
Pluripotent Stem Cells for Spinal Cord Injury Repair


Abstract
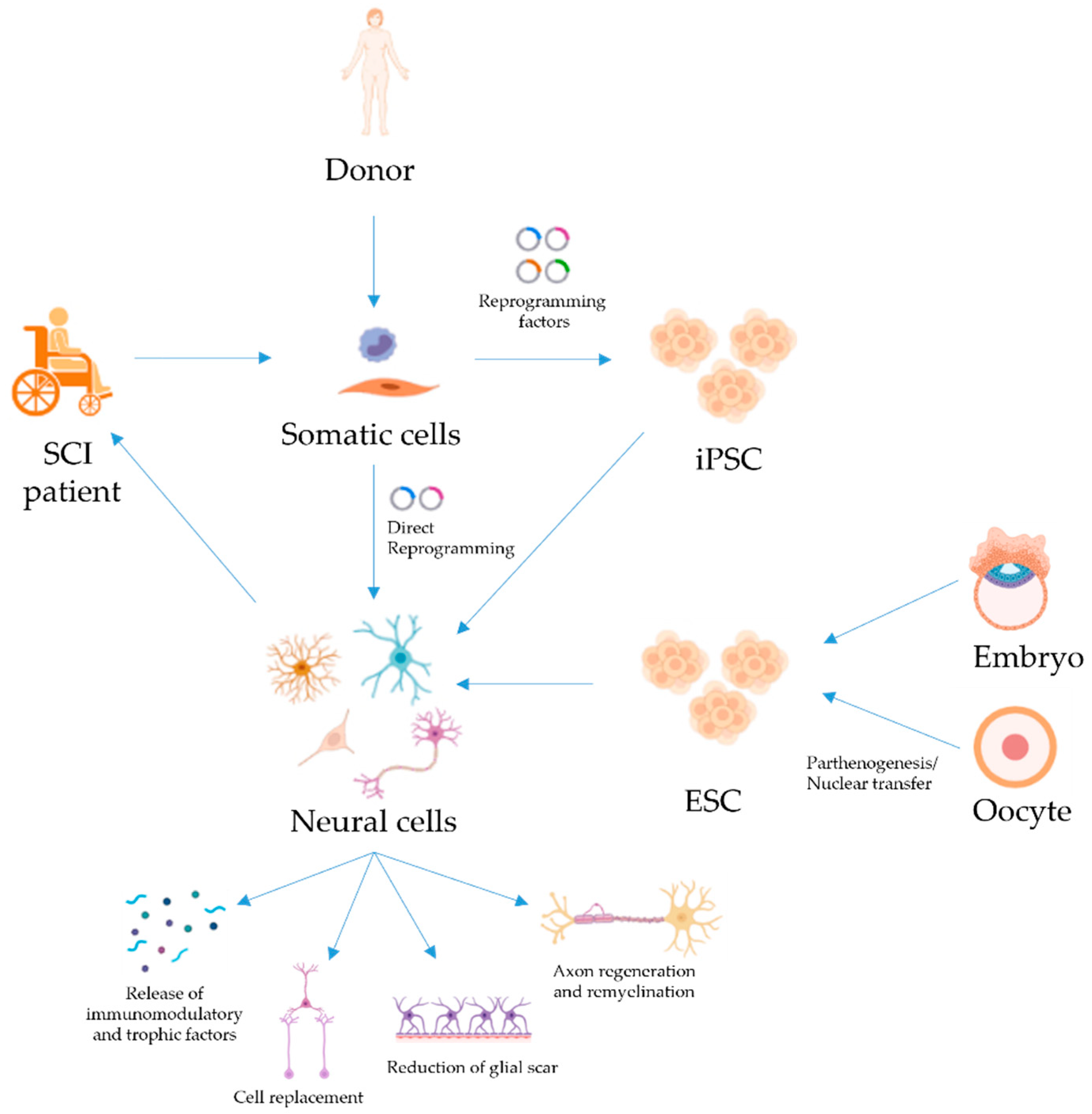
:1. Introduction
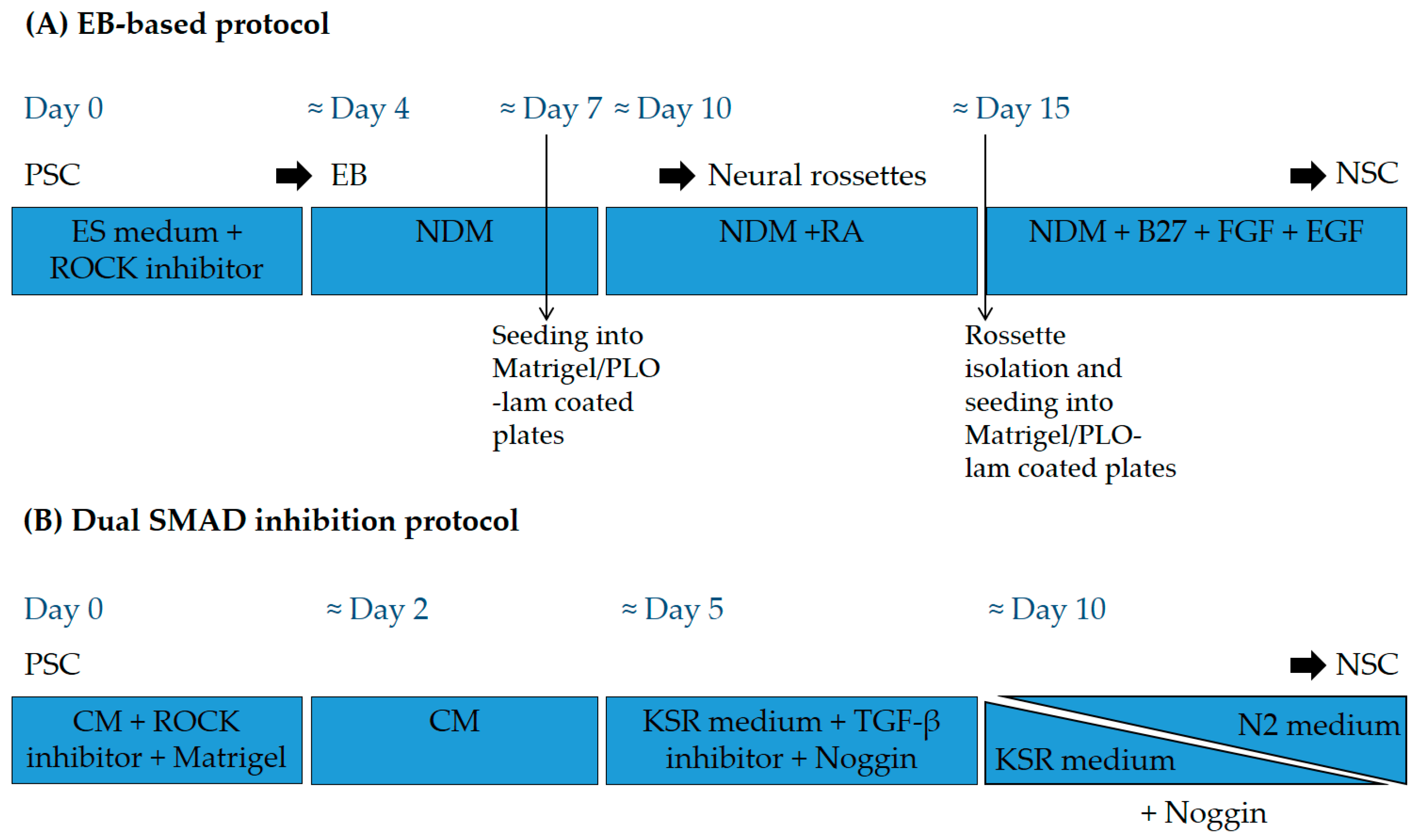
2. Embryonic Stem Cells for SCI Repair

{kind=link}
{kind=link}
| SCI Phase | Type of SCI | Animal Model | Level of Injury | Injected Cells | Number of Cells | Application Route | Timing of Transplantation | Tests Used for the Assessment of Recovery | Outcome | Year | Reference |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Acute | Contusion | Rat | Thoracic | Mouse RA-differentiated ESC–NSC | 1 × 106 | At the lesion epicenter | 9 days PI | BBB | Locomotor recovery | 1999 | [15] |
| Contusion | Rat | Thoracic | Human ESC–OPC | 2.5 × 105 or 1.5 × 106 | Rostral and caudal to the lesion epicenter | 7 days PI | BBB and 4 parameter kinematic analyses | Locomotor recovery | 2005 | [23] | |
| Contusion | Mouse | Thoracic | Mouse ESC–NSC | 2 × 104 | At the lesion epicenter | 10 days PI | Motor score, platform hang and rope walk | Locomotor recovery | 2005 | [16] | |
| Contusion | Rat | Thoracic | Human ESC–OPC | 1.5 × 106 | Rostral and caudal to the lesion epicenter | 7 days PI | BBB | Transplantation per se did not decrease locomotor function | 2006 | [24] # | |
| Contusion | Mouse | Thoracic | Mouse ESC–primary and secondary neurospheres | 5 × 105 | At the lesion epicenter | 9 days PI | BMS | Locomotor recovery | 2009 | [17] * | |
| Compression | Rat | Thoracic | Mouse ESC-motorneurons + OEC | 1 × 106 | At the lesion epicenter | 9 days PI | BBB | Locomotor recovery | 2009 | [36] | |
| Contusion | Rat | Cervical | Human ESC–OPC | 1.5 × 106 | Rostral and caudal to the lesion epicenter | 7 days PI | Forelimb movement scores | Locomotor recovery | 2010 | [30] | |
| Compression | Mouse | Thoracic | Mouse ESC–neural aggregates overexpressing L1 | 2 × 105 | Rostral and caudal to the lesion epicenter | 3 days PI | BBB score, foot-stepping angle and rump-height index | Locomotor recovery | 2011 | [20] | |
| Contusion | Rat | Thoracic | Human ESC–NSC + Schwann cells | 5 × 105 (NSC or Schwann cells) or 1 × 106 (NSC + Schwann cells) | At the lesion epicenter | 7 days PI | BBB | Locomotor recovery | 2012 | [35] | |
| Transection | Rat | Thoracic | Human ESC–NSC in fibrin with a growth factor cocktail | 2 × 106 | At the lesion epicenter | 7 days PI | BBB, 21-point locomotion rating scale, electrophysiological assessment | Locomotor recovery | 2012 | [18] | |
| Contusion | Rat; mouse | Thoracic | Human ESC–OPC | Rats: 2.4 × 105 or 2.4 × 106. Mice: 2.5 × 105 to 1 × 106 | Rats: 4 injections at the perimeter of the lesion or 1 injection rostral to the lesion epicenter. Mice: rostral to the lesion epicenter. | 6–8 days PI | None | Locomotor recovery in previous studies (see [23,24]) | 2015 | [25] | |
| Contusion | Rat | Thoracic | NG2 and MMP9 positive mouse ESC–NSC | 1 × 106 | At the lesion epicenter | 9 days PI | None | Axonal outgrowth into white matter | 2015 | [32] | |
| Contusion | Marmoset | Cervical | OPC-enriched marmoset ESC–NSC | 1 × 106 | At the lesion epicenter | 14 days PI | Open field and bar grip strength test | Locomotor recovery | 2015 | [31] * | |
| Contusion | Mouse | Thoracic | Human ESC-derived inhibitory interneuron precursors | 3 × 105 or 6–8 × 105 | Caudal to the injury epicenter | 15 days PI | BMS, Allodynia, Thermal hyperalgesia and bladder functional tests | Absence of locomotor recovery | 2016 | [34] | |
| Contusion | Mouse | Thoracic | Mouse ESC–NSC overexpressing BDNF | 1 × 105 | At the lesion epicenter | 7 days PI | BMS | Locomotor recovery | 2016 | [19] | |
| Contusion | Rat | Thoracic | Mouse ESC–NSC | 1 × 106 | At the lesion epicenter | 21 days PI | BBB and CBS | Absence of locomotor recovery | 2016 | [33] | |
| Contusion | Mouse | Thoracic | Murine iPSC and ESC–primary and secondary neurospheres | 5 × 105 | At the lesion epicenter | 9 days PI | BMS | Locomotor recovery | 2010 | [40] * | |
| Contusion | Rat | Thoracic | Mouse iPSC–derived astrocytes | 1 × 105 | At the lesion epicenter | 3–7 days PI | BBB, inclined-plane test, SCANET MV-40, sensory tests | Absence of locomotor recovery | 2011 | [41] | |
| Contusion | Marmoset | Cervical | Human iPSC–secondary and tertiary neurospheres | 1 × 106 | At the lesion epicenter | 9 days PI | Open field, bar grip, and cage climbing tests. | Locomotor recovery | 2012 | [42] * | |
