Characterization and Gene Mapping of non-open hull 1 (noh1) Mutant in Rice (Oryza sativa L.)

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Morphological and Histological Analysis of noh1
2.3. Molecular Mapping of NOH1
2.4. Linkage Map Construction
2.5. qPCR Analysis
3. Results
3.1. Morphological Analysis of noh1 Mutant
3.2. Histological Analysis of noh1 Mutant
3.3. Morphological Analysis of noh1 Lodicules during Flowering
3.4. Expression Analysis of Floral Organ Identity Genes in Lodicule of noh1 and Wild-Type
3.5. Genetic Analysis
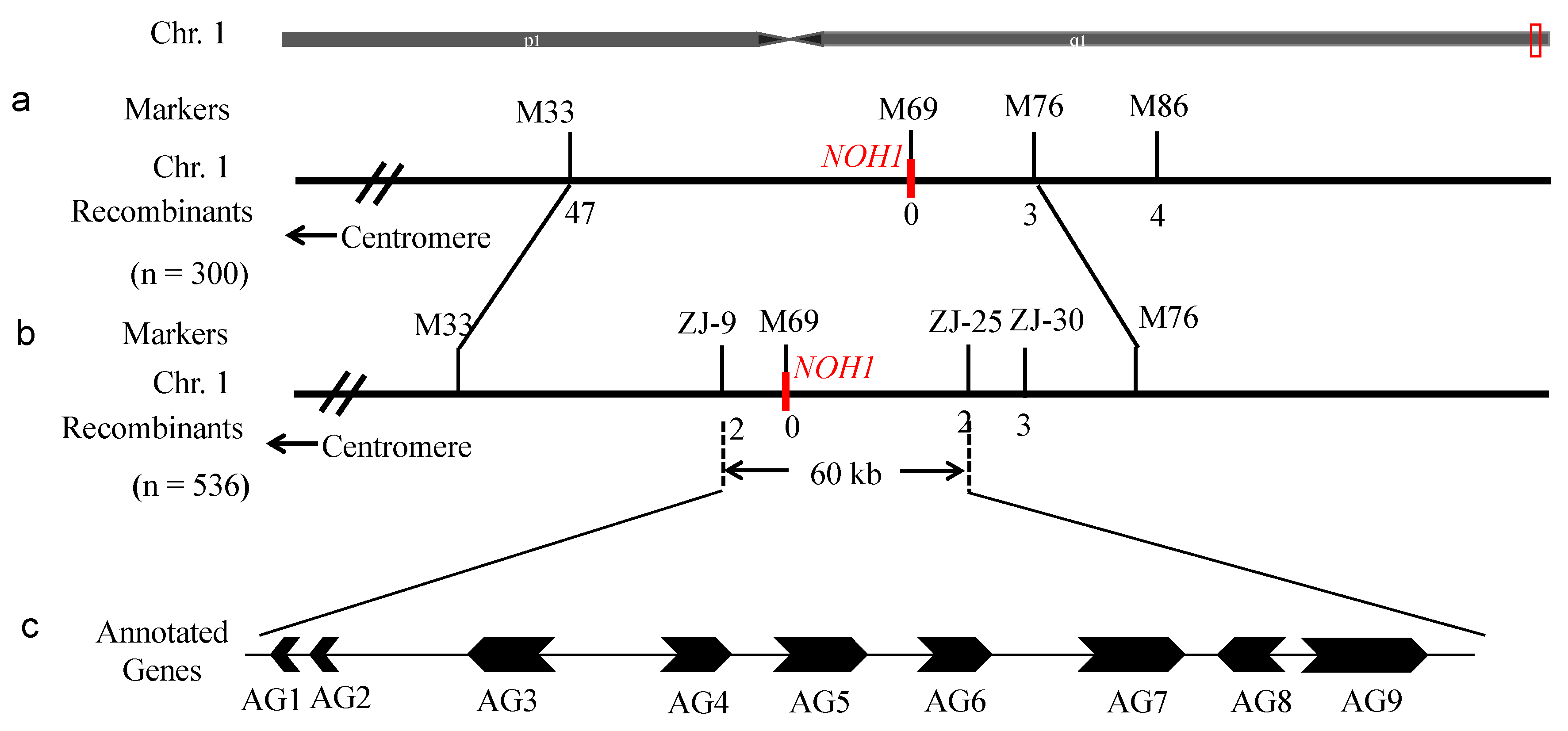
3.6. Gene Mapping of NOH1
4. Discussion and Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Yoshida, H.; Nagato, Y. Flower development in rice. J. Exp. Bot. 2011, 62, 4719–4730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, X.; Zhou, X.; Wu, X. Advances in study of opening mechanism in rice florets. Sci. Agric. Sin. 2004, 37, 188–195. [Google Scholar] [CrossRef]
- Wang, Z.; Gu, Y.; Gao, Y. Studies on the mechanism of the anthesis of rice Ⅲ. Structure of the lodicule and changes of its contents during flowering. Acta Agron. Sin. 1991, 17, 96–101. (In Chinese) [Google Scholar]
- Xiao, H.; Wang, Y.; Liu, D.; Wang, W.; Li, X.; Zhao, X.; Xu, J.; Zhai, W.; Zhu, L. Functional analysis of the rice AP3 homologue OsMADS16 by RNA interference. Plant Mol. Biol. 2003, 52, 957–966. [Google Scholar] [CrossRef] [PubMed]
- Yadav, S.R.; Prasad, K.; Vijayraghavan, U. Divergent regulatory OsMADS2 functions control size, shape and differentiation of the highly derived rice floret second-whorl organ. Genetics 2007, 176, 283–294. [Google Scholar] [CrossRef]
- Yao, S.G.; Ohmori, S.; Kimizu, M.; Yoshida, H. Unequal genetic redundancy of rice PISTILLATA orthologs, OsMADS2 and OsMADS4, in lodicule and stamen development. Plant Cell Physiol. 2008, 49, 853–857. [Google Scholar] [CrossRef]
- Li, H.; Liang, W.; Jia, R.; Yin, C.; Zong, J.; Kong, H.; Zhang, D. The AGL6-like gene OsMADS6 regulates floral organ and meristem identities in rice. Cell Res. 2010, 20, 299–313. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, L.; Cai, Q.; Hu, Y.; Jin, Z.; Zhao, X.; Fan, W.; Huang, Q.; Luo, Z.; Chen, M.; et al. OsMADS32 interacts with PI-like proteins and regulates rice flower development. J. Integr. Plant Biol. 2015, 57, 504–513. [Google Scholar] [CrossRef]
- Xiao, H.; Tang, J.; Li, Y.; Wang, W.; Li, X.; Jin, L.; Xie, R.; Luo, H.; Zhao, X.; Meng, Z. STAMENLESS 1, encoding a single C2H2 zinc finger protein, regulates floral organ identity in rice. Plant J. 2009, 59, 789–801. [Google Scholar] [CrossRef]
