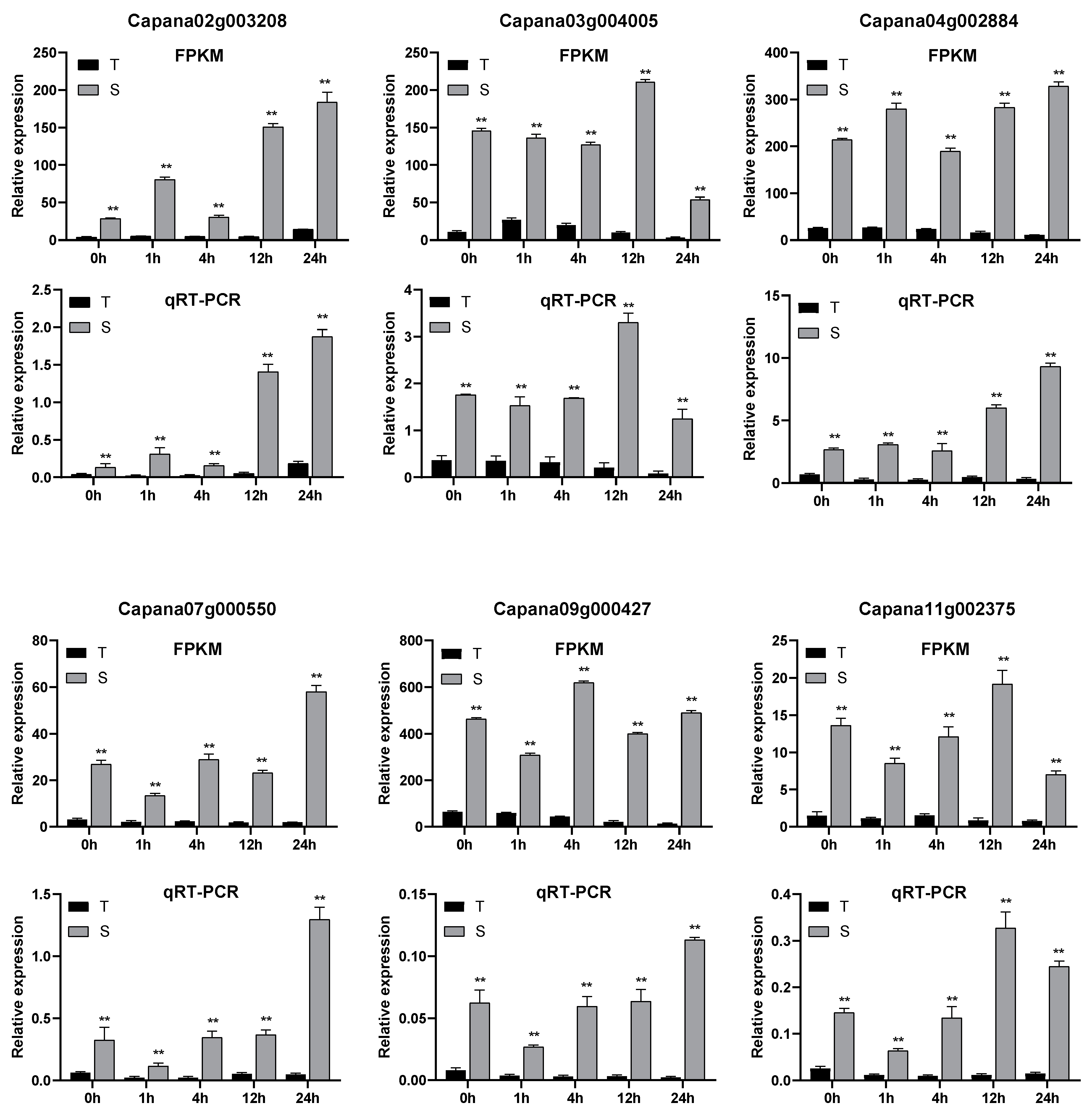
3.1.5. qRT-PCR Validation
3.2. Effect of Low Temperature on Pepper Morphology
3.3. Effect of Low Temperature on Physiological and Biochemical Indicators of Pepper
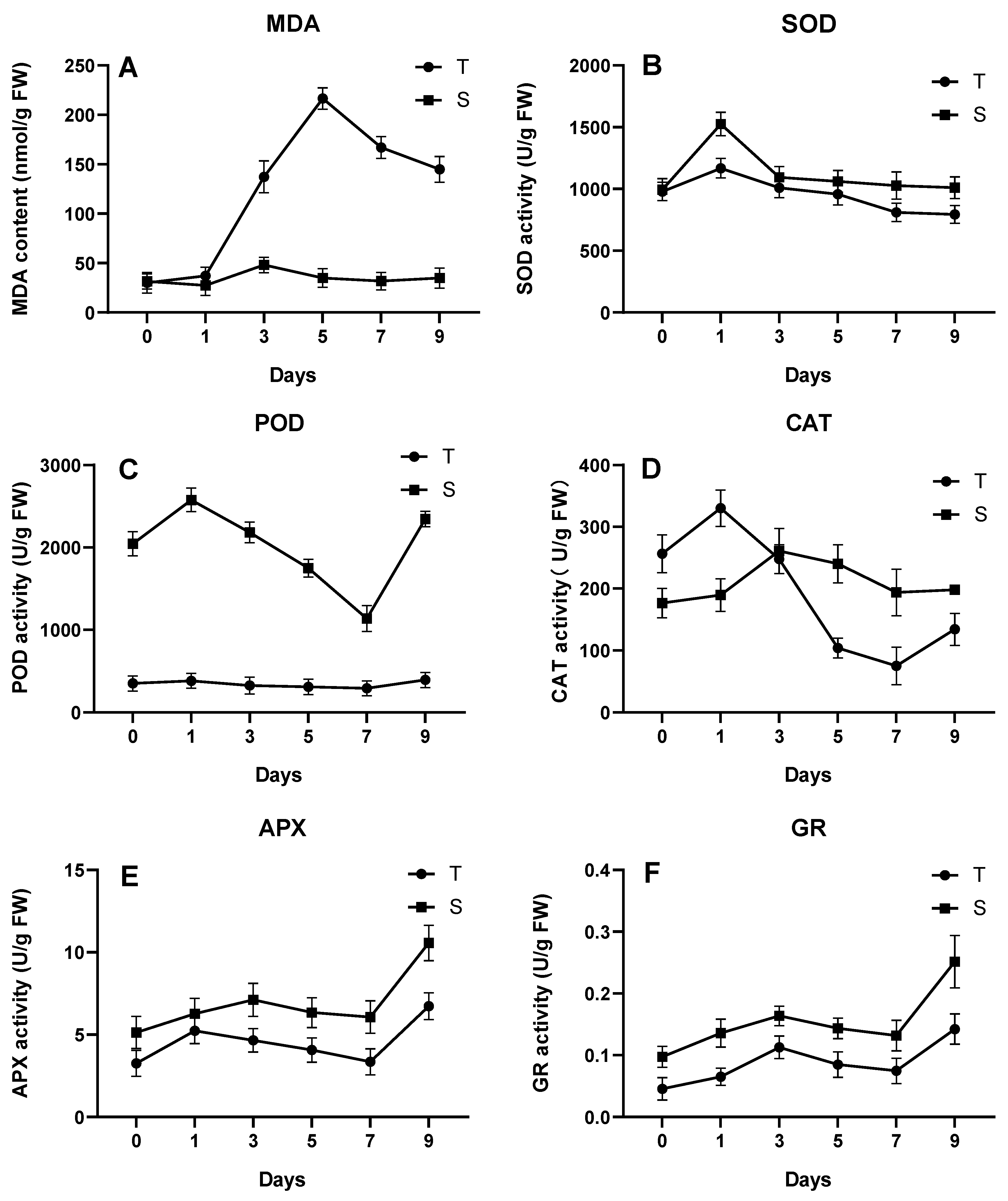
3.3.1. Effect of Low Temperature on Osmoregulation in Pepper
3.3.2. Effect of Low Temperature on Membrane Lipid Peroxidation in Pepper Seedlings
4. Discussion
4.1. Short-Term Response of Grafted Seedlings to Low Temperature
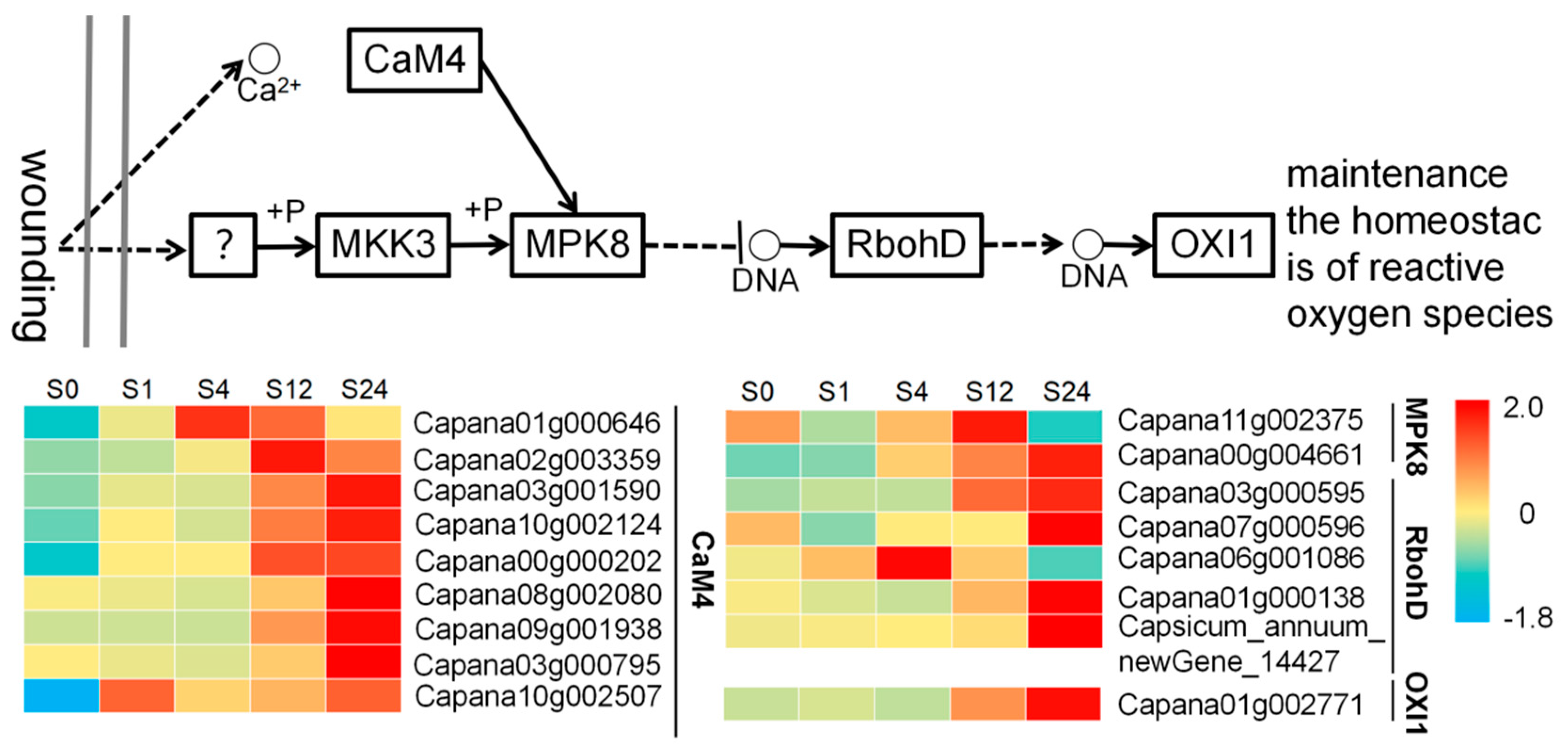
4.2. Mechanism for Improving Low Temperature Tolerance of Grafted Seedlings
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zou, X.X.; Ma, Y.Q.; Dai, X.Z.; Li, X.F.; Yang, S. Spread and industry development of pepper in China. Acta Hortic. Sin. 2020, 47, 1715–1726. [Google Scholar]
- Liu, Z.B.; Yang, B.Z.; Huang, R.Y.; Suo, H.; Zhang, Z.Q.; Chen, W.C.; Dai, X.Z.; Zou, X.X.; Ou, L.J. Transcriptome- and Proteome-wide association of Real Isogenic Line populations identified twelve core QTLs for four fruit traits in pepper (Capsicum annuum L.). Hortic. Res. 2022, 9, uhac015. [Google Scholar] [CrossRef] [PubMed]
- Chai, W.C.; Ma, R.L.; Jiao, Y.S.; Xing, G.M. Influences of low temperature stress on pepper’s growth index and physiological index. Acta Agric. Boreali Sin. 2010, 25, 168–171. [Google Scholar]
- Yang, A.M.; Shen, Z.Y. The Effects of low temperature acclimation on cold tolerance in cucumber seedlings. Acta Hortic. Sin. 1992, 1, 61–66. [Google Scholar]
- Liu, X.Y.; Wu, C.Y.; Zhang, G.C.; Yang, L.; Liang, C. Mechanisms of spraying external nitric oxide to alleviate injury of pepper seedlings caused by low temperature. J. Northwest A F Univ. Nat. Sci. Ed. 2020, 48, 63–70. [Google Scholar]