| Contusion | Mouse | Thoracic | Human iPSC–NSC | 1 × 106 | At the lesion epicenter | 7 days PI | BMS, MEPs | Locomotor recovery | 2012 | [43] | |
| Hemisection | Rat | Cervical | Human iPSC–NSC in a fibrin matrix and growth factor cocktail | 1.25 × 106 | Three pairs of injections 0.5 mm apart, at the center, rostral, and caudal to the center of the lesion cavity | 14 days PI | Grid-walking, forelimb grooming and LUAT | Absence of locomotor recovery | 2014 | [44] | |
| Compression | Rat | Thoracic | Human iPSC–NSC | 5 × 105 | At the lesion epicenter | 7 days PI | BBB, plantar test, beam walking test, and RotaRod | Locomotor recovery | 2015 | [45] | |
| Compression | Mouse | Thoracic | Mouse iPSC–NSC | 2 × 105 | Four injections flanking the injury | 7 days PI | BMS, CatWalk, mechanical and thermal allodynia tests. | Locomotor recovery | 2015 | [46] | |
| Contusion | Mouse | Thoracic | Murine iPSC-derived neurospheres | 5 × 105 | At the lesion epicenter | 9 days PI | BMS, RotaRod and DigiGait | Locomotor recovery | 2011, 2015 | [47,48] * | |
| Contusion | Mouse | Thoracic | Human iPSC–NSC | 4 × 105 | Rostral and caudal to the lesion epicenter | 7 days PI | BMS and Catwalk | Absence of locomotor recovery | 2015 | [49] | |
| Compression | Rat | Thoracic | Human iPSC–NSC | 5 × 105 | At the lesion epicenter | 7 days PI | BBB, plantar test, beam walking test and RotaRod | Locomotor recovery | 2015 | [50] | |
| Compression | Rat | Thoracic | Human iPSC-OPC in a hydrogel with RGD and PDGF-A | 8 × 105 | Rostral and caudal to the lesion epicenter | 7 days PI | BBB | Locomotor recovery | 2016 | [51] | |
| Contusion | Mouse | Thoracic | Human iPSC–OPC | 5 × 105 | At the lesion epicenter | 9 days PI | BMS, RotaRod and DigiGait | Locomotor recovery | 2016 | [52] * | |
| Contusion | Mouse | Thoracic | Human iPSC–NSC treated with γ-secretase inhibitor | 5 × 105 | At the lesion epicenter | 9 days PI | BMS | Locomotor recovery | 2016 | [53] * | |
| Contusion | Mouse | Thoracic | Human iPSC–NSC | 5 × 105 | At the lesion epicenter | 9 days PI | BMS | Locomotor recovery (declined when tumors formed) | 2017 | [54] * | |
| Compression | Rat | Thoracic | Human iPSC–NSC conditioned with EI-tPA | 1.5 × 106 | At the lesion epicenter | 7 days PI | BBB | Locomotor recovery | 2019 | [55] | |
| Contusion | Mouse | Thoracic | Human iPSC–spinal cord–NSC | 5 × 105 | At the lesion epicenter | 9 days PI | BMS, RotaRod and treadmill analysis | Locomotor recovery | 2020 | [56] * | |
| Contusion | Mouse | Thoracic | Human iPSC–NSC | 1 × 105 | Rostral to the lesion epicenter | 7 days PI | BMS | Locomotor recovery | 2021 | [57] | |
| Contusion | Rat | Thoracic | Human iPSC–NSC + MSC + PA-C | 1.8 × 106 | Rostral, caudal, and at the lesion epicenter | 7 days PI | BBB and Catwalk | Absence of locomotor recovery | 2021 | [58] | |
| Chronic | Contusion | Rat | Thoracic | Human ESC–OPC | 2.5 × 105 or 1.5 × 106 | Rostral and caudal to the lesion epicenter | 10 months PI | BBB and four-kinematic analyses | Absence of locomotor recovery | 2005 | [23] |
| Contusion | Rat | Cervical | Human iPSC–NSC | 2 × 105 | Rostral and caudal to the lesion epicenter | 30 days PI | LUAT, FRT, allodynia test | Absence of locomotor recovery | 2013 | [59] | |
| Contusion | Mouse | Thoracic | Human iPSC–NSC treated with γ-secretase inhibitor | 5 × 105 | At the lesion epicenter | 42 days PI | BMS, RotaRod and treadmill analysis | Locomotor recovery | 2018 | [60] * | |
| Compression | Rat | Thoracic | Human iPSC–NSC on Laminin-Coated pHEMA-MOETACl Hydrogel | 3 × 105 | At the lesion epicenter | 35 days PI | BBB, plantar test | Absence of locomotor recovery | 2019 | [61] | |
| Accidental SCI | Dog | Thoracic | Canine iPSC–NSC | 2 × 106 | At the lesion epicenter, and one vertebral space caudal and rostral to the lesion | >28 days PI | Neurological and electrophysiological evaluation | Absence of locomotor recovery | 2020 | [62] | |
| Contusion | Rat | Thoracic | Glial scar photo-ablation + iPSC–regionally specific spinal pre-OPC | 5 × 105 | Rostral and caudal to the lesion epicenter | 70 days post-injury | BBB | Absence of locomotor recovery | 2021 | [63] | |
| Contusion | Rat | Cervical | Human iPSC–NSC | 4 × 105 | Rostral and caudal to the lesion epicenter | 28 days PI | FRT, IBB, and LUAT | Absence of locomotor recovery | 2021 | [64] |
3. iPSC for SCI Repair
