Elevated CO2 Levels Impact Fitness Traits of Vine Mealybug Planococcus ficus Signoret, but Not Its Parasitoid Leptomastix dactylopii Howard



Abstract
:1. Introduction
2. Materials and Methods
3. Results
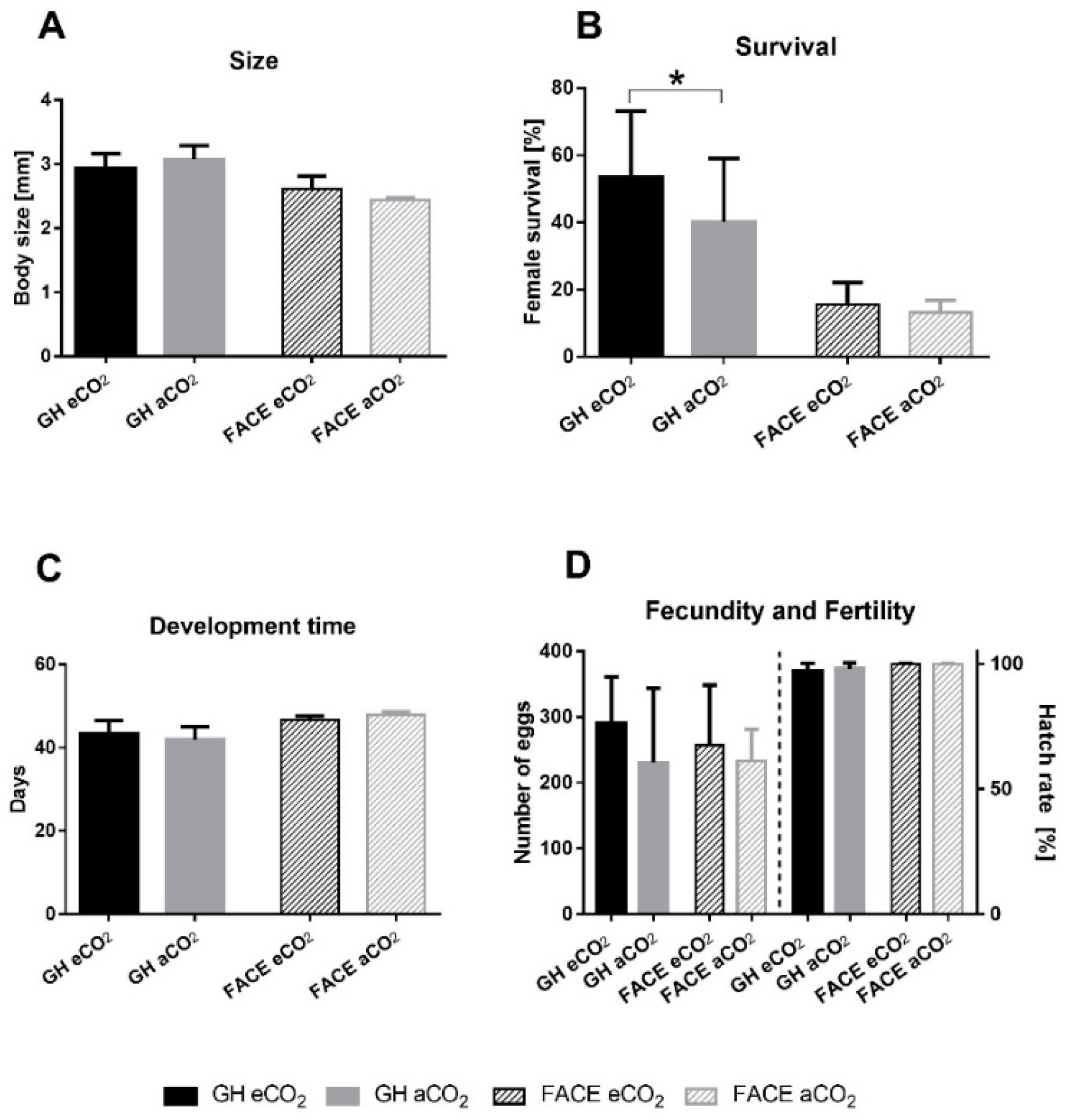
3.1. Life History Parameters of P. ficus Under Different CO2 Levels
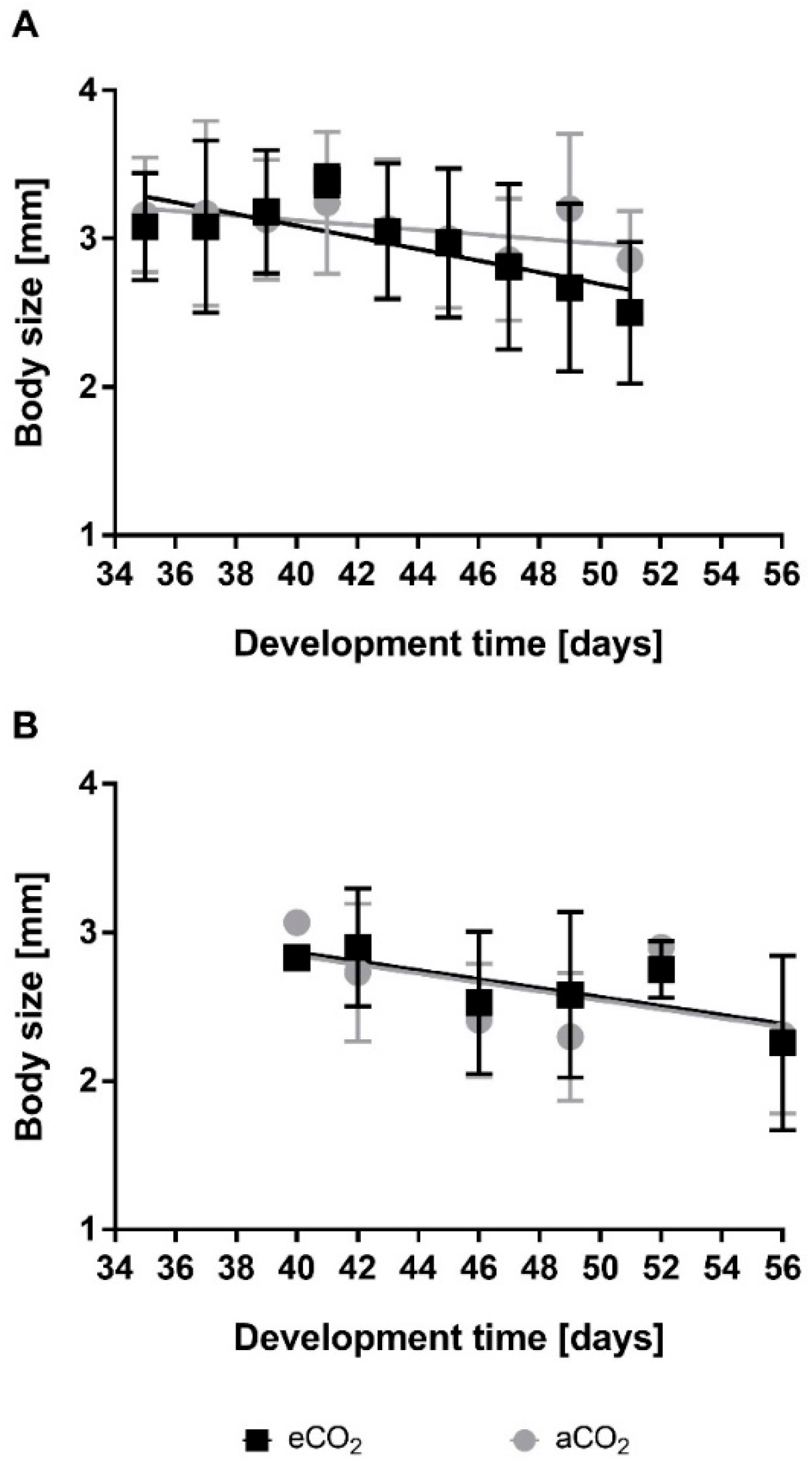
3.1.1. Greenhouse Experiments
3.1.2. VineyardFACE Experiments
3.2. Parasitism by L. dactylopii
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Pachauri, R.K.; Allen, M.R.; Barros, V.R.; Broome, J.; Cramer, W.; Christ, R.; Church, J.A.; Clarke, L.; Dahe, Q.; Dasgupta, P.; et al. IPCC Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; IPCC: Geneva, Switzerland, 2014; Available online: https://epic.awi.de/id/eprint/37530/ (accessed on 31 January 2019).
