Proteomic Adaptation of Clostridioides difficile to Treatment with the Antimicrobial Peptide Nisin

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Purification of Nisin
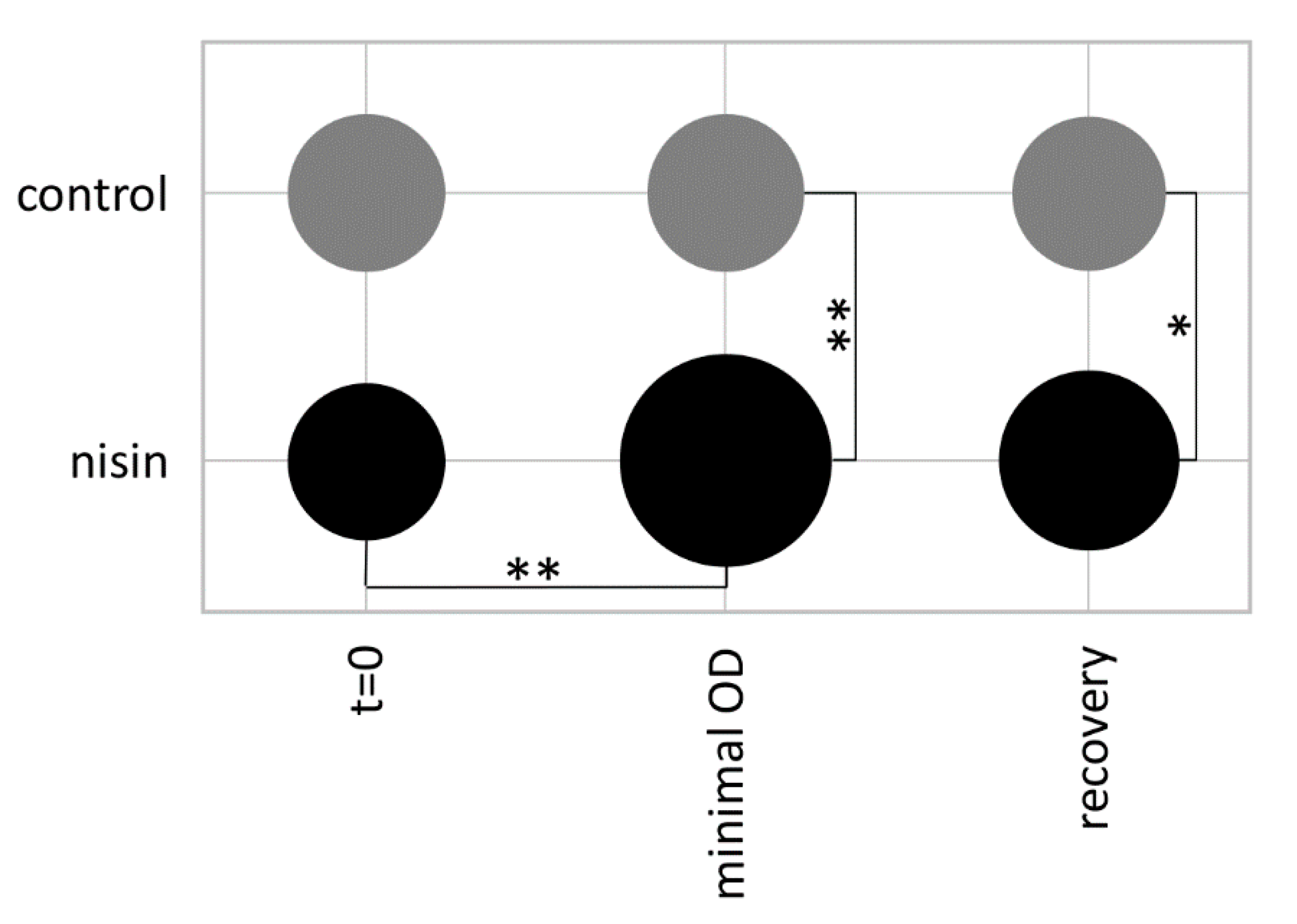
2.2. Bacterial Growth and Nisin Treatment for Proteomic Analysis
2.3. Sample Preparation for Mass Spectrometry
2.4. Liquid Chromatography and Mass Spectrometric Analysis