| Type of SCI | Clinical Trial Dentifier | PSC | Final Cell Type | References |
|---|---|---|---|---|
| Complete subacute SCI, ASIA Impairment Scale A. Last fully preserved neurological level from T-3 through T-11 (7 to 14 days following SCI) | NCT01217008 (ClinicalTrials.gov) | ESC | OPC (AST-OPC1) | [25] |
| Subacute cervical SCI, ASIA Impairment Scale A and B. Last fully preserved single neurological level from C-4 to C-7 (21 to 42 days following SCI) | NCT 02302157 (ClinicalTrials.gov) | ESC | OPC (AST-OPC1) | [25] |
| Complete subacute cervical (C4-C7) SCI, ASIA Impairment Scale A. (7 to 60 days following SCI) | NCT04812431 (ClinicalTrials.gov) | ESC | NSC (PSA-NCAM(+)) | Not found |
| Complete subacute SCI (C3/4-Th10), ASIA Impairment Scale A (within 24 days following SCI) | jRCTa031190228 (Japan Registry of Clinical Trials) | Integration-free episomal iPSC from PBMC | NSC (to dopaminergic neuron fate) | [70] |
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ahuja, C.S.; Wilson, J.R.; Nori, S.; Kotter, M.R.N.; Druschel, C.; Curt, A.; Fehlings, M.G. Traumatic spinal cord injury. Nat. Rev. Dis. Prim. 2017, 3, 17018. [Google Scholar] [CrossRef] [PubMed]
- Silva, N.A.; Sousa, N.; Reis, R.L.; Salgado, A.J. From basics to clinical: A comprehensive review on spinal cord injury. Prog. Neurobiol. 2014, 114, 25–57. [Google Scholar] [CrossRef] [PubMed]
- Ronaghi, M.; Erceg, S.; Moreno-Manzano, V.; Stojkovic, M. Challenges of stem cell therapy for spinal cord injury: Human embryonic stem cells, endogenous neural stem cells, or induced pluripotent stem cells? Stem Cells 2010, 28, 93–99. [Google Scholar] [CrossRef]
- Takami, T.; Oudega, M.; Bates, M.L.; Wood, P.M.; Kleitman, N.; Bunge, M.B. Schwann Cell But Not Olfactory Ensheathing Glia Transplants Improve Hindlimb Locomotor Performance in the Moderately Contused Adult Rat Thoracic Spinal Cord. J. Neurosci. 2002, 22, 6670–6681. [Google Scholar] [CrossRef] [Green Version]
- Levi, A.D.O.; Dancausse, H.; Li, X.; Duncan, S.; Horkey, L.; Oliviera, M. Peripheral nerve grafts promoting central nervous system regeneration after spinal cord injury in the primate. J. Neurosurg. Spine 2002, 96, 197–205. [Google Scholar] [CrossRef]
- Mitsui, T.; Fischer, I.; Shumsky, J.S.; Murray, M. Transplants of fibroblasts expressing BDNF and NT-3 promote recovery of bladder and hindlimb function following spinal contusion injury in rats. Exp. Neurol. 2005, 194, 410–431. [Google Scholar] [CrossRef] [PubMed]
- Yao, R.; Murtaza, M.; Velasquez, J.T.; Todorovic, M.; Rayfield, A.; Ekberg, J.; Barton, M.; St John, J. Olfactory Ensheathing Cells for Spinal Cord Injury: Sniffing Out the Issues. Cell Transplant. 2018, 27, 879–889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tetzlaff, W.; Okon, E.B.; Karimi-Abdolrezaee, S.; Hill, C.E.; Sparling, J.S.; Plemel, J.R.; Plunet, W.T.; Tsai, E.C.; Baptiste, D.; Smithson, L.J.; et al. A Systematic Review of Cellular Transplantation Therapies for Spinal Cord Injury. J. Neurotrauma 2011, 28, 1611–1682. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Lepski, G. Cell transplantation for spinal cord injury: A systematic review. Biomed Res. Int. 2013, 2013, 786475. [Google Scholar] [CrossRef] [Green Version]
- Cofano, F.; Boido, M.; Monticelli, M.; Zenga, F.; Ducati, A.; Vercelli, A.; Garbossa, D. Mesenchymal Stem Cells for Spinal Cord Injury: Current Options, Limitations, and Future of Cell Therapy. Int. J. Mol. Sci. 2019, 20, 2698. [Google Scholar] [CrossRef] [Green Version]
- Zou, Y.; Zhao, Y.; Xiao, Z.; Chen, B.; Ma, D.; Shen, H.; Gu, R.; Dai, J. Comparison of Regenerative Effects of Transplanting Three-Dimensional Longitudinal Scaffold Loaded-Human Mesenchymal Stem Cells and Human Neural Stem Cells on Spinal Cord Completely Transected Rats. ACS Biomater. Sci. Eng. 2020, 6, 1671–1680. [Google Scholar] [CrossRef] [PubMed]
- Yousefifard, M.; Rahimi-Movaghar, V.; Nasirinezhad, F.; Baikpour, M.; Safari, S.; Saadat, S.; Moghadas Jafari, A.; Asady, H.; Razavi Tousi, S.M.T.; Hosseini, M. Neural stem/progenitor cell transplantation for spinal cord injury treatment; A systematic review and meta-analysis. Neuroscience 2016, 322, 377–397. [Google Scholar] [CrossRef] [Green Version]
- Fernandez-Muñoz, B.; Garcia-Delgado, A.B.; Arribas-Arribas, B.; Sanchez-Pernaute, R. Human Neural Stem Cells for Cell-Based Medicinal Products. Cells 2021, 10, 2377. [Google Scholar] [CrossRef]