- Michelmore, R.W.; Paran, I.; Kesseli, R.V. Identification of markers linked to disease-resistance genes by bulked segregant analysis: A rapid method to detect markers in specific genomic regions by using segregating populations. Proc. Natl. Acad. Sci. USA 1991, 88, 9828–9832. [Google Scholar] [CrossRef]
- Murray, M.G.; Thompson, W.F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Res. 1980, 8, 4321–4325. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.; Yang, Z.; Zhong, B.; Li, Y.; Xie, R.; Zhao, F.; Ling, Y.; He, G. Genetic analysis and fine mapping of a dynamic rolled leaf gene, RL10(t), in rice (Oryza sativa L.). Genome 2007, 50, 811–817. [Google Scholar] [CrossRef] [PubMed]
- Lander, E.S.; Green, P.; Abrahamson, J.; Barlow, A.; Daly, M.J.; Lincoln, S.E.; Newberg, L.A. MAPMAKER: An interactive computer package for constructing primary genetic linkage maps of experimental and natural populations. Genomics 1987, 1, 174–181. [Google Scholar] [CrossRef]
- Yamaguchi, T.; Nagasawa, N.; Kawasaki, S.; Matsuoka, M.; Nagato, Y.; Hirano, H.Y. The YABBY gene DROOPING LEAF regulates carpel specification and midrib development in Oryza sativa. Plant Cell 2004, 16, 500–509. [Google Scholar] [CrossRef] [PubMed]
- Nagasawa, N.; Miyoshi, M.; Sano, Y.; Satoh, H.; Hirano, H.; Sakai, H.; Nagato, Y. SUPERWOMAN1 and DROOPING LEAF genes control floral organ identity in rice. Development 2003, 130, 705–718. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, A.; Suzaki, T.; Tanaka, W.; Hirano, H.Y. The homeotic gene long sterile lemma (G1) specifies sterile lemma identity in the rice spikelet. Proc. Natl. Acad. Sci. USA 2009, 106, 20103–20108. [Google Scholar] [CrossRef]
- Yoshida, A.; Ohmori, Y.; Kitano, H.; Taguchi-Shiobara, F.; Hirano, H.Y. ABERRANT SPIKELET AND PANICLE1, encoding a TOPLESS-related transcriptional co-repressor, is involved in the regulation of meristem fate in rice. Plant J. 2012, 70, 327–339. [Google Scholar] [CrossRef]
- Pelucchi, N.; Fornara, F.; Favalli, C.; Masiero, S.; Lago, C.; Pè, M.E.; Colombo, L.; Kater, M.M. Comparative analysis of rice MADS-box genes expressed during flower development. Sex. Plant Reprod. 2002, 15, 113–122. [Google Scholar] [CrossRef]
- Wang, K.J.; Tang, D.; Hong, L. L.; Xu, W.Y.; Huang, J.; Li, M.; Gu, M.H.; Xue, Y.B.; Cheng, Z.K. DEP and AFO Regulate Reproductive Habit in Rice. PLoS Genet. 2010, 6, e1000818. [Google Scholar] [CrossRef]
- Prasad, K.; Parameswaran, S.; Vijayraghavan, U. OsMADS1, a rice MADS-box factor, controls differentiation of specific cell types in the lemma and palea and is an early-acting regulator of inner floral organs. Plant J. 2005, 43, 915–928. [Google Scholar] [CrossRef]
