

Changes in Soil Rhizobia Diversity and Their Effects on the Symbiotic Efficiency of Soybean Intercropped with Maize

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Plant Sampling and Analysis
2.3. Analysis of Soil Physicochemical Characteristics
2.4. Measurement of Soybean Nodules and Nodules Dry Weight
2.5. Determination of Nitrogen-Fixing Bacteria Diversity
2.5.1. DNA Extraction and PCR Amplification
2.5.2. Illumina MiSeq Sequencing
2.5.3. Data Processing
2.6. Determination of nifH Gene
2.7. Method of Calculating Soybean Nitrogen Fixation Efficiency
2.8. Statistical Analysis
3. Results
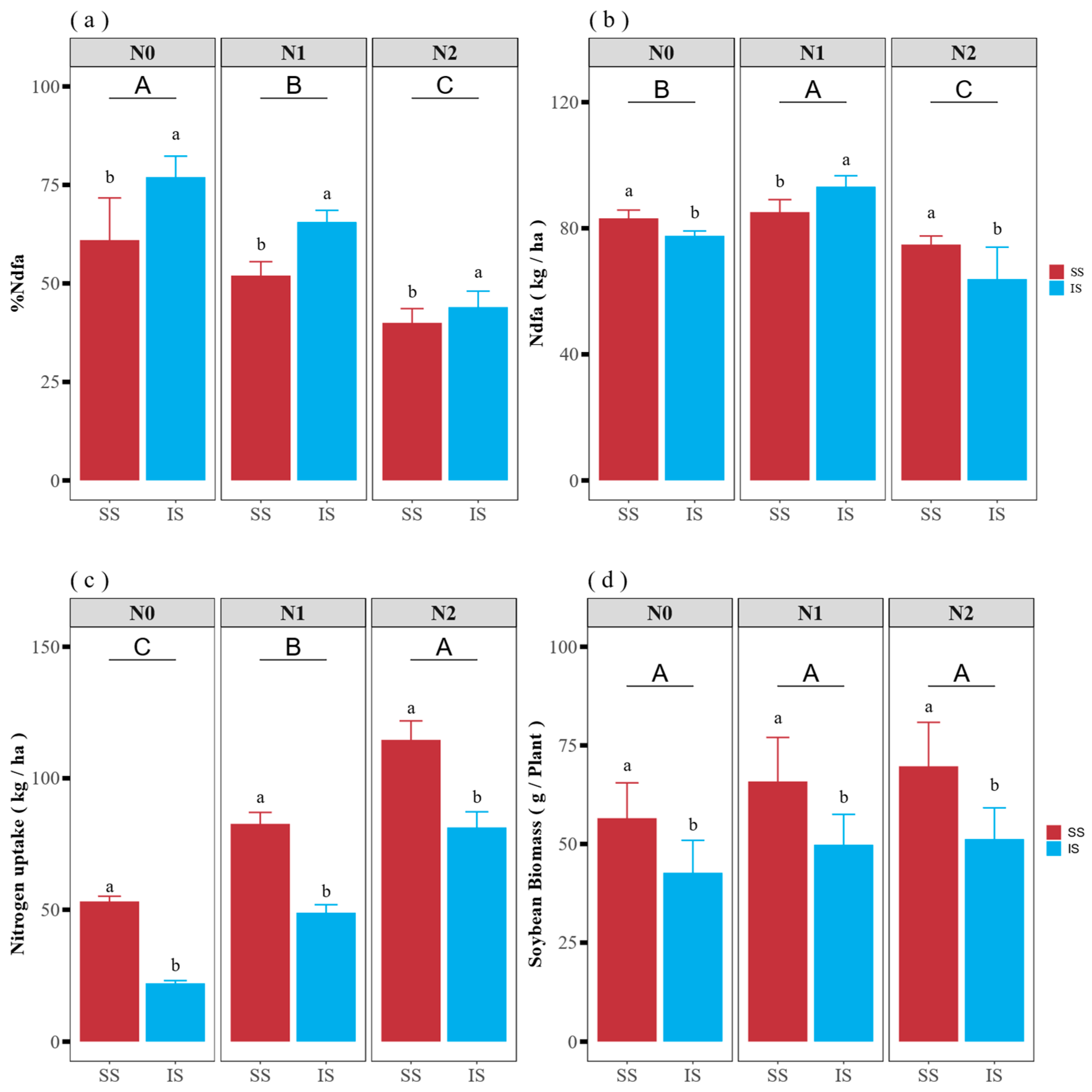
3.1. Soybean Nitrogen Fixation Efficiency, Biological N Fixation and N Uptake from the Soil
3.2. Soybean Nodules Numbers and Nodules Dry Weight
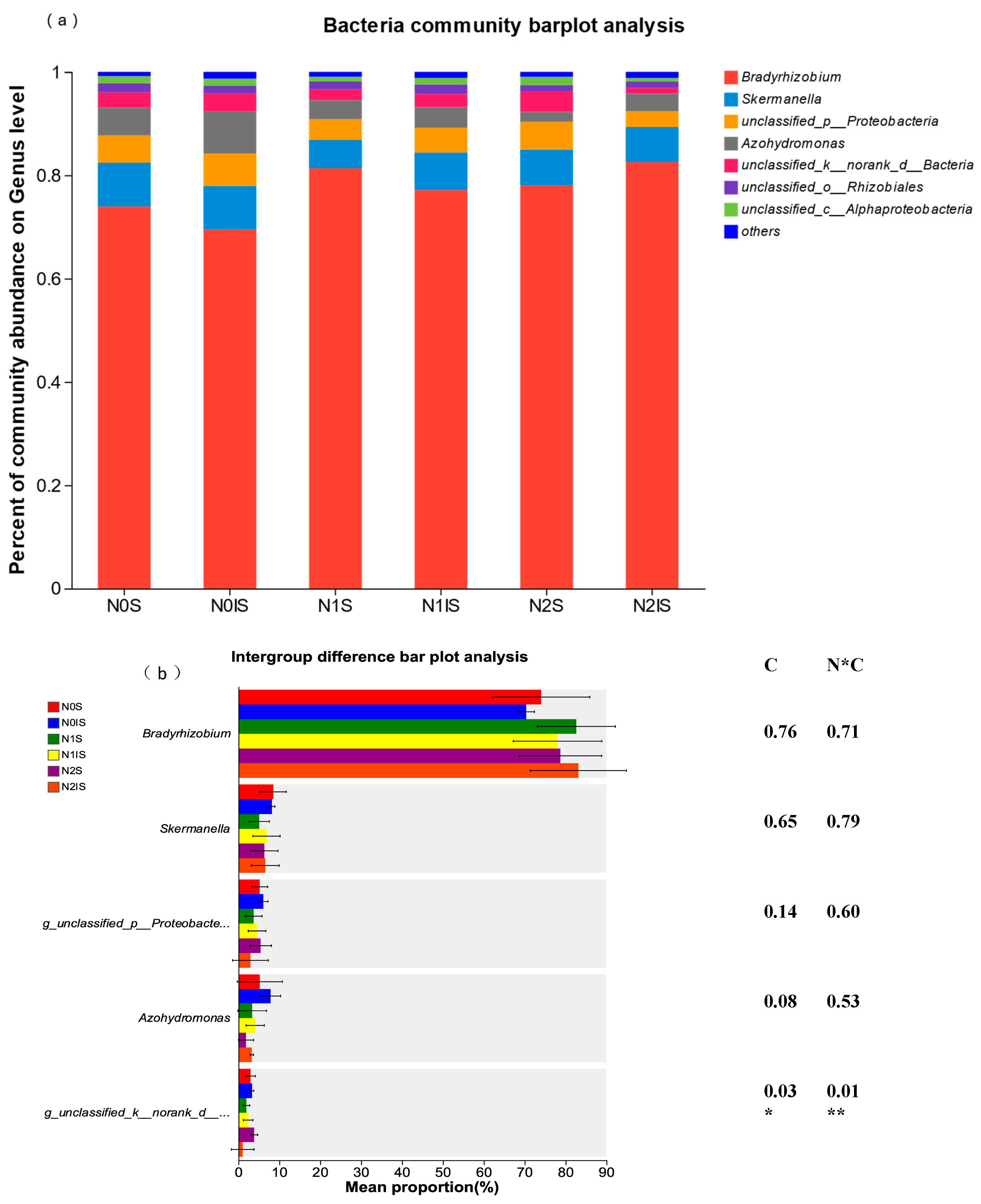
3.3. Alpha Diversity and Composition of Nitrogen-Fixing Bacteria of Soybean Rhizosphere Soil
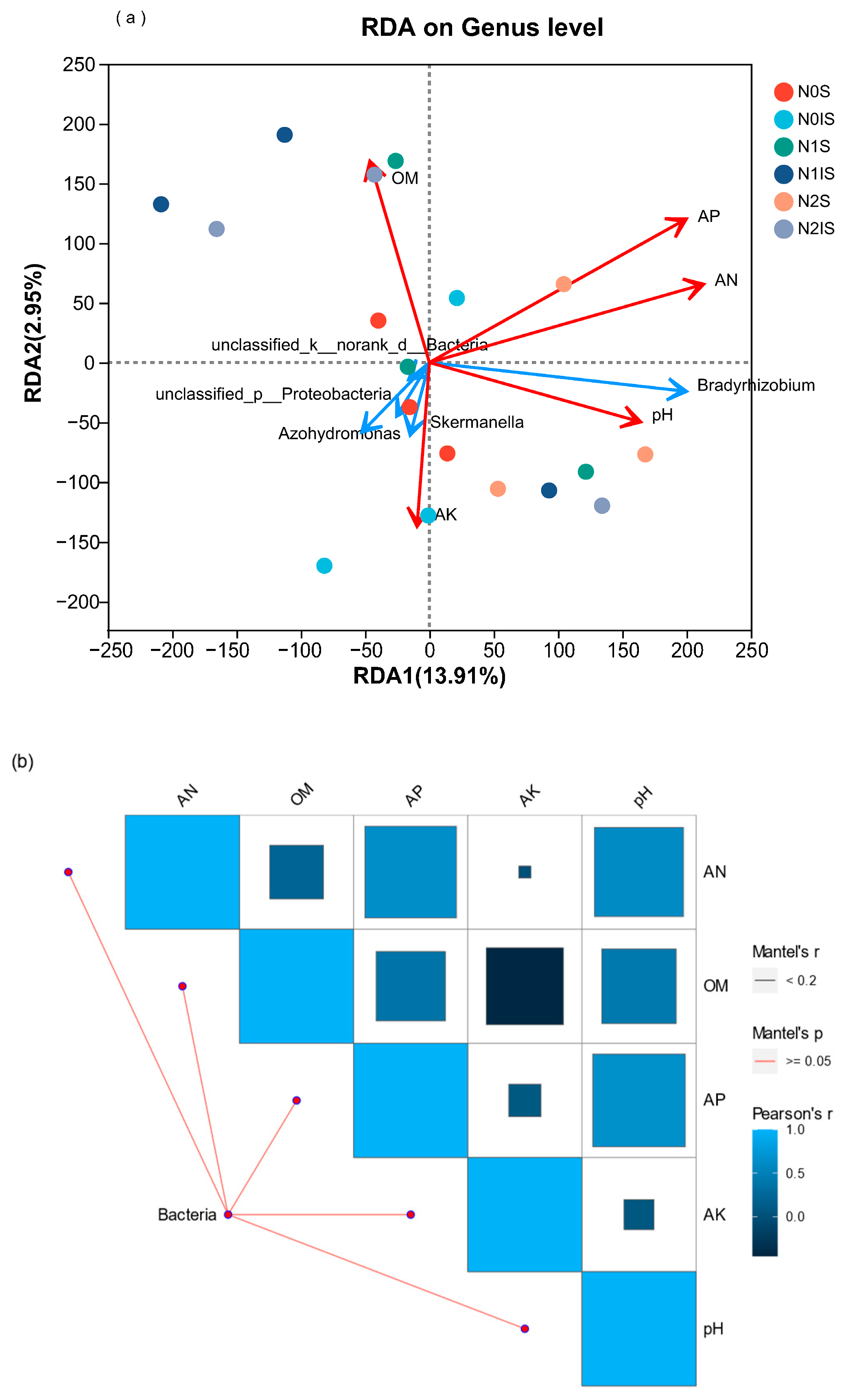
3.4. Relationship between the Nitrogen-Fixing Bacteria Community and Soil Physicochemical Characteristics
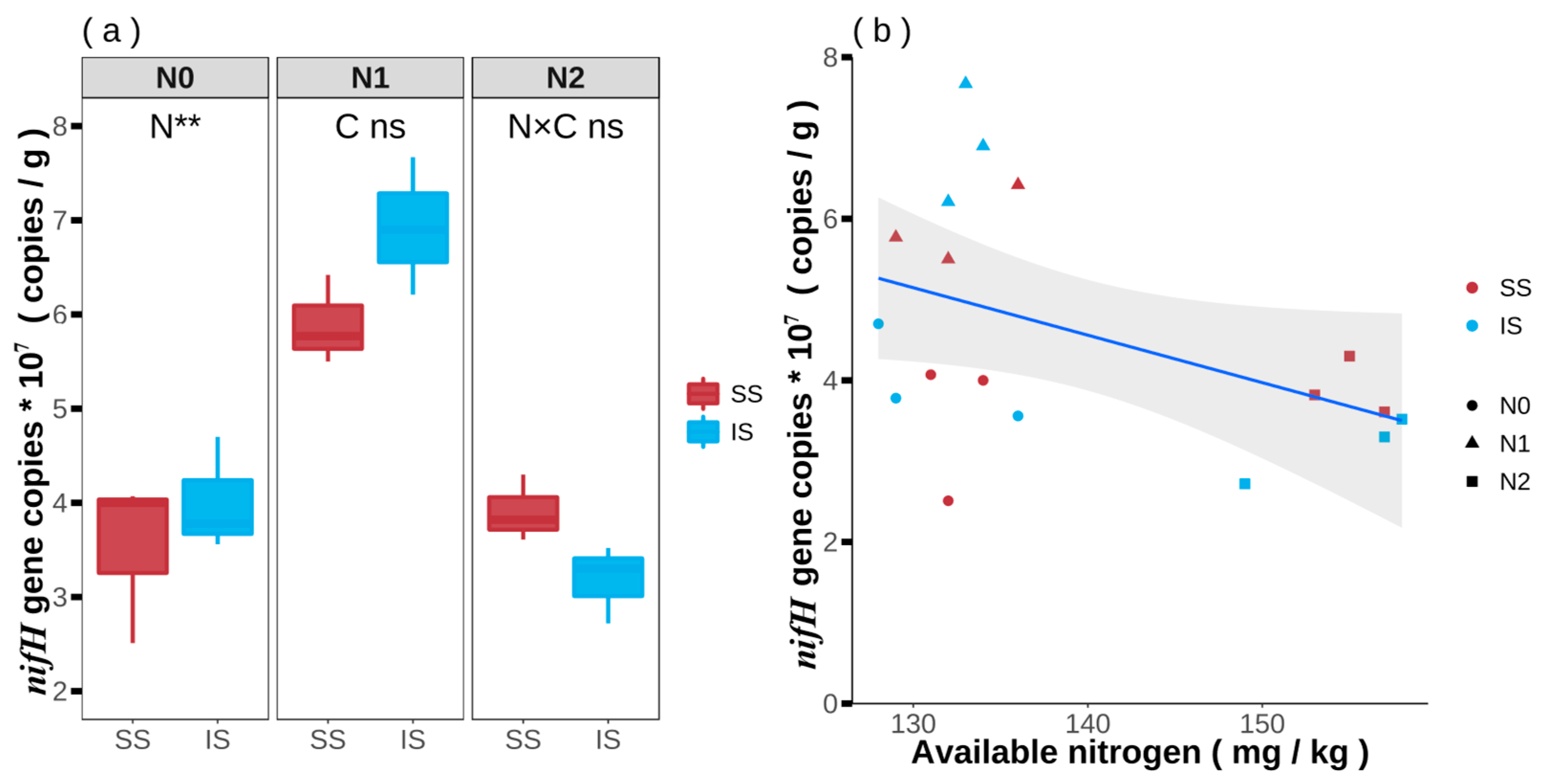
3.5. nifH Gene Copies of Nitrogen-Fixing Bacteria in Soybean and Maize Rhizosphere Soil
3.6. Correlation nifH Gene Copies and Soil Available and Biological Quantification of Nitrogen Fixation
4. Discussion
4.1. Nitrogen Fixation of Intercropped Soybean Affected by Nitrogen Application Rates
4.2. Diversity of Nitrogen-Fixing Bacteria in the Rhizosphere Soil of Intercropped Soybean Affected by N Application
4.3. NifH Gene Copies in the Rhizosphere Soil of Intercropped Soybean
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- da Silva, J.N.; Neto, F.B.; de Lima, J.S.S.; dos Santos, E.C.; Nunes, R.L.C.; Chaves, A.P. Production and benefits in carrot and vegetable cowpea associations under green manuring and spatial arrangements. Rev. Cienc. Agron. 2020, 51, e20197067. [Google Scholar] [CrossRef]
- Anas, M.; Liao, F.; Verma, K.K.; Sarwar, M.A.; Mahmood, A.; Chen, Z.L.; Li, Q.; Zeng, X.P.; Liu, Y.; Li, Y.R. Fate of nitrogen in agriculture and environment: Agronomic, eco-physiological and molecular approaches to improve nitrogen use efficiency. Biol. Res. 2020, 53, 47. [Google Scholar] [CrossRef] [PubMed]
- Nelson, K.A.; Massey, R.E.; Burdick, B.A. Harvest Aid Application Timing Affects Wheat and Relay Intercropped Soybean Yield. Agron. J. 2011, 103, 851–855. [Google Scholar] [CrossRef]