- Lee, A.S.; Tang, C.; Rao, M.S.; Weissman, I.L.; Wu, J.C. Tumorigenicity as a clinical hurdle for pluripotent stem cell therapies. Nat. Med. 2013, 19, 998–1004. [Google Scholar] [CrossRef] [Green Version]
- Mcdonald, J.W.; Liu, X.Z.; Qu, Y.; Liu, S.; Mickey, S.K.; Turetsky, D.; Gottlieb, D.I.; Choi, D.W. Transplanted embryonic stem cells survive, differentiate and promote recovery in injured rat spinal cord. Nat. Med. 1999, 5, 1410–1412. [Google Scholar] [CrossRef] [PubMed]
- Kimura, H.; Yoshikawa, M.; Matsuda, R.; Toriumi, H.; Nishimura, F.; Hirabayashi, H.; Nakase, H.; Kawaguchi, S.; Ishizaka, S.; Sakaki, T. Transplantation of embryonic stem cell-derived neural stem cells for spinal cord injury in adult mice. Neurol. Res. 2005, 27, 812–819. [Google Scholar] [CrossRef]
- Kumagai, G.; Okada, Y.; Yamane, J.; Nagoshi, N.; Kitamura, K.; Mukaino, M.; Tsuji, O.; Fujiyoshi, K.; Katoh, H.; Okada, S.; et al. Roles of ES Cell-Derived Gliogenic Neural Stem/Progenitor Cells in Functional Recovery after Spinal Cord Injury. PLoS ONE 2009, 4, e7706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, P.; Wang, Y.; Graham, L.; McHale, K.; Gao, M.; Wu, D.; Brock, J.; Blesch, A.; Rosenzweig, E.S.; Havton, L.A.; et al. Long-distance growth and connectivity of neural stem cells after severe spinal cord injury. Cell 2012, 150, 1264–1273. [Google Scholar] [CrossRef] [Green Version]
- Butenschön, J.; Zimmermann, T.; Schmarowski, N.; Nitsch, R.; Fackelmeier, B.; Friedemann, K.; Radyushkin, K.; Baumgart, J.; Lutz, B.; Leschik, J. PSA-NCAM positive neural progenitors stably expressing BDNF promote functional recovery in a mouse model of spinal cord injury. Stem Cell Res. Ther. 2016, 7, 11. [Google Scholar] [CrossRef] [Green Version]
- Cui, Y.F.; Xu, J.C.; Hargus, G.; Jakovcevski, I.; Schachner, M.; Bernreuther, C. Embryonic stem cell-derived L1 overexpressing neural aggregates enhance recovery after spinal cord injury in mice. PLoS ONE 2011, 6, 17126. [Google Scholar] [CrossRef] [Green Version]
- Steward, O.; Sharp, K.G.; Matsudaira Yee, K. Long-Distance Migration and Colonization of Transplanted Neural Stem Cells. Cell 2014, 156, 385–387. [Google Scholar] [CrossRef] [Green Version]
- Tuszynski, M.H.; Wang, Y.; Graham, L.; Gao, M.; Wu, D.; Brock, J.; Blesch, A.; Rosenzweig, E.S.; Havton, L.A.; Zheng, B.; et al. Neural stem cell dissemination after grafting to CNS injury sites. Cell 2014, 156, 388–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keirstead, H.S.; Nistor, G.; Bernal, G.; Totoiu, M.; Cloutier, F.; Sharp, K.; Steward, O. Human Embryonic Stem Cell-Derived Oligodendrocyte Progenitor Cell Transplants Remyelinate and Restore Locomotion after Spinal Cord Injury. J. Neurosci. 2005, 25, 4694–4705. [Google Scholar] [CrossRef]
- Cloutier, F.; Siegenthaler, M.M.; Nistor, G.; Keirstead, H.S. Transplantation of human embryonic stem cell-derived oligodendrocyte progenitors into rat spinal cord injuries does not cause harm. Regen. Med. 2006, 1, 469–479. [Google Scholar] [CrossRef] [Green Version]
- Priest, C.A.; Manley, N.C.; Denham, J.; Wirth, E.D.; Lebkowski, J.S. Preclinical safety of human embryonic stem cell-derived oligodendrocyte progenitors supporting clinical trials in spinal cord injury. Regen. Med. 2015, 10, 939–958. [Google Scholar] [CrossRef] [Green Version]
- Hayden, E.C. Funding windfall rescues abandoned stem-cell trial. Nature 2014, 510, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manley, N.C.; Priest, C.A.; Denham, J.; Wirth, E.D., III; Lebkowski, J.S. Human Embryonic Stem Cell-Derived Oligodendrocyte Progenitor Cells: Preclinical Efficacy and Safety in Cervical Spinal Cord Injury. Stem Cells Transl. Med. 2017, 6, 1917. [Google Scholar] [CrossRef] [PubMed]
- Asterias Provides Top Line 12 Month Data Update for Its OPC1 Phase 1/2a Clinical Trial in Severe Spinal Cord Injury. Available online: https://www.globenewswire.com/news-release/2019/01/24/1704757/0/en/Asterias-Provides-Top-Line-12-Month-Data-Update-for-its-OPC1-Phase-1-2a-Clinical-Trial-in-Severe-Spinal-Cord-Injury.html (accessed on 8 October 2021).
- Corporate Overview the Future of Cell Therapy. Available online: www.sec.gov (accessed on 13 October 2021).