- Yang, W.J.; Jiang, X.M.; Gao, H.; Liu, S.Y.; Sun, T.Y.; Ma, C.J.; Bai, G.L.; Yu, X.H. Effects of 28-homobrassinolide on photosynthetic and fluorescence characteristics of pepper seedlings under low temperature with dim light. J. South. Agric. 2018, 49, 741–747. [Google Scholar]
- Li, Z.Z. Efficient cultivation technique of overwintering stubble in solar greenhouse. China Cucurbits Veg. 2019, 32, 60–61. [Google Scholar]
- Xu, Q.Q.; Zhang, Z.M.; Xu, Y.Q.; Li, F.H.; Zhang, J.Z.; Wang, J.; Li, S.L. Preliminary research on double-head seedling cultivation technology of tomato grafting and scionChina. Vegetables 2022, 404, 122–125. [Google Scholar]
- Zhang, Q.; Nie, L.C.; Zhao, W.S.; Fang, S.Y.; Zhao, J.T.; Guo, K.D. Screening of melon cold resistant rootstocks and evaluation on cold tolerance of grafted seedlings. China Veg. 2022, 401, 85–91. [Google Scholar]
- Fu, X.; Zhang, Y.Y.; Che, H.J.; Zhang, X.W.; Feng, Y.Q.; Lu, W.L.; Ai, X.Z. Salicylic acid is involved in grafting-induced chilling tolerance in cucumber seedlings. Plant Physiol. J. 2023, 58, 1–15. [Google Scholar]
- Li, Y.M.; Cai, X.X.; Zhu, L.; Li, S.L.; Ma, C.S.; Wang, Y.; Mu, R.X.; Sun, S.R. The roles of MiR160a and its target genes in grafted watermelon seedlings under chilling stress. Chin. J. Biochem. Mol. Biol. 2022, 38, 1703–1713. [Google Scholar]
- Xu, S.L.; Chen, Q.Y.; Chen, X.Q.; Gao, J.S.; Li, S.H. Effect of grafting on ‘Jiashi’ muskmelon yield and its resistance to melon fusarium wilt. Acta Hortic. Sin. 2005, 32, 521–523. [Google Scholar]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [PubMed]
- Latef, A.A.H.A.; He, C.X. Arbuscular mycorrhizal influence on growth, photosynthetic pigments, osmotic adjustment and oxidative stress in tomato plants subjected to low temperature stress. Acta Physiol. Plant. 2011, 33, 1217–1225. [Google Scholar] [CrossRef]
- Sun, J.P.; Zhang, F.S.; Pi, Z. Effect of Low Temperature Stress on the Antioxidant System of Sugar Beet. Chin. Agric. Sci. Bull. 2022, 38, 26–32. [Google Scholar]