- Saudy, H.S. Maize–cowpea intercropping as an ecological approach for nitrogen-use rationalization and weed suppression. Arch. Agron. Soil Sci. 2014, 61, 1–14. [Google Scholar] [CrossRef]
- Coelho, M.R.R.; Marriel, I.E.; Jenkins, S.N.; Lanyon, C.V.; Seldin, L.; O’Donnell, A.G. Molecular detection and quantification of nifH gene sequences in the rhizosphere of sorghum (Sorghum bicolor) sown with two levels of nitrogen fertilizer. Appl. Soil Ecol. 2009, 42, 48–53. [Google Scholar] [CrossRef] [Green Version]
- Fu, Z.-d.; Zhou, L.; Chen, P.; Du, Q.; Pang, T.; Song, C.; Wang, X.-c.; Liu, W.-g.; Yang, W.-y.; Yong, T.-w. Effects of maize-soybean relay intercropping on crop nutrient uptake and soil bacterial community. J. Integr. Agric. 2019, 18, 2006–2018. [Google Scholar] [CrossRef]
- Chen, P.; Du, Q.; Liu, X.; Zhou, L.; Hussain, S.; Lei, L.; Song, C.; Wang, X.; Liu, W.; Yang, F.; et al. Effects of reduced nitrogen inputs on crop yield and nitrogen use efficiency in a long-term maize-soybean relay strip intercropping system. PLoS ONE 2017, 12, e0184503. [Google Scholar] [CrossRef] [Green Version]
- Shao, Z.; Wang, X.; Gao, Q.; Zhang, H.; Yu, H.; Wang, Y.; Zhang, J.; Nasar, J.; Gao, Y. Root Contact between Maize and Alfalfa Facilitates Nitrogen Transfer and Uptake Using Techniques of Foliar 15N-Labeling. Agronomy 2020, 10, 360. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Zeng, F.; Zou, Z.; Zhang, Z.; Li, Y. Nitrogen uptake and transfer in a soybean/maize intercropping system in the karst region of southwest China. Ecol. Evol. 2017, 7, 8419–8426. [Google Scholar] [CrossRef]
- Li, L.; Zhang, F.; Li, X.; Christie, P.; Sun, J.; Yang, S.; Tang, C. Interspecific facilitation of nutrient uptake by intercropped maize and faba bean. Nutr. Cycl. Agroecosystems 2003, 65, 61–71. [Google Scholar] [CrossRef]
- Xiao, Y.; Li, L.; Zhang, F. The interspecific nitrogen facilitation and the subsequent nitrogen transfer between the intercropped wheat and fababean. Sci. Agric. Sin. 2005, 38, 965–973. [Google Scholar]
- Li, S.; van der Werf, W.; Zhu, J.; Guo, Y.; Li, B.; Ma, Y.; Evers, J.B. Estimating the contribution of plant traits to light partitioning in simultaneous maize/soybean intercropping. J. Exp. Bot. 2021, 72, 3630–3646. [Google Scholar] [CrossRef]
