
Study of ALDH from Thermus thermophilus—Expression, Purification and Characterisation of the Non-Substrate Specific, Thermophilic Enzyme Displaying Both Dehydrogenase and Esterase Activity


Abstract
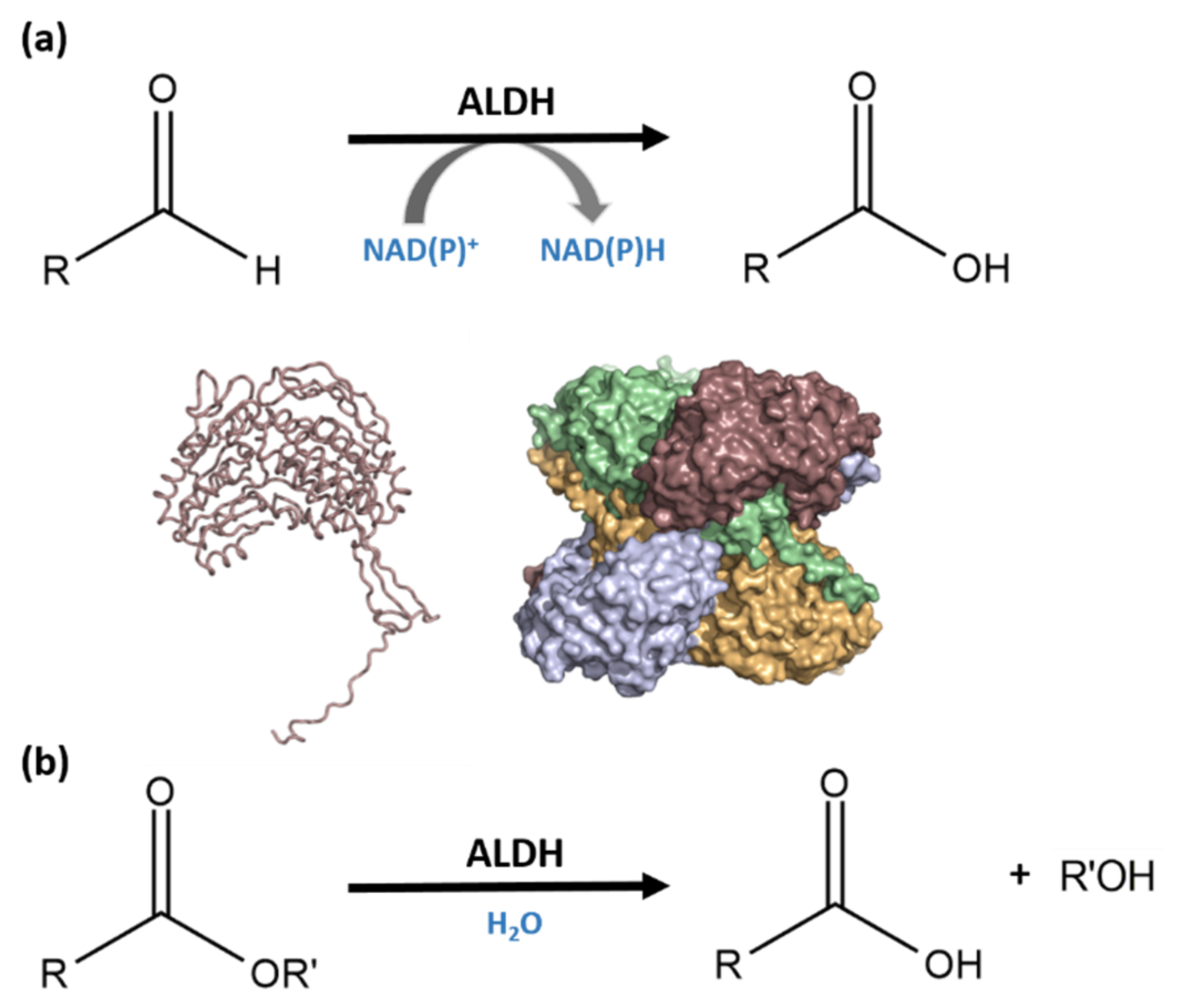
:1. Introduction
2. Materials and Methods
2.1. Protein Sequence Alignment and BLAST Analysis
2.2. Native ALDHTt Purification with caa3-Oxidase
2.3. ALDHTt Recombinant Expression and Purification
2.4. SDS-PAGE and Western Blot
2.5. MALDI-TOF Mass Spectrometry
2.6. Activity Assessment–Temperature and pH
2.7. Substrate Screening of Aldehydes
2.8. ALDHTt Reduction Reaction
2.9. Determination of Kinetic Parameters
2.10. HPLC Detection of Hexanoic Acid
2.11. Esterase Activity
3. Results
3.1. Protein Sequence Alignment
3.2. Recombinant ALDHTt Expression and Purification
3.3. Determination of ALDHTt Molecular Mass by MALDI-TOF
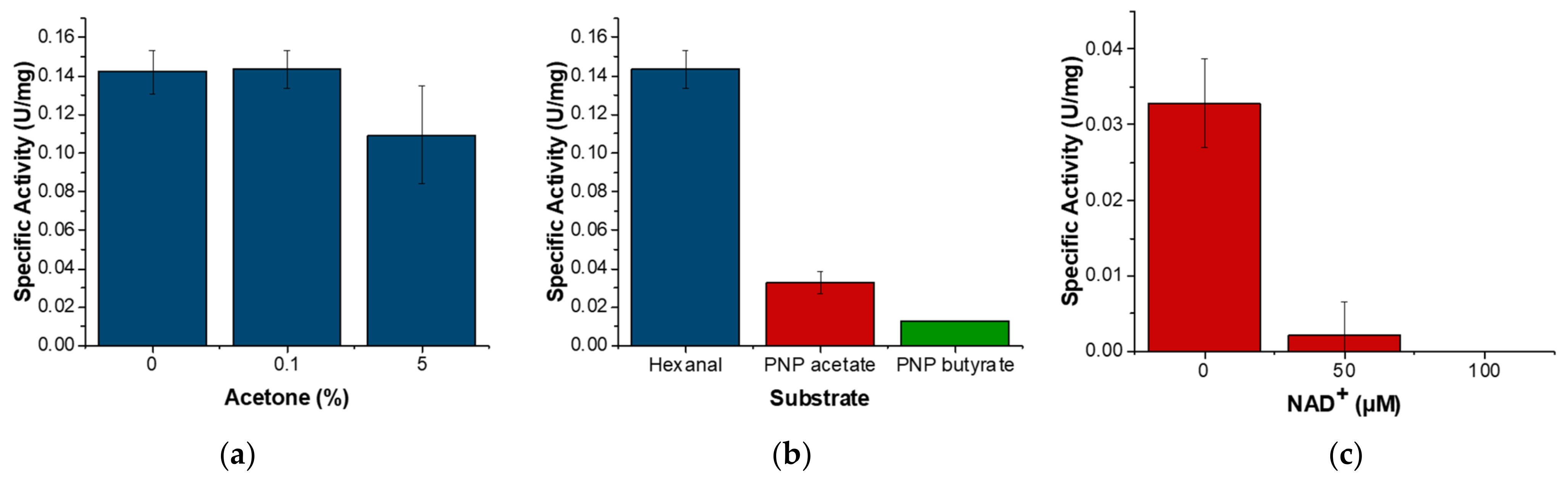
3.4. ALDHTt Enzymatic Activity Profiles
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, Z.-J.; Sun, Y.-J.; Rose, J.; Chung, Y.-J.; Hsiao, C.-D.; Chang, W.-R.; Kuo, I.; Perozich, J.; Lindahl, R.; Hempel, J. The first structure of an aldehyde dehydrogenase reveals novel interactions between NAD and the Rossmann fold. Nat. Struct. Biol. 1997, 4, 317–326. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Zavala, J.S.; Allali-Hassani, A.; Weiner, H. Characterization of E. coli tetrameric aldehyde dehydrogenases with atypical properties compared to other aldehyde dehydrogenases. Protein Sci. 2006, 15, 1387–1396. [Google Scholar] [CrossRef] [Green Version]
- Korasick, D.A.; White, T.A.; Chakravarthy, S.; Tanner, J.J. NAD+ promotes assembly of the active tetramer of aldehyde dehydrogenase 7A1. FEBS letters 2018, 592, 3229–3238. [Google Scholar] [CrossRef] [Green Version]
- Wyatt, J.W.; Korasick, D.A.; Qureshi, I.A.; Campbell, A.C.; Gates, K.S.; Tanner, J.J. Inhibition, crystal structures, and in-solution oligomeric structure of aldehyde dehydrogenase 9A1. Arch. Biochem. Biophys. 2020, 691, 108477. [Google Scholar] [CrossRef] [PubMed]
- Shortall, K.; Djeghader, A.; Magner, E.; Soulimane, T. Insights into Aldehyde Dehydrogenase Enzymes: A Structural Perspective. Front. Mol. Biosci. 2021, 8, 410. [Google Scholar] [CrossRef] [PubMed]
- Inagaki, E.; Ohshima, N.; Takahashi, H.; Kuroishi, C.; Yokoyama, S.; Tahirov, T. Crystal structure of Thermus thermophilus Δ1-pyrroline-5-carboxylate dehydrogenase. J. Mol. Biol. 2006, 362, 490–501. [Google Scholar] [CrossRef] [PubMed]
- Luo, M.; Singh, R.K.; Tanner, J.J. Structural determinants of oligomerization of Δ1-pyrroline-5-carboxylate dehydrogenase: Identification of a hexamerization hot spot. J. Mol. Biol. 2013, 425, 3106–3120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pemberton, T.A.; Srivastava, D.; Sanyal, N.; Henzl, M.T.; Becker, D.F.; Tanner, J.J. Structural studies of yeast Δ1-pyrroline-5-carboxylate dehydrogenase (ALDH4A1): Active site flexibility and oligomeric state. Biochemistry 2014, 53, 1350–1359. [Google Scholar] [CrossRef]