- Sharp, J.; Frame, J.; Siegenthaler, M.; Nistor, G.; Keirstead, H.S. Human embryonic stem cell-derived oligodendrocyte progenitor cell transplants improve recovery after cervical spinal cord injury. Stem Cells 2010, 28, 152–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwai, H.; Shimada, H.; Nishimura, S.; Kobayashi, Y.; Itakura, G.; Hori, K.; Hikishima, K.; Ebise, H.; Negishi, N.; Shibata, S.; et al. Allogeneic Neural Stem/Progenitor Cells Derived from Embryonic Stem Cells Promote Functional Recovery after Transplantation into Injured Spinal Cord of Nonhuman Primates. Stem Cells Transl. Med. 2015, 4, 708–719. [Google Scholar] [CrossRef] [Green Version]
- Vadivelu, S.; Stewart, T.J.; Qu, Y.; Horn, K.; Liu, S.; Li, Q.; Silver, J.; McDonald, J.W. NG2+ Progenitors Derived from Embryonic Stem Cells Penetrate Glial Scar and Promote Axonal Outgrowth into White Matter after Spinal Cord Injury. Stem Cells Transl. Med. 2015, 4, 401–411. [Google Scholar] [CrossRef] [Green Version]
- Hwang, I.; Hahm, S.-C.; Choi, K.-A.; Park, S.-H.; Jeong, H.; Yea, J.-H.; Kim, J.; Hong, S. Intrathecal Transplantation of Embryonic Stem Cell-Derived Spinal GABAergic Neural Precursor Cells Attenuates Neuropathic Pain in a Spinal Cord Injury Rat Model. Cell Transplant. 2016, 25, 593–607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fandel, T.M.; Trivedi, A.; Nicholas, C.R.; Zhang, H.; Chen, J.; Martinez, A.F.; Noble-Haeusslein, L.J.; Kriegstein, A.R. Transplanted Human Stem Cell-Derived Interneuron Precursors Mitigate Mouse Bladder Dysfunction and Central Neuropathic Pain after Spinal Cord Injury. Cell Stem Cell 2016, 19, 544–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niapour, A.; Karamali, F.; Nemati, S.; Taghipour, Z.; Mardaniz, M.; Nasr-Esfahani, M.H.; Baharvand, H. Cotransplantation of human embryonic stem cell-derived neural progenitors and Schwann cells in a rat spinal cord contusion injury model elicits a distinct neurogenesis and functional recovery. Cell Transplant. 2012, 21, 827–843. [Google Scholar] [CrossRef] [Green Version]
- Salehi, M.; Pasbakhsh, P.; Soleimani, M.; Abbasi, M.; Hasanzadeh, G.; Modaresi, M.H.; Sobhani, A. Repair of spinal cord injury by co-transplantation of embryonic stem cell-derived motor neuron and olfactory ensheathing cell. Iran. Biomed. J. 2009, 13, 125–135. [Google Scholar] [PubMed]
- Vanický, I.; Urdzíková, L.; Saganová, K.; Čízková, D.; Gálik, J. A Simple and Reproducible Model of Spinal Cord Injury Induced by Epidural Balloon Inflation in the Rat. J. Neurotrauma 2001, 18, 1399–1407. [Google Scholar] [CrossRef] [PubMed]
- Lo, B.; Parham, L. Ethical Issues in Stem Cell Research. Endocr. Rev. 2009, 30, 204. [Google Scholar] [CrossRef]
- Kobold, S.; Guhr, A.; Mah, N.; Bultjer, N.; Seltmann, S.; Seiler Wulczyn, A.E.M.; Stacey, G.; Jie, H.; Liu, W.; Löser, P.; et al. A Manually Curated Database on Clinical Studies Involving Cell Products Derived from Human Pluripotent Stem Cells. Stem Cell Rep. 2020, 15, 546–555. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, O.; Miura, K.; Okada, Y.; Fujiyoshi, K.; Mukaino, M.; Nagoshi, N.; Kitamura, K.; Kumagai, G.; Nishino, M.; Tomisato, S.; et al. Therapeutic potential of appropriately evaluated safe-induced pluripotent stem cells for spinal cord injury. Proc. Natl. Acad. Sci. USA 2010, 107, 12704–12709. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, K.; Hashimoto, M.; Koda, M.; Naito, A.T.; Murata, A.; Okawa, A.; Takahashi, K.; Yamazaki, M. Increase of sensitivity to mechanical stimulus after transplantation of murine induced pluripotent stem cell–derived astrocytes in a rat spinal cord injury model. J. Neurosurg. Spine 2011, 15, 582–593. [Google Scholar] [CrossRef]
- Kobayashi, Y.; Okada, Y.; Itakura, G.; Iwai, H.; Nishimura, S.; Yasuda, A.; Nori, S.; Hikishima, K.; Konomi, T.; Fujiyoshi, K.; et al. Pre-Evaluated Safe Human iPSC-Derived Neural Stem Cells Promote Functional Recovery after Spinal Cord Injury in Common Marmoset without Tumorigenicity. PLoS ONE 2012, 7, e52787. [Google Scholar] [CrossRef] [Green Version]
- Fujimoto, Y.; Abematsu, M.; Falk, A.; Tsujimura, K.; Sanosaka, T.; Juliandi, B.; Semi, K.; Namihira, M.; Komiya, S.; Smith, A.; et al. Treatment of a mouse model of spinal cord injury by transplantation of human induced pluripotent stem cell-derived long-term self-renewing neuroepithelial-like stem cells. Stem Cells 2012, 30, 1163–1173. [Google Scholar] [CrossRef]