- Chen, W.C.; Yang, B.Z.; Zhou, S.D.; Dai, X.Z. Effects of different inducers on cold resistance physiological characteristics of Capsicum annuum. J. Hunan Agric. Univ. Nat. Sci. 2011, 37, 396–399. [Google Scholar] [CrossRef]
- Nie, X.M.; Luan, H.; Feng, G.L. Effects of Silicon Nutrition and Grafting Rootstocks on Chilling Tolerance of Cucumber Seedlings. Acta Hortic. Sin. 2022, 49, 1795–1804. [Google Scholar]
- Airaki, M.; Leterrier, M.; Mateos, R.M.; Valderrama, R.; Chaki, M.; Barroso, J.B.; DelRio, L.A.; Palma, J.M.; Corpas, F.J. Metabolism of reactive oxygen species and reactive nitrogen species in pepper (Capsicum annuum L.) plants under low temperature stress. Plant Cell Environ. 2021, 35, 281–295. [Google Scholar] [CrossRef]
- Wu, H.; Qin, Y.; Lin, C.Y.; Tang, S.Y.; Yang, W.Y. The comparison test between cleft grafting and casing pipe grafting method on Capsicum annuum L. North. Hortic. 2010, 208, 40–42. [Google Scholar]
- Zhou, L.X.; Yang, M.D.; Zhao, Z.H. Physiological Response and Drought Resistance Evaluation of Drought Stress and Rehydration on Different Elaeis Guineensis Varieties. Molecular Plant Breeding:1-20[2023-01-29]. Available online: http://kns.cnki.net/kcms/detail/46.1068.S.20220825.1206.008.html (accessed on 12 December 2022).
- Wang, T.F.; Pei, Y.H.; Guo, X.M.; Li, J.; Song, X.Y. Identification of the cold tolerance of three maize varieties in the seedling stage. J. Nucl. Agric. Sci. 2017, 31, 803–808. [Google Scholar]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of proline for water stress studies. Plant Soil 1973, 39, 305–307. [Google Scholar] [CrossRef]
- Zhang, S.W.; Zong, Y.J.; Fang, C.Y.; Huang, S.H.; Li, J.; Xu, J.H.; Wang, Y.F.; Liu, C.H. Optimization of anthrone colorimetric method for rapid determination of soluble sugar in barley leaves Food. Res. Dev. 2020, 41, 196–200. [Google Scholar]