- Fan, F.; Zhang, F.; Song, Y.; Sun, J.; Bao, X.; Guo, T.; Li, L. Nitrogen Fixation of Faba Bean (Vicia faba L.) Interacting with a Non-legume in Two Contrasting Intercropping Systems. Plant Soil 2006, 283, 275–286. [Google Scholar] [CrossRef]
- Homulle, Z.; George, T.S.; Karley, A.J. Root traits with team benefits: Understanding belowground interactions in intercropping systems. Plant Soil 2021, 471, 1–26. [Google Scholar] [CrossRef]
- Zhang, W.P.; Liu, G.C.; Sun, J.H.; Fornara, D.; Zhang, L.Z.; Zhang, F.F.; Li, L.; Niels, A. Temporal dynamics of nutrient uptake by neighbouring plant species: Evidence from intercropping. Funct. Ecol. 2016, 31, 469–479. [Google Scholar] [CrossRef]
- Solanki, M.K.; Wang, Z.; Wang, F.Y.; Li, C.N.; Gupta, C.L.; Singh, R.K.; Malviya, M.K.; Singh, P.; Yang, L.T.; Li, Y.R. Assessment of Diazotrophic Proteobacteria in Sugarcane Rhizosphere When Intercropped With Legumes (Peanut and Soybean) in the Field. Front. Microbiol. 2020, 11, 1814. [Google Scholar] [CrossRef] [PubMed]
- Malviya, M.K.; Solanki, M.K.; Li, C.-N.; Wang, Z.; Zeng, Y.; Verma, K.K.; Singh, R.K.; Singh, P.; Huang, H.-R.; Yang, L.-T.; et al. Sugarcane-Legume Intercropping Can Enrich the Soil Microbiome and Plant Growth. Front. Sustain. Food Syst. 2021, 5, 606595. [Google Scholar] [CrossRef]
- Lai, H.; Gao, F.; Su, H.; Zheng, P.; Li, Y.; Yao, H. Nitrogen Distribution and Soil Microbial Community Characteristics in a Legume–Cereal Intercropping System: A Review. Agronomy 2022, 12, 1900. [Google Scholar] [CrossRef]
- Hayden, H.L.; Drake, J.; Imhof, M.; Oxley, A.P.A.; Norng, S.; Mele, P.M. The abundance of nitrogen cycle genes amoA and nifH depends on land-uses and soil types in South-Eastern Australia. Soil Biol. Biochem. 2010, 42, 1774–1783. [Google Scholar] [CrossRef]
- Nicol, G.W.; Leininger, S.; Schleper, C.; Prosser, J.I. The influence of soil pH on the diversity, abundance and transcriptional activity of ammonia oxidizing archaea and bacteria. Environ. Microbiol. 2008, 10, 2966–2978. [Google Scholar] [CrossRef]
- Kasa, P.; Modugapalem, H.; Battini, K. Isolation, screening, and molecular characterization of plant growth promoting rhizobacteria isolates of Azotobacter and Trichoderma and their beneficial activities. J. Nat. Sci. Biol. Med. 2015, 6, 360–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, K.; Delgado-Baquerizo, M.; Guo, X.; Wang, D.; Wu, Y.; Zhu, M.; Yu, W.; Yao, H.; Zhu, Y.G.; Chu, H. Suppressed N fixation and diazotrophs after four decades of fertilization. Microbiome 2019, 7, 143. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Mu, Y.; Li, X.; Li, S.; Sang, P.; Wang, X.; Wu, H.; Xu, N. Response of the arbuscular mycorrhizal fungi diversity and community in maize and soybean rhizosphere soil and roots to intercropping systems with different nitrogen application rates. Sci. Total Environ. 2020, 740, 139810. [Google Scholar] [CrossRef] [PubMed]
- Simonsen, A.K.; Han, S.; Rekret, P.; Rentschler, C.S.; Heath, K.D.; Stinchcombe, J.R. Short-term fertilizer application alters phenotypic traits of symbiotic nitrogen fixing bacteria. PeerJ 2015, 3, e1291. [Google Scholar] [CrossRef] [Green Version]
- Hui, T. Effects of long-term fertilization on nifH gene diversity in agricultural black soil. Afr. J. Microbiol. Res. 2012, 6, 2659–2666. [Google Scholar] [CrossRef]
- Calderoli, P.A.; Collavino, M.M.; Behrends Kraemer, F.; Morras, H.J.M.; Aguilar, O.M. Analysis of nifH-RNA reveals phylotypes related to Geobacter and Cyanobacteria as important functional components of the N(2) -fixing community depending on depth and agricultural use of soil. Microbiologyopen 2017, 6, e00502. [Google Scholar] [CrossRef]
- Nguyen, H.P.; Miwa, H.; Obirih-Opareh, J.; Suzaki, T.; Yasuda, M.; Okazaki, S. Novel rhizobia exhibit superior nodulation and biological nitrogen fixation even under high nitrate concentrations. FEMS Microbiol. Ecol. 2020, 96, fiz184. [Google Scholar] [CrossRef]
- Martiny, J.B.H.; Bohannan, B.J.M.; Brown, J.H.; Colwell, R.K.; Fuhrman, J.A.; Green, J.L.; Horner-Devine, M.C.; Kane, M.; Krumins, J.A.; Kuske, C.R.; et al. Microbial biogeography: Putting microorganisms on the map. Nat. Rev. Microbiol. 2006, 4, 102–112. [Google Scholar] [CrossRef]
- Bremner, J.M. Nitrogen-Total. In Methods of Soil Analysis; Soil Science Society of America: Madison, WI, USA, 1996; pp. 1085–1121. [Google Scholar]
- Bray, R.H.; Kurtz, L.T. Determination of Total, Organic, and Available Forms of Phosphorus in Soils. Soil Sci. 1945, 59, 39–46. [Google Scholar] [CrossRef]
- Mulvaney, R.L.; Khan, S.A. Diffusion Methods to Determine Different Forms of Nitrogen in Soil Hydrolysates. Soil Sci. Soc. Am. J. 2001, 65, 1284–1292. [Google Scholar] [CrossRef]
- Kalembasa, S.J.; Jenkinson, D.S. A comparative study of titrimetric and gravimetric methods for the determination of organic carbon in soil. J. Sci. Food Agric. 1973, 24, 1085–1090. [Google Scholar] [CrossRef]