- Long, S.P.; Ainsworth, E.A.; Rogers, A.; Ort, D.R. Rising atmospheric carbon dioxide: Plants FACE the future. Annu. Rev. Plant Biol. 2004, 55, 591–628. [Google Scholar] [CrossRef] [PubMed]
- Robinson, E.A.; Ryan, G.D.; Newman, J.A. A meta-analytical review of the effects of elevated CO2 on plant-arthropod interactions highlights the importance of interacting environmental and biological variables. New Phytol. 2012, 194, 321–336. [Google Scholar] [CrossRef] [PubMed]
- Bindi, M.; Fibbi, L.; Miglietta, F. Free air CO2 enrichment (FACE) of grapevine (Vitis vinifera L.): II. Growth and quality of grape and wine in response to elevated CO2 concentrations. Eur. J. Agron. 2001, 14, 145–155. [Google Scholar] [CrossRef]
- Wohlfahrt, Y.; Smith, J.P.; Tittmann, S.; Honermeier, B.; Stoll, M. Primary productivity and physiological responses of Vitis vinifera L. cvs. under Free Air Carbon dioxide Enrichment (FACE). Eur. J. Agron. 2018, 101, 149–162. [Google Scholar] [CrossRef]
- Reineke, A.; Thiéry, D. Grapevine insect pests and their natural enemies in the age of global warming. J. Pest Sci. (2004) 2016, 89, 313–328. [Google Scholar] [CrossRef]
- Bezemer, T.M.; Jones, T.H. Plant-insect herbivore interactions in elevated atmospheric CO2: Quantitative analyses and guild effects. Oikos 1998, 82, 212–222. [Google Scholar] [CrossRef]
- Sudderth, E.A.; Stinson, K.A.; Bazzaz, F.A. Host-specific aphid population responses to elevated CO2 and increased N availability. Glob. Chang. Biol. 2005, 11, 1997–2008. [Google Scholar] [CrossRef]
- Walton, V.M.; Pringle, K.L. Vine mealybug, Planococcus ficus (Signoret) (Hemiptera: Pseudococcidae), a key pest in South African vineyards: A Review. S. Afr. J. Enol. Vitic. 2004, 25, 54–62. [Google Scholar] [CrossRef]
- Daane, K.M.; Almeida, R.P.P.; Bell, V.A.; Walker, J.T.S.; Botton, M.; Fallahzadeh, M.; Mani, M.; Miano, J.L.; Sforza, R.; Walton, V.M. Biology and management of mealybugs in vineyards. In Arthropod Management in Vineyards; Bostanian, N.J., Vincent, C., Isaacs, R., Eds.; Springer: New York, NY, USA, 2012; pp. 271–307. [Google Scholar]
- Chiotta, M.L.; Ponsone, M.L.; Torres, A.M.; Combina, M.; Chulze, S.N. Influence of Planococcus ficus on Aspergillus section Nigri and ochratoxin A incidence in vineyards from Argentina. Lett. Appl. Microbiol. 2010, 51, 212–218. [Google Scholar] [CrossRef]
- Bordeu, E.; Troncoso, D.O.; Zaviezo, T. Influence of mealybug (Pseudococcus spp.)-infested bunches on wine quality in Carmenere and Chardonnay grapes. Int. J. Food Sci. Technol. 2012, 47, 232–239. [Google Scholar] [CrossRef]
- Engelbrecht, D.J.; Kasdorf, G.G.F. Field spreas of corky bark, fleck, leafroll and Shiraz decline diseases and associated viruses in South African grapevines. Phytophylactica 1990, 22, 347–354. [Google Scholar]
- Almeida, R.P.P.; Daane, K.M.; Bell, V.A.; Blaisdell, G.K.; Cooper, M.L.; Herrbach, E.; Pietersen, G. Ecology and management of grapevine leafroll disease. Front. Microbiol. 2013, 4, 94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becerra, V.; Gonzalez, M.; Herrera, M.E.; Miano, J.L. Population dynamics of vine mealybug Planococcus ficus Sign. in vineyards. Mendoza. Rev. Fac. Ciencias Agrar. Univ. Nac. Cuyo 2006, 1, 1–6. [Google Scholar]
- Prabhaker, N.; Gispert, C.; Castle, S.J. Baseline susceptibility of Planococcus ficus (Hemiptera: Pseudococcidae) from California to select insecticides. J. Econ. Entomol. 2012, 105, 1392–1400. [Google Scholar] [CrossRef] [PubMed]