- Feldman, R.I.; Weiner, H. Horse liver aldehyde dehydrogenase II. Kinetics and mechanistic implications of the dehydrogenase and esterase activity. J. Biol. Chem. 1972, 247, 267–272. [Google Scholar] [CrossRef]
- MAcGIBBON, A.K.; Haylock, S.J.; Buckley, P.D.; Blackwell, L. Kinetic studies on the esterase activity of cytoplasmic sheep liver aldehyde dehydrogenase. Biochem. J. 1978, 171, 533–538. [Google Scholar] [CrossRef] [Green Version]
- Sydow, K.; Daiber, A.; Oelze, M.; Chen, Z.; August, M.; Wendt, M.; Ullrich, V.; Mülsch, A.; Schulz, E.; Keaney, J.F. Central role of mitochondrial aldehyde dehydrogenase and reactive oxygen species in nitroglycerin tolerance and cross-tolerance. J. Clin. Investig. 2004, 113, 482–489. [Google Scholar] [CrossRef] [PubMed]
- Duncan, R. Aldehyde dehydrogenase. An enzyme with two distinct catalytic activities at a single type of active site. Biochem. J. 1985, 230, 261–267. [Google Scholar] [CrossRef] [Green Version]
- Koppaka, V.; Thompson, D.C.; Chen, Y.; Ellermann, M.; Nicolaou, K.C.; Juvonen, R.O.; Petersen, D.; Deitrich, R.A.; Hurley, T.D.; Vasiliou, V. Aldehyde dehydrogenase inhibitors: A comprehensive review of the pharmacology, mechanism of action, substrate specificity, and clinical application. Pharmacol. Rev. 2012, 64, 520–539. [Google Scholar] [CrossRef] [Green Version]
- Daiber, A.; Oelze, M.; Coldewey, M.; Bachschmid, M.; Wenzel, P.; Sydow, K.; Wendt, M.; Kleschyov, A.L.; Stalleicken, D.; Ullrich, V. Oxidative stress and mitochondrial aldehyde dehydrogenase activity: A comparison of pentaerythritol tetranitrate with other organic nitrates. Mol. Pharmacol. 2004, 66, 1372–1382. [Google Scholar] [CrossRef]
- Mann, C.J.; Weiner, H. Differences in the roles of conserved glutamic acid residues in the active site of human class 3 and class 2 aldehyde dehydrogenases. Protein Sci. 1999, 8, 1922–1929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoshino, T.; Yamabe, E.; Hawari, M.A.; Tamura, M.; Kanamaru, S.; Yoshida, K.; Koesoema, A.A.; Matsuda, T. Oxidation of aromatic and aliphatic aldehydes to carboxylic acids by Geotrichum candidum aldehyde dehydrogenase. Tetrahedron 2020, 76, 131387. [Google Scholar] [CrossRef]
- Ambroziak, W.; Pietruszko, R. Human aldehyde dehydrogenase. Activity with aldehyde metabolites of monoamines, diamines, and polyamines. J. Biol. Chem. 1991, 266, 13011–13018. [Google Scholar] [CrossRef]