- Gao, X.; Liang, W.; Yin, C.; Ji, S.; Wang, H.; Su, X.; Guo, C.; Kong, H.; Xue, H.; Zhang, D. The SEPALLATA-like gene OsMADS34 is required for rice inflorescence and spikelet development. Plant Physiol. 2010, 153, 728–740. [Google Scholar] [CrossRef] [PubMed]
- Bommert, P.; Satohnagasawa, N.; Jackson, D.; Hirano, H.Y. Genetics and evolution of inflorescence and flower development in grasses. Plant Cell Physiol. 2005, 46, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Whipple, C.J.; Zanis, M.J.; Kellogg, E.A.; Schmidt, R.J. Conservation of B class gene expression in the second whorl of a basal grass and outgroups links the origin of lodicules and petals. Proc. Natl. Acad. Sci. USA 2007, 104, 1081–1086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prasad, K.; Vijayraghavan, U. Double-stranded RNA interference of a rice PI/GLO paralog, OsMADS2, uncovers its second-whorl-specific function in floral organ patterning. Genetics 2003, 165, 2301–2305. [Google Scholar] [CrossRef] [PubMed]
- Ohmori, S.; Kimizu, M.; Sugita, M.; Miyao, A.; Hirochika, H.; Uchida, E.; Nagato, Y.; Yoshida, H. MOSAIC FLORAL ORGANS1, an AGL6-like MADS box gene, regulates floral organ identity and meristem fate in rice. Plant Cell 2009, 21, 3008–3025. [Google Scholar] [CrossRef] [PubMed]
- Sang, X.; Li, Y.; Luo, Z.; Ren, D.; Fang, L.; Wang, N.; Zhao, F.; Ling, Y.; Yang, Z.; Liu, Y.; He, G. CHIMERIC FLORAL ORGANS 1, encoding a Monocot-specific MADS-box Protein, Regulates Floral Organ Identity in Rice. Plant Physiol. 2012, 160, 788–807. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, H.; Itoh, J.I.; Ohmori, S.; Miyoshi, K.; Horigome, A.; Uchida1, E.; Kimizu, M.; Matsumura, Y.; Kusaba, M.; Satoh, H.; Nagato, Y. superwoman1-cleistogamy, a hopeful allele for gene containment in GM rice. Plant Biotechnol J. 2007, 5, 835–846. [Google Scholar] [CrossRef] [PubMed]
- Kellogg, E.A. The evolutionary history of Ehrhartoideae, Oryzeae, and Oryza. Rice 2009, 2, 1–14. [Google Scholar] [CrossRef]
- Ren, D.; Li, Y.; Zhao, F.; Sang, X.; Shi, J.; Wang, N.; Guo, S.; Ling, Y.; Zhang, C.; Yang, Z.; He, G. MULTI-FLORET SPIKELET1, which encodes an AP2/ERF protein, determines spikelet meristem fate and sterile lemma identity in rice. Plant Physiol. 2013, 162, 872–884. [Google Scholar] [CrossRef] [PubMed]
- Hong, L.; Qian, Q.; Zhu, K.; Tang, D.; Huang, Z.; Gao, L.; Li, M.; Gu, M.; Cheng, Z. ELE restrains empty glumes from developing into lemmas. J. Genet. Genom. 2010, 37, 101–115. [Google Scholar] [CrossRef]