- Lu, P.; Woodruff, G.; Wang, Y.; Graham, L.; Hunt, M.; Wu, D.; Boehle, E.; Ahmad, R.; Poplawski, G.; Brock, J.; et al. Long-Distance Axonal Growth from Human Induced Pluripotent Stem Cells after Spinal Cord Injury. Neuron 2014, 83, 789–796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romanyuk, N.; Amemori, T.; Turnovcova, K.; Prochazka, P.; Onteniente, B.; Sykova, E.; Jendelova, P. Beneficial Effect of Human Induced Pluripotent Stem Cell-Derived Neural Precursors in Spinal Cord Injury Repair. Cell Transplant. 2015, 24, 1781–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salewski, R.P.; Mitchell, R.A.; Li, L.; Shen, C.; Milekovskaia, M.; Nagy, A.; Fehlings, M.G. Transplantation of Induced Pluripotent Stem Cell-Derived Neural Stem Cells Mediate Functional Recovery Following Thoracic Spinal Cord Injury Through Remyelination of Axons. Stem Cells Transl. Med. 2015, 4, 743–754. [Google Scholar] [CrossRef]
- Nori, S.; Okada, Y.; Yasuda, A.; Tsuji, O.; Takahashi, Y.; Kobayashi, Y.; Fujiyoshi, K.; Koike, M.; Uchiyama, Y.; Ikeda, E.; et al. Grafted human-induced pluripotent stem-cell-derived neurospheres promote motor functional recovery after spinal cord injury in mice. Proc. Natl. Acad. Sci. USA 2011, 108, 16825–16830. [Google Scholar] [CrossRef] [Green Version]
- Nori, S.; Okada, Y.; Nishimura, S.; Sasaki, T.; Itakura, G.; Kobayashi, Y.; Renault-Mihara, F.; Shimizu, A.; Koya, I.; Yoshida, R.; et al. Long-term safety issues of iPSC-based cell therapy in a spinal cord injury model: Oncogenic transformation with epithelial-mesenchymal transition. Stem Cell Rep. 2015, 4, 360–373. [Google Scholar] [CrossRef] [Green Version]
- Pomeshchik, Y.; Puttonen, K.A.; Kidin, I.; Ruponen, M.; Lehtonen, S.; Malm, T.; Åkesson, E.; Hovatta, O.; Koistinaho, J. Transplanted Human Induced Pluripotent Stem Cell-Derived Neural Progenitor Cells Do Not Promote Functional Recovery of Pharmacologically Immunosuppressed Mice with Contusion Spinal Cord Injury. Cell Transplant. 2015, 24, 1799–1812. [Google Scholar] [CrossRef] [Green Version]
- Amemori, T.; Ruzicka, J.; Romanyuk, N.; Jhanwar-Uniyal, M.; Sykova, E.; Jendelova, P. Comparison of intraspinal and intrathecal implantation of induced pluripotent stem cell-derived neural precursors for the treatment of spinal cord injury in rats. Stem Cell Res. Ther. 2015, 6, 257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Führmann, T.; Tam, R.Y.; Ballarin, B.; Coles, B.; Elliott Donaghue, I.; van der Kooy, D.; Nagy, A.; Tator, C.H.; Morshead, C.M.; Shoichet, M.S. Injectable hydrogel promotes early survival of induced pluripotent stem cell-derived oligodendrocytes and attenuates longterm teratoma formation in a spinal cord injury model. Biomaterials 2016, 83, 23–36. [Google Scholar] [CrossRef]
- Kawabata, S.; Takano, M.; Numasawa-Kuroiwa, Y.; Itakura, G.; Kobayashi, Y.; Nishiyama, Y.; Sugai, K.; Nishimura, S.; Iwai, H.; Isoda, M.; et al. Grafted Human iPS Cell-Derived Oligodendrocyte Precursor Cells Contribute to Robust Remyelination of Demyelinated Axons after Spinal Cord Injury. Stem Cell Rep. 2016, 6, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Okubo, T.; Iwanami, A.; Kohyama, J.; Itakura, G.; Kawabata, S.; Nishiyama, Y.; Sugai, K.; Ozaki, M.; Iida, T.; Matsubayashi, K.; et al. Pretreatment with a γ-Secretase Inhibitor Prevents Tumor-like Overgrowth in Human iPSC-Derived Transplants for Spinal Cord Injury. Stem Cell Rep. 2016, 7, 649–663. [Google Scholar] [CrossRef] [Green Version]
- Itakura, G.; Kawabata, S.; Ando, M.; Nishiyama, Y.; Sugai, K.; Ozaki, M.; Iida, T.; Ookubo, T.; Kojima, K.; Kashiwagi, R.; et al. Fail-Safe System against Potential Tumorigenicity after Transplantation of iPSC Derivatives. Stem Cell Rep. 2017, 8, 673–684. [Google Scholar] [CrossRef] [PubMed]
- Shiga, Y.; Shiga, A.; Mesci, P.; Kwon, H.; Brifault, C.; Kim, J.H.; Jeziorski, J.J.; Nasamran, C.; Ohtori, S.; Muotri, A.R.; et al. Tissue-type plasminogen activator-primed human iPSC-derived neural progenitor cells promote motor recovery after severe spinal cord injury. Sci. Rep. 2019, 9, 19291. [Google Scholar] [CrossRef] [PubMed]
- Kajikawa, K.; Imaizumi, K.; Shinozaki, M.; Shibata, S.; Shindo, T.; Kitagawa, T.; Shibata, R.; Kamata, Y.; Kojima, K.; Nagoshi, N.; et al. Cell therapy for spinal cord injury by using human iPSC-derived region-specific neural progenitor cells. Mol. Brain 2020, 13, 120. [Google Scholar] [CrossRef] [PubMed]