- Kitamura, T.; Naganuma, T.; Abe, K.; Nakahara, K.; Ohno, Y.; Kihara, A. Substrate specificity, plasma membrane localization, and lipid modification of the aldehyde dehydrogenase ALDH3B1. Biochim. Et Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2013, 1831, 1395–1401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ouyang, Y.; Li, Q.; Kuang, X.; Wang, H.; Wu, J.; Ayepa, E.; Chen, H.; Abrha, G.T.; Zhang, Z.; Li, X. YMR152W from Saccharomyces cerevisiae encoding a novel aldehyde reductase for detoxification of aldehydes derived from lignocellulosic biomass. J. Biosci. Bioeng. 2020, 131, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, S.; Xiao, T.; Vergara, L.A.; Srivastava, S.; Nees, D.; Piatigorsky, J.; Ansari, N.H. Role of aldehyde dehydrogenase isozymes in the defense of rat lens and human lens epithelial cells against oxidative stress. Investig. Ophthalmol. Vis. Sci. 2005, 46, 259–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchitti, S.A.; Deitrich, R.A.; Vasiliou, V. Neurotoxicity and metabolism of the catecholamine-derived 3, 4-dihydroxyphenylacetaldehyde and 3, 4-dihydroxyphenylglycolaldehyde: The role of aldehyde dehydrogenase. Pharmacol. Rev. 2007, 59, 125–150. [Google Scholar] [CrossRef] [PubMed]
- Pequerul, R.; Vera, J.; Giménez-Dejoz, J.; Crespo, I.; Coines, J.; Porté, S.; Rovira, C.; Parés, X.; Farrés, J. Structural and kinetic features of aldehyde dehydrogenase 1A (ALDH1A) subfamily members, cancer stem cell markers active in retinoic acid biosynthesis. Arch. Biochem. Biophys. 2020, 681, 108256. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, A.; Hsu, L.C.; Davé, V. Retinal oxidation activity and biological role of human cytosolic aldehyde dehydrogenase. Enzyme 1992, 46, 239–244. [Google Scholar] [CrossRef]
- Marchitti, S.A.; Chen, Y.; Thompson, D.C.; Vasiliou, V. Ultraviolet radiation: Cellular antioxidant response and the role of ocular aldehyde dehydrogenase enzymes. Eye Contact Lens 2011, 37, 206. [Google Scholar] [CrossRef] [Green Version]
- Voulgaridou, G.-P.; Tsochantaridis, I.; Tolkas, C.; Franco, R.; Giatromanolaki, A.; Panayiotidis, M.I.; Pappa, A. Aldehyde dehydrogenase 3A1 confers oxidative stress resistance accompanied by altered DNA damage response in human corneal epithelial cells. Free. Radic. Biol. Med. 2020, 150, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Piatigorsky, J. Multifunctional lens crystallins and corneal enzymes: More than meets the eye. Ann. N. Y. Acad. Sci. 1998, 842, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Moreb, J.S. Aldehyde dehydrogenase as a marker for stem cells. Curr. Stem Cell Res. Ther. 2008, 3, 237–246. [Google Scholar] [CrossRef] [PubMed]
- Vassalli, G. Aldehyde dehydrogenases: Not just markers, but functional regulators of stem cells. Stem Cells Int. 2019, 2019, 3904645–3904660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.-K.; Tanner, J.J. Crystal structure of aldehyde dehydrogenase 16 reveals trans-hierarchical structural similarity and a new dimer. J. Mol. Biol. 2019, 431, 524–541. [Google Scholar] [CrossRef]
- Hayes, K.; Noor, M.; Djeghader, A.; Armshaw, P.; Pembroke, T.; Tofail, S.; Soulimane, T. The quaternary structure of Thermus thermophilus aldehyde dehydrogenase is stabilized by an evolutionary distinct C-terminal arm extension. Sci. Rep. 2018, 8, 1–14. [Google Scholar]