- Kobayashi, K.; Maekawa, M.; Miyao, A.; Hirohiko, H.; Kyozuka, J. PANICLE PHYTOMER2 (PAP2), encoding a SEPALLATA subfamily MADS-box protein, positively controls spikelet meristem identity in rice. Plant Cell Physiol. 2010, 51, 47–57. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Wu, F.; Du, X.; Shi, X.; Liu, Y.; Liu, S.; Hu, Y.; Theißen, G.; Meng, Z. The pleiotropic SEPALLATA-like gene OsMADS34 reveals that the ‘empty glumes’of rice (Oryza sativa) spikelets are in fact rudimentary lemmas. New Phytol. 2014, 202, 689–702. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Yoshida, A.; Takahashi, M.; Maekawa, M.; Kojima, M.; Sakakibara, H.; Kyozuka, J. SAD1, an RNA polymerase I subunit A34.5 of rice, interacts with Mediator and controls various aspects of plant development. Plant J. 2015, 81, 282–291. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Qiao, M.; Liu, Z.; Xiang, F. Diversification function of WRKY transcription factor. Chin. Bull. Life Sci. 2010, 22, 345–351. [Google Scholar] [CrossRef]
- Nair, S.K.; Wang, N.; Turuspekov, Y.; Pourkheirandish, M.; Sinsuwongwat, S.; Chen, G.; Sameri, M.; Tagiri, A.; Honda, I.; Watanabe, Y.; et al. Cleistogamous Flowering in Barley Arises from the Suppression of MicroRNA-Guided HvAP2 mRNA Cleavage. Proc. Natl. Acad. Sci. USA 2010, 107, 490–495. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Forward Sequence (5′–3′) | Revers Sequence (5′–3′) |
|---|---|---|
| M33 | CTTGAGTTCGAAGCGAGAAGACG | CACTTGAGCTCGAGACGTAGCC |
| ZJ-9 | CAGATGGAGTACATGAAGTGCCAATG | GCATTGTGTCAACAACTCAGGTCC |
| ZJ-25 | CACGGTAATGTGCTAAAGCTCCTG | GTGGGTTGTGGAGAGACAACCTG |
| ZJ-30 | CAAGAAGCTCAACCAGGACGGCTTC | GAGAGTAGAGTTGAGGCACCGAATCG |
| M69 | CTCTACAGCTTGAGTTTGGTACATCC | GTGTTGGTGAGCTAGCTGTTGC |
| M76 | GTCGACGGCTTCCTCAAGATTGG | TGAGACCTCTGTGAAGGCACTCG |
| M86 | CTCACTCACTGACCCACAACTCC | TTAAGATGATGGCTCCTCTCTGC |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Zheng, H.; Zeng, X.; Zhuang, H.; Wang, H.; Tang, J.; Chen, H.; Ling, Y.; Li, Y. Characterization and Gene Mapping of non-open hull 1 (noh1) Mutant in Rice (Oryza sativa L.). Agronomy 2019, 9, 56. https://doi.org/10.3390/agronomy9020056
Zhang J, Zheng H, Zeng X, Zhuang H, Wang H, Tang J, Chen H, Ling Y, Li Y. Characterization and Gene Mapping of non-open hull 1 (noh1) Mutant in Rice (Oryza sativa L.). Agronomy. 2019; 9(2):56. https://doi.org/10.3390/agronomy9020056
Chicago/Turabian StyleZhang, Jun, Hao Zheng, Xiaoqin Zeng, Hui Zhuang, Honglei Wang, Jun Tang, Huan Chen, Yinghua Ling, and Yunfeng Li. 2019. "Characterization and Gene Mapping of non-open hull 1 (noh1) Mutant in Rice (Oryza sativa L.)" Agronomy 9, no. 2: 56. https://doi.org/10.3390/agronomy9020056
APA StyleZhang, J., Zheng, H., Zeng, X., Zhuang, H., Wang, H., Tang, J., Chen, H., Ling, Y., & Li, Y. (2019). Characterization and Gene Mapping of non-open hull 1 (noh1) Mutant in Rice (Oryza sativa L.). Agronomy, 9(2), 56. https://doi.org/10.3390/agronomy9020056