- Kong, D.; Feng, B.; Amponsah, A.E.; He, J.; Guo, R.; Liu, B.; Du, X.; Liu, X.; Zhang, S.; Lv, F.; et al. hiPSC-derived NSCs effectively promote the functional recovery of acute spinal cord injury in mice. Stem Cell Res. Ther. 2021, 12, 172. [Google Scholar] [CrossRef] [PubMed]
- Bonilla, P.; Hernandez, J.; Giraldo, E.; González-Pérez, M.A.; Alastrue-Agudo, A.; Elkhenany, H.; Vicent, M.J.; Navarro, X.; Edel, M.; Moreno-Manzano, V. Human-induced neural and mesenchymal stem cell therapy combined with a curcumin nanoconjugate as a spinal cord injury treatment. Int. J. Mol. Sci. 2021, 22, 5966. [Google Scholar] [CrossRef]
- Nutt, S.E.; Chang, E.-A.; Suhr, S.T.; Schlosser, L.O.; Mondello, S.E.; Moritz, C.T.; Cibelli, J.B.; Horner, P.J. Caudalized human iPSC-derived neural progenitor cells produce neurons and glia but fail to restore function in an early chronic spinal cord injury model. Exp. Neurol. 2013, 248, 491–503. [Google Scholar] [CrossRef] [Green Version]
- Okubo, T.; Nagoshi, N.; Kohyama, J.; Tsuji, O.; Shinozaki, M.; Shibata, S.; Kase, Y.; Matsumoto, M.; Nakamura, M.; Okano, H. Treatment with a Gamma-Secretase Inhibitor Promotes Functional Recovery in Human iPSC- Derived Transplants for Chronic Spinal Cord Injury. Stem Cell Rep. 2018, 11, 1416–1432. [Google Scholar] [CrossRef] [Green Version]
- Ruzicka, J.; Romanyuk, N.; Jirakova, K.; Hejcl, A.; Janouskova, O.; Machova, L.U.; Bochin, M.; Pradny, M.; Vargova, L.; Jendelova, P. The Effect of iPS-Derived Neural Progenitors Seeded on Laminin-Coated pHEMA-MOETACl Hydrogel with Dual Porosity in a Rat Model of Chronic Spinal Cord Injury. Cell Transplant. 2019, 28, 400–412. [Google Scholar] [CrossRef] [Green Version]
- Chow, L.; McGrath, S.; de Arruda Saldanha, C.; Whalen, L.R.; Packer, R.; Dow, S. Generation of Neural Progenitor Cells from Canine Induced Pluripotent Stem Cells and Preliminary Safety Test in Dogs with Spontaneous Spinal Cord Injuries. Front. Vet. Sci. 2020, 7, 575938. [Google Scholar] [CrossRef]
- Patil, N.; Walsh, P.; Carrabre, K.; Holmberg, E.G.; Lavoie, N.; Dutton, J.R.; Parr, A.M. Regionally Specific Human Pre-Oligodendrocyte Progenitor Cells Produce Both Oligodendrocytes and Neurons after Transplantation in a Chronically Injured Spinal Cord Rat Model after Glial Scar Ablation. J. Neurotrauma 2021, 38, 777–788. [Google Scholar] [CrossRef] [PubMed]
- Martín-López, M.; González-Muñoz, E.; Gómez-González, E.; Sánchez-Pernaute, R.; Márquez-Rivas, J.; Fernández-Muñoz, B. Modeling chronic cervical spinal cord injury in aged rats for cell therapy studies. J. Clin. Neurosci. 2021, 94, 76–85. [Google Scholar] [CrossRef]
- Kjell, J.; Olson, L. Rat models of spinal cord injury: From pathology to potential therapies. Dis. Model. Mech. 2016, 9, 1125–1137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alaverdashvili, M.; Whishaw, I.Q. Compensation aids skilled reaching in aging and in recovery from forelimb motor cortex stroke in the rat. Neuroscience 2010, 167, 21–30. [Google Scholar] [CrossRef]
- Jain, N.B.; Ayers, G.D.; Peterson, E.N.; Harris, M.B.; Morse, L.; O’Connor, K.C.; Garshick, E. Traumatic Spinal Cord Injury in the United States, 1993-2012. JAMA 2015, 313, 2236. [Google Scholar] [CrossRef] [PubMed]
- Nishi, R.A.; Badner, A.; Hooshmand, M.J.; Creasman, D.A.; Liu, H.; Anderson, A.J. The effects of mouse strain and age on a model of unilateral cervical contusion spinal cord injury. PLoS ONE 2020, 15, e0234245. [Google Scholar] [CrossRef]
- Miura, K.; Okada, Y.; Aoi, T.; Okada, A.; Takahashi, K.; Okita, K.; Nakagawa, M.; Koyanagi, M.; Tanabe, K.; Ohnuki, M.; et al. Variation in the safety of induced pluripotent stem cell lines. Nat. Biotechnol. 2009, 27, 743–745. [Google Scholar] [CrossRef] [Green Version]
- Sugai, K.; Sumida, M.; Shofuda, T.; Yamaguchi, R.; Tamura, T.; Kohzuki, T.; Abe, T.; Shibata, R.; Kamata, Y.; Ito, S.; et al. First-in-human clinical trial of transplantation of iPSC-derived NS/PCs in subacute complete spinal cord injury: Study protocol. Regen. Ther. 2021, 18, 321–333. [Google Scholar] [CrossRef]
- Barker, R.A.; Studer, L.; Cattaneo, E.; Takahashi, J. G-Force PD: A global initiative in coordinating stem cell-based dopamine treatments for Parkinson’s disease. NPJ Park. Dis. 2015, 1, 15017. [Google Scholar] [CrossRef] [Green Version]