- Riveros-Rosas, H.; Julián-Sánchez, A.; Moreno-Hagelsieb, G.; Muñoz-Clares, R.A. Aldehyde dehydrogenase diversity in bacteria of the Pseudomonas genus. Chemico-Biol. Interact. 2019, 304, 83–87. [Google Scholar] [CrossRef]
- Miah, R.; Nina, S.; Murate, T.; Kataoka, N.; Matsutani, M.; Matsushita, K.; Yakushi, T. Major aldehyde dehydrogenase AldFGH of Gluconacetobacter diazotrophicus is independent of pyrroloquinoline quinone but dependent on molybdopterin for acetic acid fermentation. Appl. Microbiol. Biotechnol. 2021, 105, 2341–2350. [Google Scholar] [CrossRef] [PubMed]
- Hempel, J.; Kaiser, R.; Jörnvall, H. Mitochondrial aldehyde dehydrogenase from human liver: Primary structure, differences in relation to the cytosolic enzyme, and functional correlations. Eur. J. Biochem. 1985, 153, 13–28. [Google Scholar] [CrossRef]
- Boyd, L.A.; Adam, L.; Pelcher, L.E.; McHughen, A.; Hirji, R.; Selvaraj, G. Characterization of an Escherichia coli gene encoding betaine aldehyde dehydrogenase (BADH): Structural similarity to mammalian ALDHs and a plant BADH. Gene 1991, 103, 45–52. [Google Scholar] [CrossRef]
- Kato, T.; Miyanaga, A.; Kanaya, S.; Morikawa, M. Gene cloning and characterization of an aldehyde dehydrogenase from long-chain alkane-degrading Geobacillus thermoleovorans B23. Extremophiles 2010, 14, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Miles, J.A.; Machattou, P.; Nevin-Jones, D.; Webb, M.E.; Millard, A.; Scanlan, D.J.; Taylor, P.C. Identification of a cyanobacterial aldehyde dehydrogenase that produces retinoic acid in vitro. Biochem. Biophys. Res. Commun. 2019, 510, 27–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, S.-H.; Ngo, H.-P.-T.; Nam, H.-K.; Kim, K.-R.; Kang, L.-W.; Oh, D.-K. Alternative biotransformation of retinal to retinoic acid or retinol by an aldehyde dehydrogenase from Bacillus cereus. Appl. Environ. Microbiol. 2016, 82, 3940–3946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coitinho, J.B.; Pereira, M.S.; Costa, D.b.M.; Guimaraes, S.L.; Araujo, S.S.; Hengge, A.C.; Brandao, T.A.; Nagem, R.A. Structural and kinetic properties of the aldehyde dehydrogenase NahF, a broad substrate specificity enzyme for aldehyde oxidation. Biochemistry 2016, 55, 5453–5463. [Google Scholar] [CrossRef]
- Li, X.; Li, Y.; Wei, D.; Li, P.; Wang, L.; Feng, L. Characterization of a broad-range aldehyde dehydrogenase involved in alkane degradation in Geobacillus thermodenitrificans NG80-2. Microbiol. Res. 2010, 165, 706–712. [Google Scholar] [CrossRef]
- Knaus, T.; Tseliou, V.; Humphreys, L.D.; Scrutton, N.S.; Mutti, F.G. A biocatalytic method for the chemoselective aerobic oxidation of aldehydes to carboxylic acids. Green Chem. 2018, 20, 3931–3943. [Google Scholar] [CrossRef]