- Li, H.S. Principles and Techniques of Plant Physiological and Biochemical Experiments; Higher Education Press: Beijing, China, 2000; pp. 134–169. [Google Scholar]
- Zhu, G.L.; Zhong, W.H.; Zhang, A.Q. Physiological Experiments in Plants; Peking University Press: Beijing, China, 1990; pp. 37–40. [Google Scholar]
- Trevor, E.; Kraus, R.; Austin, F. Paclobutrazol protects wheat seedlings from heat and paraquat injury is detoxification of active oxygen involved Plant. Cell Physiol. 1994, 35, 45–52. [Google Scholar]
- Tan, Y.L.; Zhang, Y.Y.; Gao, D.D.; Chen, D.; Shen, W.Q.; Zhou, J.H.; Huang, C.P. Effects of chilling stress on ascorbate peroxidase activity, malondialdehyde and proline contents of dendrobium candidum. J. Zhejiang Univ. Agric. Life Sci. 2012, 38, 400–406. [Google Scholar]
- Halliwell, B.; Foyer, C.H. Properties and physiological function of a glutathione reductase purified from spinach leaves by affinity chromatography. Planta 1978, 139, 9–17. [Google Scholar] [CrossRef]
- Yang, S.; Wang, J.; Tang, Z. Calcium/calmodulin modulates salt responses by binding a novel interacting protein SAMS1 in peanut (Arachis hypogaea L.). Crop J. 2023, 11, 21–32. [Google Scholar] [CrossRef]
- Agarwal, P.K.; Gupta, K.; Jha, B. Molecular characterization of the Salicornia brachiata SbMAPKK gene and its expression by abiotic stress. Mol. Biol. Rep. 2010, 37, 981–986. [Google Scholar] [CrossRef]
- Gu, L.K.; Liu, Y.K.; Zong, X.J.; Liu, L.X.; Li, D.P.; Li, D.Q. Overexpression of maize mitogen-activated protein kinase gene, ZmSIMK1 in Arabidopsis increases tolerance to salt stres. Mol. Biol. Rep. 2010, 37, 4067–4073. [Google Scholar] [CrossRef]
- Yu, Y.; Chen, H.Y.; Cheng, L.L. Cloning and expression analysis of MAPK gene under saline- alkaline stress in flax. J. Northeast. Agric. Univ. 2015, 46, 21–28. [Google Scholar]