- Cowan, D.A.; Sohm, J.A.; Makhalanyane, T.P.; Capone, D.G.; Green, T.G.A.; Cary, S.C.; Tuffin, I.M. Hypolithic communities: Important nitrogen sources in Antarctic desert soils. Environ. Microbiol. Rep. 2011, 3, 581–586. [Google Scholar] [CrossRef] [PubMed]
- Heller, P.; Tripp, H.J.; Turk-Kubo, K.; Zehr, J.P. ARBitrator: A software pipeline for on-demand retrieval of auto-curated nifH sequences from GenBank. Bioinformatics 2014, 30, 2883–2890. [Google Scholar] [CrossRef] [Green Version]
- Shearer, G.B.; Kohl, D.H. N2-Fixation in Field Settings: Estimations Based on Natural 15N Abundance. Aust. J. Plant Physiol. 1986, 13, 699–756. [Google Scholar]
- Chamkhi, I.; Cheto, S.; Geistlinger, J.; Zeroual, Y.; Kouisni, L.; Bargaz, A.; Ghoulam, C. Legume-based intercropping systems promote beneficial rhizobacterial community and crop yield under stressing conditions. Ind. Crops Prod. 2022, 183, 114958. [Google Scholar] [CrossRef]
- Reid, D.E.; Ferguson, B.J.; Hayashi, S.; Lin, Y.H.; Gresshoff, P.M. Molecular mechanisms controlling legume autoregulation of nodulation. Ann. Bot. 2011, 108, 789–795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Te, X.; Din, A.M.U.; Cui, K.; Raza, M.A.; Fraz Ali, M.; Xiao, J. Inter-specific root interactions and water use efficiency of maize/soybean relay strip intercropping. Field Crops Res. 2023, 291, 108793. [Google Scholar] [CrossRef]
- Wang, H.; Li, H.; Zhang, M.; Song, Y.; Huang, J.; Huang, H.; Shao, M.; Liu, Y.; Kang, Z. Carbon Dots Enhance the Nitrogen Fixation Activity of Azotobacter Chroococcum. ACS Appl. Mater. Interfaces 2018, 10, 16308–16314. [Google Scholar] [CrossRef]
- Lankau, R.A.; George, I.; Miao, M. Crop performance is predicted by soil microbial diversity across phylogenetic scales. Ecosphere 2022, 13, e4029. [Google Scholar] [CrossRef]
- Venieraki, A.; Dimou, M.; Pergalis, P.; Kefalogianni, I.; Chatzipavlidis, I.; Katinakis, P. The Genetic Diversity of Culturable Nitrogen-Fixing Bacteria in the Rhizosphere of Wheat. Microb. Ecol. 2011, 61, 277–285. [Google Scholar] [CrossRef]