- Wu, S.; Zhou, Y.; Seet, D.; Li, Z. Regio-and Stereoselective Oxidation of Styrene Derivatives to Arylalkanoic Acids via One-Pot Cascade Biotransformations. Adv. Synth. Catal. 2017, 359, 2132–2141. [Google Scholar] [CrossRef] [Green Version]
- Winkler, T.; Gröger, H.; Hummel, W. Enantioselective Rearrangement Coupled with Water Addition: Direct Synthesis of Enantiomerically Pure Saturated Carboxylic Acids from α, β-Unsaturated Aldehydes. ChemCatChem 2014, 6, 961–964. [Google Scholar] [CrossRef]
- Atalah, J.; Cáceres-Moreno, P.; Espina, G.; Blamey, J.M. Thermophiles and the applications of their enzymes as new biocatalysts. Bioresour. Technol. 2019, 280, 478–488. [Google Scholar] [CrossRef] [PubMed]
- Lyons, J.A.; Aragão, D.; Slattery, O.; Pisliakov, A.V.; Soulimane, T.; Caffrey, M. Structural insights into electron transfer in caa 3-type cytochrome oxidase. Nature 2012, 487, 514–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimmerling, J.; Oelschlägel, M.; Großmann, C.; Voitel, M.; Schlömann, M.; Tischler, D. Biochemical Characterization of Phenylacetaldehyde Dehydrogenases from Styrene-degrading Soil Bacteria. Appl. Biochem. Biotechnol. 2021, 193, 650–667. [Google Scholar] [CrossRef]
- Vasiliou, V.; Bairoch, A.; Tipton, K.F.; Nebert, D.W. Eukaryotic aldehyde dehydrogenase (ALDH) genes: Human polymorphisms, and recommended nomenclature based on divergent evolution and chromosomal mapping. Pharm. Genom. 1999, 9, 421–434. [Google Scholar]
- Moretti, A.; Li, J.; Donini, S.; Sobol, R.W.; Rizzi, M.; Garavaglia, S. Crystal structure of human aldehyde dehydrogenase 1A3 complexed with NAD+ and retinoic acid. Sci. Rep. 2016, 6, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sobreira, T.J.; Marlétaz, F.; Simões-Costa, M.; Schechtman, D.; Pereira, A.C.; Brunet, F.; Sweeney, S.; Pani, A.; Aronowicz, J.; Lowe, C.J. Structural shifts of aldehyde dehydrogenase enzymes were instrumental for the early evolution of retinoid-dependent axial patterning in metazoans. Proc. Natl. Acad. Sci. USA 2011, 108, 226–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klyosov, A.A. Kinetics and specificity of human liver aldehyde dehydrogenases toward aliphatic, aromatic, and fused polycyclic aldehydes. Biochemistry 1996, 35, 4457–4467. [Google Scholar] [CrossRef] [PubMed]
- Moore, S.A.; Baker, H.M.; Blythe, T.J.; Kitson, K.E.; Kitson, T.M.; Baker, E.N. Sheep liver cytosolic aldehyde dehydrogenase: The structure reveals the basis for the retinal specificity of class 1 aldehyde dehydrogenases. Structure 1998, 6, 1541–1551. [Google Scholar] [CrossRef] [Green Version]
- Lorentzen, E.; Hensel, R.; Knura, T.; Ahmed, H.; Pohl, E. Structural basis of allosteric regulation and substrate specificity of the non-phosphorylating glyceraldehyde 3-phosphate dehydrogenase from Thermoproteus tenax. J. Mol. Biol. 2004, 341, 815–828. [Google Scholar] [CrossRef]