- Venkatesan, T.; Jalali, S.K.; Ramya, S.L.; Prathibha, M. Insecticide resistance and its management in mealybugs. In Mealybugs and Their Management in Agricultural and Horticultural crops; Springer: New Delhi, India, 2016; pp. 223–229. [Google Scholar]
- Muscas, E.; Cocco, A.; Mercenaro, L.; Cabras, M.; Lentini, A.; Porqueddu, C.; Nieddu, G. Effects of vineyard floor cover crops on grapevine vigor, yield, and fruit quality, and the development of the vine mealybug under a Mediterranean climate. Agric. Ecosyst. Environ. 2017, 237, 203–212. [Google Scholar] [CrossRef]
- Cocco, A.; Muscas, E.; Mura, A.; Iodice, A.; Savino, F.; Lentini, A. Influence of mating disruption on the reproductive biology of the vine mealybug, Planococcus ficus (Hemiptera: Pseudococcidae), under field conditions. Pest Manag. Sci. 2018, 74, 2806–2816. [Google Scholar] [CrossRef] [PubMed]
- Walton, V.M.; Daane, K.M.; Bentley, W.J.; Millar, J.G.; Larsen, T.E.; Malakar-Kuenen, R. Pheromone-based mating disruption of Planococcus ficus (Hemiptera: Pseudococcidae) in California vineyards. J. Econ. Entomol. 2006, 99, 1280–1290. [Google Scholar] [CrossRef]
- Mansour, R.; Grissa Lebdi, K.; Rezgui, S. Assessment of the performance of some new insecticides for the control of the vine mealybug Planococcus ficus in a Tunisian vineyard. Entomol. Hell. 2017, 19, 21–33. [Google Scholar] [CrossRef]
- Daane, K.M.; Bentley, W.J.; Walton, V.M.; Malakar-Kuenen, R.; Millar, J.G.; Ingels, C.A.; Weber, E.A.; Gispert, C. New controls investigated for vine mealybug. Calif. Agric. 2006, 60, 31–38. [Google Scholar] [CrossRef]
- Gutierrez, A.P.; Ponti, L.; D’Oultremont, T.; Ellis, C.K. Climate change effects on poikilotherm tritrophic interactions. Clim. Chang. 2008, 87, 167–192. [Google Scholar] [CrossRef]
- Paul Gutierrez, A.; Daane, K.M.; Ponti, L.; Walton, V.M.; Ellis, C.K. Prospective evaluation of the biological control of vine mealybug: Refuge effects and climate. J. Appl. Ecol. 2008, 45, 524–536. [Google Scholar] [CrossRef]
- Chong, J.-H.; van lersel, M.W.; Oetting, R.D. Effects of elevated carbon dioxide levels and temperature on the life history of the Madeira mealybug (Hemiptera: Pseudococcidae). J. Entomol. Sci. 2004, 39, 387–397. [Google Scholar] [CrossRef]
- Kol-Maimon, H.; Ghanim, M.; Franco, J.C.; Mendel, Z. Evidence for gene flow between two sympatric mealybug species (Insecta; Coccoidea; Pseudococcidae). PLoS ONE 2014, 9, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Noyes, J.S.; Hayat, M. Oriental mealybug parasitoids of the Anagyrini (Hymenoptera: Encyrtidae); CAB International: Wallingford, UK, 1994. [Google Scholar]
- Jong, P.W.; Alphen, J.J.M. Host size selection and sex allocation in Leptomastix dactylopii, a parasitoid of Planococcus citri. Entomol. Exp. Appl. 1989, 50, 161–169. [Google Scholar] [CrossRef]
- Krishnamoorthy, A.; Singh, S.P. Biological control of citrus mealybug, Planococcus citri with an introduced parasite, Leptomastix dactylopii in India. Entomophaga 1987, 32, 143–148. [Google Scholar] [CrossRef]
- Fallahzadeh, M.; Japoshvili, G.; Saghaei, N.; Daane, K.M. Natural enemies of Planococcus ficus (Hemiptera: Pseudococcidae) in Fars Province vineyards, Iran. Biocontrol Sci. Technol. 2011, 21, 427–433. [Google Scholar] [CrossRef]
- Mahfoudhi, N.; Dhouibi, M.H. Survey of Mealybugs (Hemiptera: Pseudococcidae) and their Natural Enemies in Tunisian Vineyards. Afr. Entomol. 2009, 17, 154–160. [Google Scholar] [CrossRef]