- Tang, X.; Zhong, R.; Jiang, J.; He, L.; Huang, Z.; Shi, G.; Wu, H.; Liu, J.; Xiong, F.; Han, Z.; et al. Cassava/peanut intercropping improves soil quality via rhizospheric microbes increased available nitrogen contents. BMC Biotechnol. 2020, 20, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.D.; Feng, X.M.; Hu, Y.G.; Ren, C.Z.; Zeng, Z.H. Effects of legume-oat intercropping on abundance and community structure of soil N2-fixing bacteria. J. Appl. Ecol. 2017, 28, 957–965. [Google Scholar] [CrossRef]
- Nourbakhsh, F.; Koocheki, A.; Mahallati, M.N. Investigation of Biodiversity and Some of the Ecosystem Services in the Intercropping of Corn, Soybean and Marshmallow. Int. J. Plant Prod. 2019, 13, 35–46. [Google Scholar] [CrossRef]
- Koocheki, A.; Solouki, H.; Karbor, S. Study of ecological aspects of Sesame (Sesamum indicum L.) and Mung Bean (Vigna radiata L.) intercropping in weed control. Iran. J. Pulses Res. 2016, 7, 27–44. [Google Scholar] [CrossRef]
- Ren, N.; Wang, Y.; Ye, Y.; Zhao, Y.; Huang, Y.; Fu, W.; Chu, X. Effects of Continuous Nitrogen Fertilizer Application on the Diversity and Composition of Rhizosphere Soil Bacteria. Front. Microbiol. 2020, 11, 1948. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Jiang, X.; Wei, D.; Zhao, B.; Ma, M.; Chen, S.; Cao, F.; Shen, D.; Guan, D.; Li, J. Consistent effects of nitrogen fertilization on soil bacterial communities in black soils for two crop seasons in China. Sci. Rep. 2017, 7, 3267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antoninka, A.; Reich, P.B.; Johnson, N.C. Seven years of carbon dioxide enrichment, nitrogen fertilization and plant diversity influence arbuscular mycorrhizal fungi in a grassland ecosystem. New Phytol. 2011, 192, 200–214. [Google Scholar] [CrossRef]
- Liu, Y.; Shi, G.; Mao, L.; Cheng, G.; Jiang, S.; Ma, X.; An, L.; Du, G.; Collins Johnson, N.; Feng, H. Direct and indirect influences of 8 yr of nitrogen and phosphorus fertilization on Glomeromycota in an alpine meadow ecosystem. New Phytol. 2012, 194, 523–535. [Google Scholar] [CrossRef]
- Sun, L.; Song, F.; Liu, S.; Cao, Q.; Liu, F.; Zhu, X. Integrated agricultural management practice improves soil quality in Northeast China. Arch. Agron. Soil Sci. 2018, 64, 1932–1943. [Google Scholar] [CrossRef]
- Jia, Y.; Liao, Z.; Chew, H.; Wang, L.; Lin, B.; Chen, C.; Lu, G.; Lin, Z. Effect of Pennisetum giganteum z.x.lin mixed nitrogen-fixing bacterial fertilizer on the growth, quality, soil fertility and bacterial community of pakchoi (Brassica chinensis L.). PLoS ONE 2020, 15, e0228709. [Google Scholar] [CrossRef] [Green Version]