- Liu, T.; Hao, L.; Wang, R.; Liu, B. Molecular characterization of a thermostable aldehyde dehydrogenase (ALDH) from the hyperthermophilic archaeon Sulfolobus tokodaii strain 7. Extremophiles 2013, 17, 181–190. [Google Scholar] [CrossRef]
- Ettema, T.J.; Ahmed, H.; Geerling, A.C.; van der Oost, J.; Siebers, B. The non-phosphorylating glyceraldehyde-3-phosphate dehydrogenase (GAPN) of Sulfolobus solfataricus: A key-enzyme of the semi-phosphorylative branch of the Entner–Doudoroff pathway. Extremophiles 2008, 12, 75–88. [Google Scholar] [CrossRef] [PubMed]
- Millard, A.; Scanlan, D.J.; Gallagher, C.; Marsh, A.; Taylor, P.C. Unexpected evolutionary proximity of eukaryotic and cyanobacterial enzymes responsible for biosynthesis of retinoic acid and its oxidation. Mol. BioSystems 2014, 10, 380–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brash, A.R.; Niraula, N.P.; Boeglin, W.E.; Mashhadi, Z. An ancient relative of cyclooxygenase in cyanobacteria is a linoleate 10S-dioxygenase that works in tandem with a catalase-related protein with specific 10S-hydroperoxide lyase activity. J. Biol. Chem. 2014, 289, 13101–13111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ponting, C.; Aravind, L.; Schultz, J.; Bork, P.; Koonin, E. Eukaryotic signalling domain homologues in archaea and bacteria. Ancient ancestry and horizontal gene transfer. J. Mol. Biol. 1999, 289, 729–745. [Google Scholar] [CrossRef] [PubMed]
- Rawlings, N.D. Bacterial calpains and the evolution of the calpain (C2) family of peptidases. Biol. Direct 2015, 10, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Sun, F.; Zhang, X.-Z.; Myung, S.; Zhang, Y.-H.P. Thermophilic Thermotoga maritima ribose-5-phosphate isomerase RpiB: Optimized heat treatment purification and basic characterization. Protein Expr. Purif. 2012, 82, 302–307. [Google Scholar] [CrossRef]
- Xu, J.; Luo, H.; López, C.; Xiao, J.; Chang, Y. Novel immobilization process of a thermophilic catalase: Efficient purification by heat treatment and subsequent immobilization at high temperature. Bioprocess Biosyst. Eng. 2015, 38, 1983–1991. [Google Scholar] [CrossRef]
- Patchett, M.; Neal, T.; Schofield, L.; Strange, R.; Daniel, R.; Morgan, H. Heat treatment purification of thermostable cellulase and hemicellulase enzymes expressed in E. coli. Enzym. Microb. Technol. 1989, 11, 113–115. [Google Scholar] [CrossRef]
- Pennacchio, A.; Pucci, B.; Secundo, F.; La Cara, F.; Rossi, M.; Raia, C.A. Purification and characterization of a novel recombinant highly enantioselective short-chain NAD (H)-dependent alcohol dehydrogenase from Thermus thermophilus. Appl. Environ. Microbiol. 2008, 74, 3949–3958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inagaki, E.; Takahashi, H.; Kuroishi, C.; Tahirov, T.H. Crystallization and avoiding the problem of hemihedral twinning in crystals of Δ1-pyrroline-5-carboxylate dehydrogenase from Thermus thermophilus. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2005, 61, 609–611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyazaki, K.; Yaoi, T.; Oshima, T. Expression, purification, and substrate specificity of isocitrate dehydrogenase from Thermus thermophilus HB8. Eur. J. Biochem. 1994, 221, 899–903. [Google Scholar] [CrossRef]