- Li, Q.Y.; Li, P.; Htwe, S.P.M.N. Antepenultimate residue at the C-terminus of NADPH oxidase RBOHD is critical for its function in the production of reactive oxygen species in Arabidopsis. J. Zhejiang Univ. Sci. B Biomed. Biotechnol. 2019, 20, 783. [Google Scholar] [CrossRef]
- Zhang, T.G.; Lai, J.; Li, P. Expression analysis of RbohA and RbohD genes in Brassica campestris under different treatments. Chin. J. Ecol. 2019, 38, 173–180. [Google Scholar]
- Takahashi, F.; Mizoguchi, T.; Yoshida, R.; Ichimura, K.; Shinozaki, K. Calmodulin-Dependent Activation of MAP Kinase for ROS Homeostasis in Arabidopsis. Mol. Cell 2011, 41, 649–660. [Google Scholar] [CrossRef]
- Huang, X.C.; Chen, S.D.; Li, W.P.; Tang, L.L. ROS regulated reversible protein phase separation synchronizes plant flowering. Nat. Chem. Biol. 2021, 17, 549–557. [Google Scholar] [CrossRef]
- Leon, J.; Rojo, E.; Sanchez-Serrano, J.J. Wound signalling in plants. J. Exp. Bot. 2021, 52, 1–9. [Google Scholar] [CrossRef]
- Liao, H.; Zhu, M.; Cui, H. MARIS plays important roles in Arabidopsis pollen tube and root hair growth. J. Integr. Plant Biol. 2016, 58, 927–940. [Google Scholar] [CrossRef]
- Xiao, G.Z.; Wu, X.L.; Teng, K.; Chao, Y.H.; Li, W.T.; Han, L.B. Hyperspectral analysis and electrolyte leakage inversion of creeping bentgrass under salt stress. Spectrosc. Spectr. Anal. 2016, 36, 3630–3636. [Google Scholar]
- Li, W.M.; Xin, J.P.; Wei, C.Y.; Tian, R.N. Research progress on cold resistance in plants. Jiangsu Agric. Sci. 2017, 45, 6–11. [Google Scholar]
- Mahajan, S.; Tuteja, N. Cold, salinity and drought stresses: An overview. Arch. Biochem. Biophys. 2005, 444, 139–158. [Google Scholar] [CrossRef]
- Wang, R.; Ren, Y.; Yue, W. Effect of low temperature stress on the survival and physiological and biochemical indexes of alfalfa seedlings. Acta Laser Biol. Sin. 2006, 15, 342. [Google Scholar]