- Daane, K.M.; Cooper, M.L.; Triapitsyn, S.V.; Andrews, J.W.; Ripa, R. Parasitoids of obscure mealybug, Pseudococcus viburni (Hem.: Pseudococcidae) in California: Establishment of Pseudaphycus flavidulus (Hym.: Encyrtidae) and discussion of related parasitoid species. Biocontrol Sci. Technol. 2008, 18, 43–57. [Google Scholar] [CrossRef]
- Marras, P.M.; Cocco, A.; Muscas, E.; Lentini, A. Laboratory evaluation of the suitability of vine mealybug, Planococcus ficus, as a host for Leptomastix dactylopii. Biol. Control 2016, 95, 57–65. [Google Scholar] [CrossRef]
- Hentley, W.T.; Wade, R.N. Global change, herbivores and their natural enemies. In Global Climate Change and Terrestrial Invertebrates; John Wiley & Sons, Ltd.: Chichester, UK, 2016; pp. 177–200. [Google Scholar]
- Facey, S.L.; Ellsworth, D.S.; Staley, J.T.; Wright, D.J.; Johnson, S.N. Upsetting the order: How climate and atmospheric change affects herbivore–enemy interactions. Curr. Opin. Insect Sci. 2014, 5, 66–74. [Google Scholar] [CrossRef]
- Mumm, R.; Dicke, M. Variation in natural plant products and the attraction of bodyguards involved in indirect plant defense. Can. J. Zool. 2010, 88, 628–667. [Google Scholar] [CrossRef]
- Timm, A.E.; Reineke, A. First insights into grapevine transcriptional responses as a result of vine mealybug Planococcus ficus feeding. Arthropod. Plant. Interact. 2014, 8, 495–505. [Google Scholar] [CrossRef]
- DeLucia, E.H.; Nabity, P.D.; Zavala, J.A.; Berenbaum, M.R. Climate change: Resetting plant-insect interactions. Plant Physiol. 2012, 160, 1677–1685. [Google Scholar] [CrossRef] [PubMed]
- Boudon-Padieu, É.; Maixner, M. Potential effects of climate change on distribution and activity of insect vectors of grapevine pathogens. In Proceedings of the International and multi-disciplinary Colloquim-Global warming, which potential impacts on the vineyards? Beaune, France, 28–30 March 2007; p. 23. [Google Scholar] [CrossRef]
- Hogendorp, B.K.; Cloyd, R.A.; Swiader, J.M. Effect of nitrogen fertility on reproduction and development of Citrus mealybug, Planococcus citri Risso (Homoptera: Pseudococcidae), feeding on two colors of coleus, Solenostemon scutellarioides L. Codd. Environ. Entomol. 2006, 35, 201–211. [Google Scholar] [CrossRef]
- Reineke, A.; Selim, M. Elevated atmospheric CO2 concentrations alter grapevine (Vitis vinifera) systemic transcriptional response to European grapevine moth (Lobesia botrana) herbivory. Sci. Rep. 2019, 9, 2995. [Google Scholar] [CrossRef] [PubMed]
- Ross, L.; Dealey, E.J.; Beukeboom, L.W.; Shuker, D.M. Temperature, age of mating and starvation determine the role of maternal effects on sex allocation in the mealybug Planococcus citri. Behav. Ecol. Sociobiol. 2011, 65, 909–919. [Google Scholar] [CrossRef] [PubMed]
- Cocco, A.; Marras, P.M.; Muscas, E.; Mura, A.; Lentini, A. Variation of life-history parameters of Planococcus ficus (Hemiptera: Pseudococcidae) in response to grapevine nitrogen fertilization. J. Appl. Entomol. 2015, 139, 519–528. [Google Scholar] [CrossRef]
- Bindi, M.; Fibbi, L.; Lanini, M.; Miglietta, F. Free air CO2 enrichment (FACE) of grapevine (Vitis vinifera L.): I. Development and testing of the system for CO2 enrichment. Eur. J. Agron. 2001, 14, 135–143. [Google Scholar] [CrossRef]
- Ryan, G.D.; Shukla, K.; Rasmussen, S.; Shelp, B.J.; Newman, J.A. Phloem phytochemistry and aphid responses to elevated CO2, nitrogen fertilization and endophyte infection. Agric. For. Entomol. 2014, 16, 273–283. [Google Scholar] [CrossRef]