- Ng, M.Y.; Tan, W.S.; Abdullah, N.; Ling, T.C.; Tey, B.T. Heat treatment of unclarified Escherichia coli homogenate improved the recovery efficiency of recombinant hepatitis B core antigen. J. Virol. Methods 2006, 137, 134–139. [Google Scholar] [CrossRef]
- Kalthoff, C. A novel strategy for the purification of recombinantly expressed unstructured protein domains. J. Chromatogr. B 2003, 786, 247–254. [Google Scholar] [CrossRef]
- Jonasson, P.; Nygren, P.-Å.; Jörnvall, H.; Johansson, B.-L.; Wahren, J.; Uhlén, M.; Ståhl, S. Integrated bioprocess for production of human proinsulin C-peptide via heat release of an intracellular heptameric fusion protein. J. Biotechnol. 2000, 76, 215–226. [Google Scholar] [CrossRef]
- Oshima, T.; Imahori, K. Description of Thermus thermophilus (Yoshida and Oshima) comb. nov., a nonsporulating thermophilic bacterium from a Japanese thermal spa. Int. J. Syst. Evol. Microbiol. 1974, 24, 102–112. [Google Scholar] [CrossRef] [Green Version]
- Ohtani, N.; Tomita, M.; Itaya, M. An extreme thermophile, Thermus thermophilus, is a polyploid bacterium. J. Bacteriol. 2010, 192, 5499–5505. [Google Scholar] [CrossRef] [Green Version]
- Miyamoto, T.; Moriya, T.; Homma, H.; Oshima, T. Enzymatic properties and physiological function of glutamate racemase from Thermus thermophilus. Biochim. Biophys. Acta (BBA)-Proteins Proteom. 2020, 1868, 140461. [Google Scholar] [CrossRef] [PubMed]
- Kozono, I.; Hibi, M.; Takeuchi, M.; Ogawa, J. Purification and characterization of molybdenum-containing aldehyde dehydrogenase that oxidizes benzyl maltol derivative from Pseudomonas nitroreducens SB32154. Biosci. Biotechnol. Biochem. 2020, 84, 2390–2400. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Kaufholdt, D.; Awadalah, M.; Hänsch, R.; Beerhues, L.; Gaid, M. Cytosolic aromatic aldehyde dehydrogenase provides benzoic acid for xanthone biosynthesis in Hypericum. Plant Physiol. Biochem. 2021, 160, 82–93. [Google Scholar] [CrossRef] [PubMed]
- Pang, J.; Zheng, M.; Sun, R.; Wang, A.; Wang, X.; Zhang, T. Synthesis of ethylene glycol and terephthalic acid from biomass for producing PET. Green Chem. 2016, 18, 342–359. [Google Scholar] [CrossRef]
- Tomas, R.R.A.; Bordado, J.O.C.; Gomes, J.O.F. p-Xylene oxidation to terephthalic acid: A literature review oriented toward process optimization and development. Chem. Rev. 2013, 113, 7421–7469. [Google Scholar] [CrossRef]
- Ho, K.K.; Hurley, T.D.; Weiner, H. Selective alteration of the rate-limiting step in cytosolic aldehyde dehydrogenase through random mutagenesis. Biochemistry 2006, 45, 9445–9453. [Google Scholar] [CrossRef] [PubMed]