- Zou, K.X.; Shang, S.; Tian, L.B.; Zhu, G.P.; Zhou, M.M.; Pan, Q.Y.; Zeng, L.P. Effects of low temperature stress on osmotic solutes of grafted bitter gourd seedlings. Chin. J. Trop. Crops 2018, 39, 1533–1539. [Google Scholar]
- Levitt, J. Responses of Plants to Environmental Stresses; Academic Press: New York, NY, USA, 1972. [Google Scholar]
- Zhang, S.P.; Gu, X.F.; Wang, Y.; Zhang, S.Q. Effect of low temperature stress on the physiological and biochemical indexes in cucumber seedling grafted on wild cucumber(sicyos angulatus). Acta Bot. Boreali Occident. Sin. 2005, 25, 1428–1432. [Google Scholar]
- Crowe, J.H. Advances in Experimental Medicine and Biology; IwayaInoue, M., Sakurai, M., Uemura, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2018; Volume 1081, pp. 5–6. [Google Scholar]
- Gallego, S.M.; Benavides, M.P.; Tomaro, M.L. Effect of heavy metal ion excess on sunflower leaves: Evidence for involvement of oxidative stress. Plant Sci. 1996, 121, 151–159. [Google Scholar] [CrossRef]
- Wang, J.N.; Shang, S.; Tian, L.B.; Zhou, M.M.; Pan, Q.Y.; Zou, K.X.; Du, Z.T. Effects of low temperature stress on antioxidant system of grafted bitter gourd seedlings. Chin. J. Trop. Crops 2018, 39, 237–245. [Google Scholar]
- Lu, X.G.; Zhou, M.D.; Li, M. Effects of low temperature stress on growth and physiological characteristics of grafted and own root melon seedlings. China Veg. 2016, 53–57. [Google Scholar]
- Gao, J.J.; Qin, A.G.; Yu, C.X. Effects of grafting on cucumber leaf SOD and CAT gene expression and activities under low temperature stress. Chin. J. Appl. Ecol. 2009, 20, 213–217. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Forward Primer | Reverse Primer |
|---|---|---|
| Capana02g003208 | GGTATGCGCTTGCCTCCCAT | TGGACCTCCTCGCCGAAAGA |
| Capana03g004005 | CGATGCTCCAAACAAGCGCA | GCCCAGCCCAAGTTACCCAG |