- Callahan, A.; Anderson, K.D.; Beattie, M.S.; Bixby, J.L.; Ferguson, A.R.; Fouad, K.; Jakeman, L.B.; Nielson, J.L.; Popovich, P.G.; Schwab, J.M.; et al. Developing a data sharing community for spinal cord injury research. Exp. Neurol. 2017, 295, 135–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheriyan, T.; Dj, R.; Weinreb, J.H.; Cheriyan, J.; Paul, J.C.; Lafage, V.; Kirsch, T.; Errico, T.J. Spinal cord injury models: A review. Spinal Cord 2014, 52, 588–595. [Google Scholar] [CrossRef]
- Winanto, Z.J.K.; Hor, J.H.; Ng, S.Y. Spinal cord organoids add an extra dimension to traditional motor neuron cultures. Neural Regen. Res. 2019, 14, 1515. [Google Scholar] [CrossRef]
- Andersen, J.; Revah, O.; Miura, Y.; Thom, N.; Amin, N.D.; Kelley, K.W.; Singh, M.; Chen, X.; Thete, M.V.; Walczak, E.M.; et al. Generation of Functional Human 3D Cortico-Motor Assembloids. Cell 2020, 183, 1913–1929.e26. [Google Scholar] [CrossRef]
- Tannenbaum, C.; Day, D. Age and sex in drug development and testing for adults. Pharmacol. Res. 2017, 121, 83–93. [Google Scholar] [CrossRef]
- Vrinten, D.H.; Hamers, F.F. ‘CatWalk’ automated quantitative gait analysis as a novel method to assess mechanical allodynia in the rat; a comparison with von Frey testing. Pain 2003, 102, 203–209. [Google Scholar] [CrossRef]
- Guo, J.; Zeng, Y.; Liang, Y.; Wang, L.; Su, H.; Wu, W. Cyclosporine affects the proliferation and differentiation of neural stem cells in culture. Neuroreport 2007, 18, 863–868. [Google Scholar] [CrossRef] [PubMed]
- Hunt, J.; Cheng, A.; Hoyles, A.; Jervis, E.; Morshead, C.M. Cyclosporin A Has Direct Effects on Adult Neural Precursor Cells. J. Neurosci. 2010, 30, 2888–2896. [Google Scholar] [CrossRef] [Green Version]
- Sontag, C.J.; Nguyen, H.X.; Kamei, N.; Uchida, N.; Anderson, A.J.; Cummings, B.J. Immunosuppressants Affect Human Neural Stem Cells In Vitro but Not in an In Vivo Model of Spinal Cord Injury. Stem Cells Transl. Med. 2013, 2, 731–744. [Google Scholar] [CrossRef] [PubMed]
- Sachewsky, N.; Hunt, J.; Cooke, M.J.; Azimi, A.; Zarin, T.; Miu, C.; Shoichet, M.S.; Morshead, C.M. Cyclosporin A enhances neural precursor cell survival in mice through a calcineurin-independent pathway. Dis. Model. Mech. 2014, 7, 953–961. [Google Scholar] [CrossRef] [Green Version]
- Matthews, K.R.; Moralí, D. National human embryo and embryoid research policies: A survey of 22 top research-intensive countries. Regen. Med. 2020, 15, 1905–1917. [Google Scholar] [CrossRef]
- Narsinh, K.H.; Plews, J.; Wu, J.C. Comparison of Human Induced Pluripotent and Embryonic Stem Cells: Fraternal or Identical Twins? Mol. Ther. 2011, 19, 635–638. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, S.; Fairchild, P.J.; Marsh, S.G.E.; Müller, C.R.; Turner, M.L.; Song, J.; Turner, D. Haplobanking induced pluripotent stem cells for clinical use. Stem Cell Res. 2020, 49, 102035. [Google Scholar] [CrossRef] [PubMed]
- Frederiksen, H.R.; Doehn, U.; Tveden-Nyborg, P.; Freude, K.K. Non-immunogenic Induced Pluripotent Stem Cells, a Promising Way Forward for Allogenic Transplantations for Neurological Disorders. Front. Genome Ed. 2021, 2, 30. [Google Scholar] [CrossRef] [PubMed]
- Ahuja, C.S.; Mothe, A.; Khazaei, M.; Badhiwala, J.H.; Gilbert, E.A.; Kooy, D.; Morshead, C.M.; Tator, C.; Fehlings, M.G. The leading edge: Emerging neuroprotective and neuroregenerative cell-based therapies for spinal cord injury. Stem Cells Transl. Med. 2020, 9, 1509–1530. [Google Scholar] [CrossRef] [PubMed]


| 1. PSC’ intrinsic tumorigenic potential |
| 2. Immunogenicity problems associated with the allogeneic use (immunosuppressants required) |
| 3. Optimal cell type, dose, route, timing and immunosuppression regime for each condition still not known |
| 4. Lack of efficacy in some conditions, especially in chronic SCI |
| 5. High production costs |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martin-Lopez, M.; Fernandez-Muñoz, B.; Canovas, S. Pluripotent Stem Cells for Spinal Cord Injury Repair. Cells 2021, 10, 3334. https://doi.org/10.3390/cells10123334
Martin-Lopez M, Fernandez-Muñoz B, Canovas S. Pluripotent Stem Cells for Spinal Cord Injury Repair. Cells. 2021; 10(12):3334. https://doi.org/10.3390/cells10123334
Chicago/Turabian StyleMartin-Lopez, Maria, Beatriz Fernandez-Muñoz, and Sebastian Canovas. 2021. "Pluripotent Stem Cells for Spinal Cord Injury Repair" Cells 10, no. 12: 3334. https://doi.org/10.3390/cells10123334
APA StyleMartin-Lopez, M., Fernandez-Muñoz, B., & Canovas, S. (2021). Pluripotent Stem Cells for Spinal Cord Injury Repair. Cells, 10(12), 3334. https://doi.org/10.3390/cells10123334