- Sidhu, R.S.; Blair, A.H. Human liver aldehyde dehydrogenase. Esterase activity. J. Biol. Chem. 1975, 250, 7894–7898. [Google Scholar] [CrossRef]
- Ferrández, A.; Prieto, M.a.A.; Garcia, J.L.; Diaz, E. Molecular characterization of PadA, a phenylacetaldehyde dehydrogenase from Escherichia coli. FEBS Lett. 1997, 406, 23–27. [Google Scholar] [CrossRef] [Green Version]
- Blackwell, L.F.; Bennett, A.F.; Buckley, P.D. Relationship between the mechanisms of the esterase and dehydrogenase activities of the cytoplasmic aldehyde dehydrogenase from sheep liver. An alternative view. Biochemistry 1983, 22, 3784–3791. [Google Scholar] [CrossRef]
- Lindahl, R.; Evces, S. Rat liver aldehyde dehydrogenase. I. Isolation and characterization of four high Km normal liver isozymes. J. Biol. Chem. 1984, 259, 11986–11990. [Google Scholar] [CrossRef]
- Rodríguez-Zavala, J.S.; Ortiz-Cruz, M.A.; Moreno-Sanchez, R. Characterization of an aldehyde dehydrogenase from Euglena gracilis. J. Eukaryot. Microbiol. 2006, 53, 36–42. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrate | °C | KM (mM) | Vmax (µmol/min) | kcat (s−1) |
|---|---|---|---|---|
| Hexanal | 25 °C | 1.08 ± 0.46 | 0.0021 ± 0.0006 | 0.51 ± 0.15 |
| 50 °C | 0.99 ± 0.24 | 0.027 ± 0.0043 | 5.71 ± 0.64 | |
| Terephthalaldehyde | 25 °C | 0.11 ± 0.067 | 0.0041 ± 0.001 | 1.05 ± 0.27 |
| 50 °C | 0.38 ± 0.11 | 0.047 ± 0.010 | 12.08 ± 2.50 | |
| Benzaldehyde | 25 °C | 1.41 ± 1.15 | 0.0008 ± 0.0003 | 0.19 ± 0.09 |
| 50 °C | 1.52 ± 0.51 | 0.0057 ± 0.001 | 1.46 ± 0.29 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shortall, K.; Durack, E.; Magner, E.; Soulimane, T. Study of ALDH from Thermus thermophilus—Expression, Purification and Characterisation of the Non-Substrate Specific, Thermophilic Enzyme Displaying Both Dehydrogenase and Esterase Activity. Cells 2021, 10, 3535. https://doi.org/10.3390/cells10123535
Shortall K, Durack E, Magner E, Soulimane T. Study of ALDH from Thermus thermophilus—Expression, Purification and Characterisation of the Non-Substrate Specific, Thermophilic Enzyme Displaying Both Dehydrogenase and Esterase Activity. Cells. 2021; 10(12):3535. https://doi.org/10.3390/cells10123535
Chicago/Turabian StyleShortall, Kim, Edel Durack, Edmond Magner, and Tewfik Soulimane. 2021. "Study of ALDH from Thermus thermophilus—Expression, Purification and Characterisation of the Non-Substrate Specific, Thermophilic Enzyme Displaying Both Dehydrogenase and Esterase Activity" Cells 10, no. 12: 3535. https://doi.org/10.3390/cells10123535
APA StyleShortall, K., Durack, E., Magner, E., & Soulimane, T. (2021). Study of ALDH from Thermus thermophilus—Expression, Purification and Characterisation of the Non-Substrate Specific, Thermophilic Enzyme Displaying Both Dehydrogenase and Esterase Activity. Cells, 10(12), 3535. https://doi.org/10.3390/cells10123535