| Capana04g002884 | GTGGCTGCCAATGACGTTGC | GGAGTGGCTGGGGAGAGCTA |
| Capana07g000550 | AACAGGACATTGGCTGGGGC | TGCGACTCCCTTTGATGGCG |
| Capana09g000427 | TTCCGCGAGTCTCTCTGGCT | ATGACACCTCCTGCCCCCAT |
| Capana11g002375 | CCAAGGCGACCTGCCTCTTT | GTCGTCCCAAACTGCCCCAA |
| β-Actin | CCACCTCTTCACTCTCTGCTCT | ACTAGGAAAAACAGCCCTTGGT |
| Up-Regulated | Down Regulated | Total DEGs | |
|---|---|---|---|
| S0 vs. S1 | 2162 | 2623 | 4785 |
| S0 vs. S4 | 2572 | 2924 | 5496 |
| S0 vs. S12 | 4437 | 4535 | 8972 |
| S0 vs. S24 | 6282 | 5590 | 11,872 |
| Total | 15,453 | 15,672 | 31,125 |
| Treating Days | T 1 | S 2 |
|---|---|---|
| 0 | 24.97% ± 0.010 gh | 19.01% ± 0.015 i |
| 1 | 59.76% ± 0.031 d | 23.01% ± 0.020 h |
| 3 | 66.96% ± 0.006 bc | 38.89% ± 0.010 e |
| 5 | 79.23% ± 0.040 a | 34.83% ± 0.015 f |
| 7 | 69.18% ± 0.015 b | 27.63% ± 0.006 g |
| 9 | 65.81% ± 0.015 c | 38.60% ± 0.012 e |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Long, H.; Li, Z.; Suo, H.; Ou, L.; Miao, W.; Deng, W. Study on the Mechanism of Grafting to Improve the Tolerance of Pepper to Low Temperature. Agronomy 2023, 13, 1347. https://doi.org/10.3390/agronomy13051347
Long H, Li Z, Suo H, Ou L, Miao W, Deng W. Study on the Mechanism of Grafting to Improve the Tolerance of Pepper to Low Temperature. Agronomy. 2023; 13(5):1347. https://doi.org/10.3390/agronomy13051347
Chicago/Turabian StyleLong, Huijun, Ziyu Li, Huan Suo, Lijun Ou, Wu Miao, and Wenqiao Deng. 2023. "Study on the Mechanism of Grafting to Improve the Tolerance of Pepper to Low Temperature" Agronomy 13, no. 5: 1347. https://doi.org/10.3390/agronomy13051347
APA StyleLong, H., Li, Z., Suo, H., Ou, L., Miao, W., & Deng, W. (2023). Study on the Mechanism of Grafting to Improve the Tolerance of Pepper to Low Temperature. Agronomy, 13(5), 1347. https://doi.org/10.3390/agronomy13051347