- Sepp, S.K.; Vasar, M.; Davison, J.; Oja, J.; Anslan, S.; Al-Quraishy, S.; Bahram, M.; Bueno, C.G.; Cantero, J.J.; Fabiano, E.C.; et al. Global diversity and distribution of nitrogen-fixing bacteria in the soil. Front. Plant Sci. 2023, 14, 1100235. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.; Zhang, J.; Yang, D.; Chen, X.; Zhao, J.; Xiu, W.; Lai, X.; Li, G. Effects of different land use patterns on nifH genetic diversity of soil nitrogen-fixing microbial communities in Leymus Chinensis steppe. Acta Ecol. Sin. 2011, 31, 150–156. [Google Scholar] [CrossRef]
- Li, Y.; Pan, F.; Yao, H. Response of symbiotic and asymbiotic nitrogen-fixing microorganisms to nitrogen fertilizer application. J. Soils Sediments 2019, 19, 1948–1958. [Google Scholar] [CrossRef]
- Kubota, M.; Matsushita, N.; Nakamura, T.; Fukuda, K. Nitrogen fixation and nifH gene diversity in cyanobacteria living on feather mosses in a subalpine forest of Mt. Fuji. Oecologia 2023, 201, 749–760. [Google Scholar] [CrossRef]
- Din, I.; Khan, H.; Ahmad Khan, N.; Khil, A. Inoculation of nitrogen fixing bacteria in conjugation with integrated nitrogen sources induced changes in phenology, growth, nitrogen assimilation and productivity of wheat crop. J. Saudi Soc. Agric. Sci. 2021, 20, 459–466. [Google Scholar] [CrossRef]
- Wang, H.; Gu, C.; Liu, X.; Yang, C.; Li, W.; Wang, S. Impact of Soybean Nodulation Phenotypes and Nitrogen Fertilizer Levels on the Rhizosphere Bacterial Community. Front. Microbiol. 2020, 11, 750. [Google Scholar] [CrossRef]
- Gu, Y.; Wang, J.; Cai, W.; Li, G.; Mei, Y.; Yang, S. Different Amounts of Nitrogen Fertilizer Applications Alter the Bacterial Diversity and Community Structure in the Rhizosphere Soil of Sugarcane. Front. Microbiol. 2021, 12, 721441. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | No.of Nodule Per Plant (no/Plant) | DM of Nodule Per Plant (g/Plant) | ||||
|---|---|---|---|---|---|---|
| SS | IS | AVE | SS | IS | AVE | |
| N0 | 196 ± 6.02 b | 218 ± 4.35 a | 207 ± 17.91 A | 0.87 ± 0.15 a | 0.99 ± 0.16 a | 0.93 ± 0.15 A |
| N1 | 164 ± 9.45 b | 209 ± 3.51 a | 187 ± 13.47 B | 0.80 ± 0.09 a | 0.88 ± 0.02 a | 0.84 ± 0.07 A |
| N2 | 119 ± 11.2 b | 151 ± 14.4 a | 135 ± 21.16 C | 0.51 ± 0.01 a | 0.57 ± 0.11 a | 0.54 ± 0.08 B |
| ANOVE | p | p | ||||
| N | 0.00 | 0.00 | ||||
| C | 0.00 | 0.11 | ||||
| N×C | 0.03 | 0.93 | ||||