- Zvereva, E.L.; Kozlov, M.V. Consequences of simultaneous elevation of carbon dioxide and temperature for plant-herbivore interactions: A metaanalysis. Glob. Chang. Biol. 2006, 12, 27–41. [Google Scholar] [CrossRef]
- Hughes, L.; Bazzaz, F.A. Effects of elevated CO2 on five plant-aphid interactions. Entomol. Exp. Appl. 2001, 99, 87–96. [Google Scholar] [CrossRef]
- Awmack, C.; Harrington, R.; Leather, S. Host plant effects on the performance of the aphid Aulacorthum solani (Kalt.) (Homoptera: Aphididae) at ambient and elevated CO2. Glob. Chang. Biol. 1997, 3, 545–549. [Google Scholar] [CrossRef]
- Poorter, H. Interspecific variation in the growth response of plants to an elevated ambient CO2 concentration. In CO2 and Biosphere; Springer: Dordrecht, The Netherlands, 1993; pp. 77–98. [Google Scholar]
- Bezemer, T.M.; Knight, K.J.; Newington, J.E.; Jones, T.H. How general are aphid responses to elevated atmospheric CO2? Ann. Entomol. Soc. Am. 1999, 92, 724–730. [Google Scholar] [CrossRef]
- Ryan, G.D.; Sylvester, E.V.A.; Shelp, B.J.; Newman, J.A. Towards an understanding of how phloem amino acid composition shapes elevated CO2-induced changes in aphid population dynamics. Ecol. Entomol. 2015, 40, 247–257. [Google Scholar] [CrossRef]
- Klaiber, J.; Najar-Rodriguez, A.J.; Dialer, E.; Dorn, S. Elevated carbon dioxide impairs the performance of a specialized parasitoid of an aphid host feeding on Brassica plants. Biol. Control 2013, 66, 49–55. [Google Scholar] [CrossRef]
- Jun Chen, F.; Wu, G.; Parajulee, M.N.; Ge, F. Impact of elevated CO2 on the third trophic level: A predator Harmonia axyridis and a parasitoid Aphidius picipes. Biocontrol Sci. Technol. 2007, 17, 313–324. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Site | Treatment | Parasitism Success (%) | Development Time (days) | Sex Ratio (% Females; % Males) |
|---|---|---|---|---|
| Greenhouse | aCO2 | 15 ± 17, n = 20 | 23.86 ± 1.35, n = 14 | 69.23; 30.77, n = 13 |
| eCO2 | 16 ± 22.1, n = 20 | 23.69 ± 0.79, n = 16 | 80; 20, n = 15 | |
| p | 0.87 | 0.67 | 0.67 | |
| VineyardFACE | aCO2 | 34.67 ± 34.01, n = 30 | 25.67 ± 3.34, n = 44 | 61.36; 38.63, n = 44 |
| eCO2 | 29.33 ± 27.66, n = 30 | 25.13 ± 2.54, n = 36 | 58.33; 41.67, n = 36 | |
| p | 0.51 | 0.41 | 0.82 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schulze-Sylvester, M.; Reineke, A. Elevated CO2 Levels Impact Fitness Traits of Vine Mealybug Planococcus ficus Signoret, but Not Its Parasitoid Leptomastix dactylopii Howard. Agronomy 2019, 9, 326. https://doi.org/10.3390/agronomy9060326
Schulze-Sylvester M, Reineke A. Elevated CO2 Levels Impact Fitness Traits of Vine Mealybug Planococcus ficus Signoret, but Not Its Parasitoid Leptomastix dactylopii Howard. Agronomy. 2019; 9(6):326. https://doi.org/10.3390/agronomy9060326
Chicago/Turabian StyleSchulze-Sylvester, Maria, and Annette Reineke. 2019. "Elevated CO2 Levels Impact Fitness Traits of Vine Mealybug Planococcus ficus Signoret, but Not Its Parasitoid Leptomastix dactylopii Howard" Agronomy 9, no. 6: 326. https://doi.org/10.3390/agronomy9060326
APA StyleSchulze-Sylvester, M., & Reineke, A. (2019). Elevated CO2 Levels Impact Fitness Traits of Vine Mealybug Planococcus ficus Signoret, but Not Its Parasitoid Leptomastix dactylopii Howard. Agronomy, 9(6), 326. https://doi.org/10.3390/agronomy9060326