| Treatments | AN (mg/kg) | AP (mg/kg) | AK (mg/kg) | pH | OM (g/kg) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| SS | IS | AVE | SS | IS | AVE | SS | IS | AVE | SS | IS | AVE | SS | IS | AVE | |
| N0 | 132 ± 1.52 a | 131 ± 4.35 a | 131 ± 3.01 B | 71 ± 0.55 b | 70 ± 0.26 a | 70 ± 0.48 B | 141 ± 1.07 a | 143 ± 2.50 a | 142 ± 1.85 A | 6.66 ± 0.08 a | 6.60 ± 0.03 a | 6.63 ± 0.06 B | 30 ± 0.65 a | 30 ± 1.03 a | 30 ± 0.77 A |
| N1 | 132 ± 3.51 a | 132 ± 1.01 a | 132 ± 2.31 B | 71 ± 0.14 b | 70 ± 0.39 a | 71 ± 0.32 A | 141 ± 1.55 a | 143 ± 1.12 a | 142 ± 1.39 A | 6.67 ± 0.08 a | 6.65 ± 0.08 a | 6.66 ± 0.07 AB | 31 ± 0.65 a | 31 ± 1.79 a | 31 ± 0.68 A |
| N2 | 155 ± 2.02 a | 154 ± 4.93 a | 155 ± 3.37 A | 72 ± 0.56 b | 71 ± 0.05 a | 71 ± 0.57 A | 142 ± 0.57 a | 142 ± 2.95 a | 142 ± 1.90 A | 6.75 ± 0.02 a | 6.72 ± 0.02 a | 6.73 ± 0.02 A | 31 ± 0.28 a | 31 ± 0.97 a | 31 ± 0.70 A |
| ANOVE | p | p | p | p | p | ||||||||||
| N | 0.00 | 0.01 | 0.95 | 0.03 | 0.29 | ||||||||||
| C | 0.67 | 0.03 | 0.40 | 0.25 | 0.35 | ||||||||||
| N×C | 0.95 | 0.59 | 0.71 | 0.84 | 0.89 | ||||||||||
| Soil Characteristic | RDA1 | RDA2 | r2 | p |
|---|---|---|---|---|
| AN | 0.8445 | 0.5356 | 0.2044 | 0.176 |
| OM | 0.3247 | 0.9458 | 0.0527 | 0.683 |
| AP | 0.8044 | 0.594 | 0.2444 | 0.114 |
| AK | −0.5725 | −0.8199 | 0.0502 | 0.681 |
| pH | 0.9566 | 0.2915 | 0.0672 | 0.577 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, Z.; Meng, L.; Yin, T.; Li, Y.; Zhang, Y.; Li, S. Changes in Soil Rhizobia Diversity and Their Effects on the Symbiotic Efficiency of Soybean Intercropped with Maize. Agronomy 2023, 13, 997. https://doi.org/10.3390/agronomy13040997
Cheng Z, Meng L, Yin T, Li Y, Zhang Y, Li S. Changes in Soil Rhizobia Diversity and Their Effects on the Symbiotic Efficiency of Soybean Intercropped with Maize. Agronomy. 2023; 13(4):997. https://doi.org/10.3390/agronomy13040997
Chicago/Turabian StyleCheng, Zeyu, Lingbo Meng, Tengjiao Yin, Ying Li, Yuhang Zhang, and Shumin Li. 2023. "Changes in Soil Rhizobia Diversity and Their Effects on the Symbiotic Efficiency of Soybean Intercropped with Maize" Agronomy 13, no. 4: 997. https://doi.org/10.3390/agronomy13040997
APA StyleCheng, Z., Meng, L., Yin, T., Li, Y., Zhang, Y., & Li, S. (2023). Changes in Soil Rhizobia Diversity and Their Effects on the Symbiotic Efficiency of Soybean Intercropped with Maize. Agronomy, 13(4), 997. https://doi.org/10.3390/agronomy13040997