2.5. Data Processing and Data Analysis
2.6. Electron Microscopy
3. Results
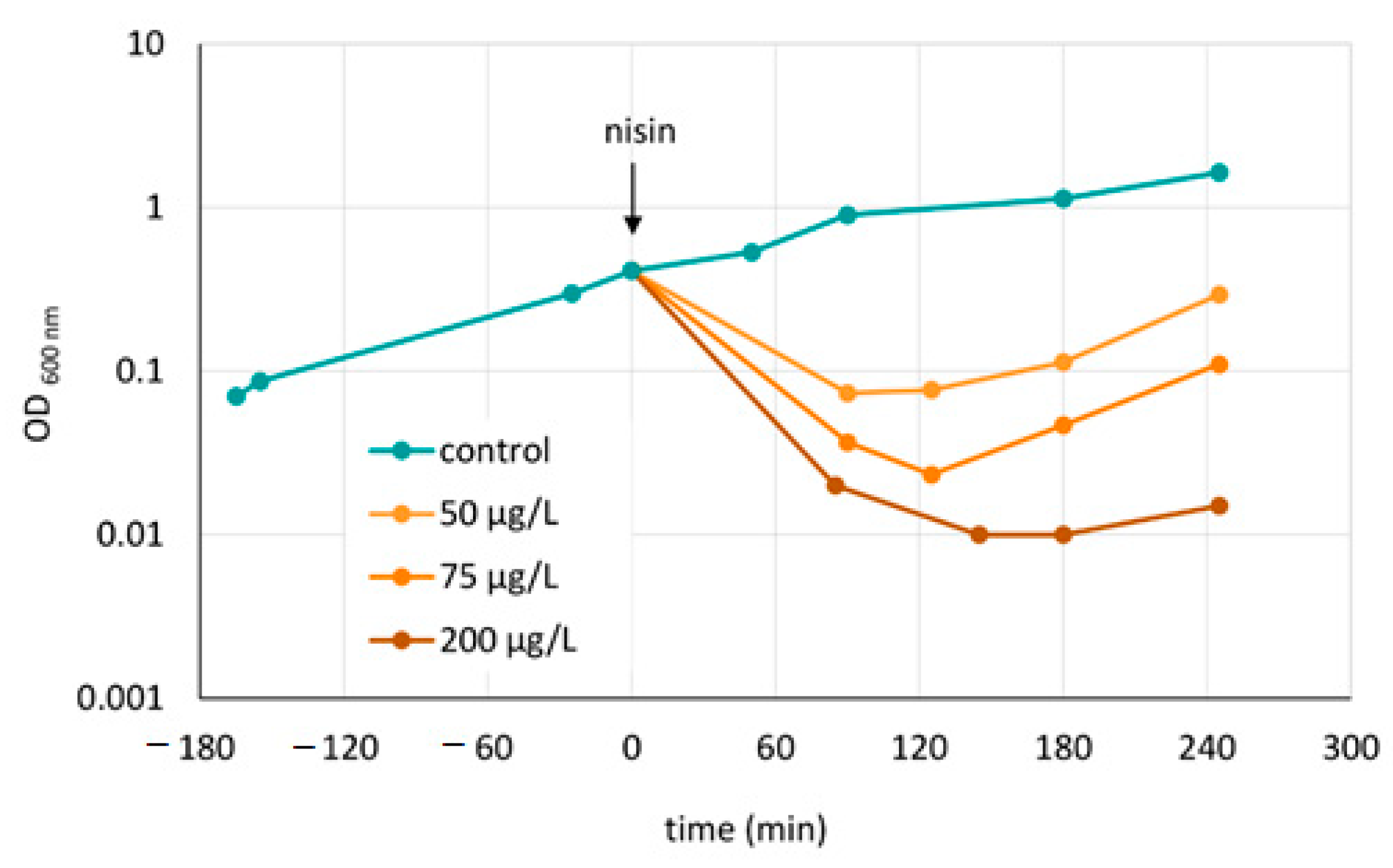
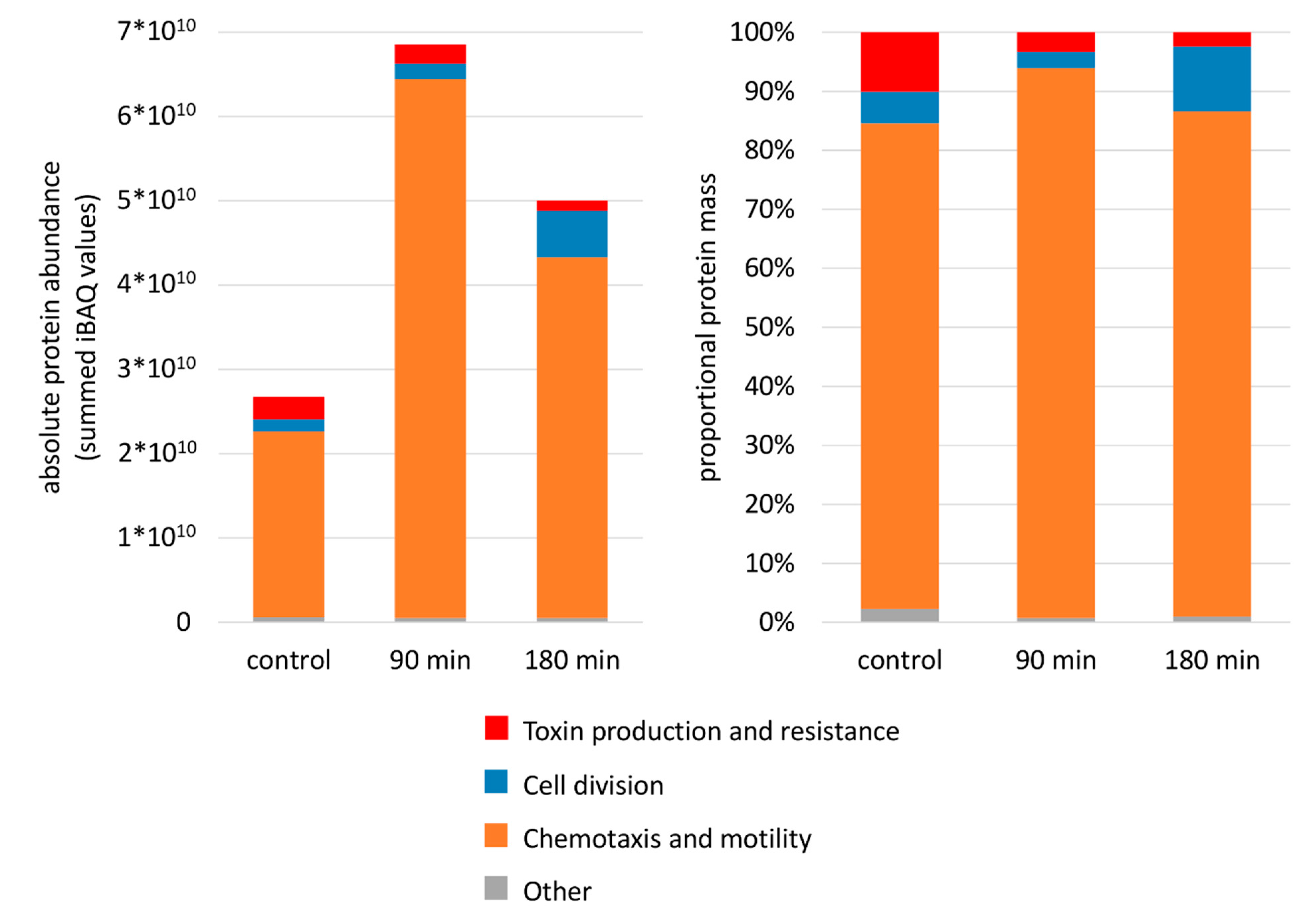
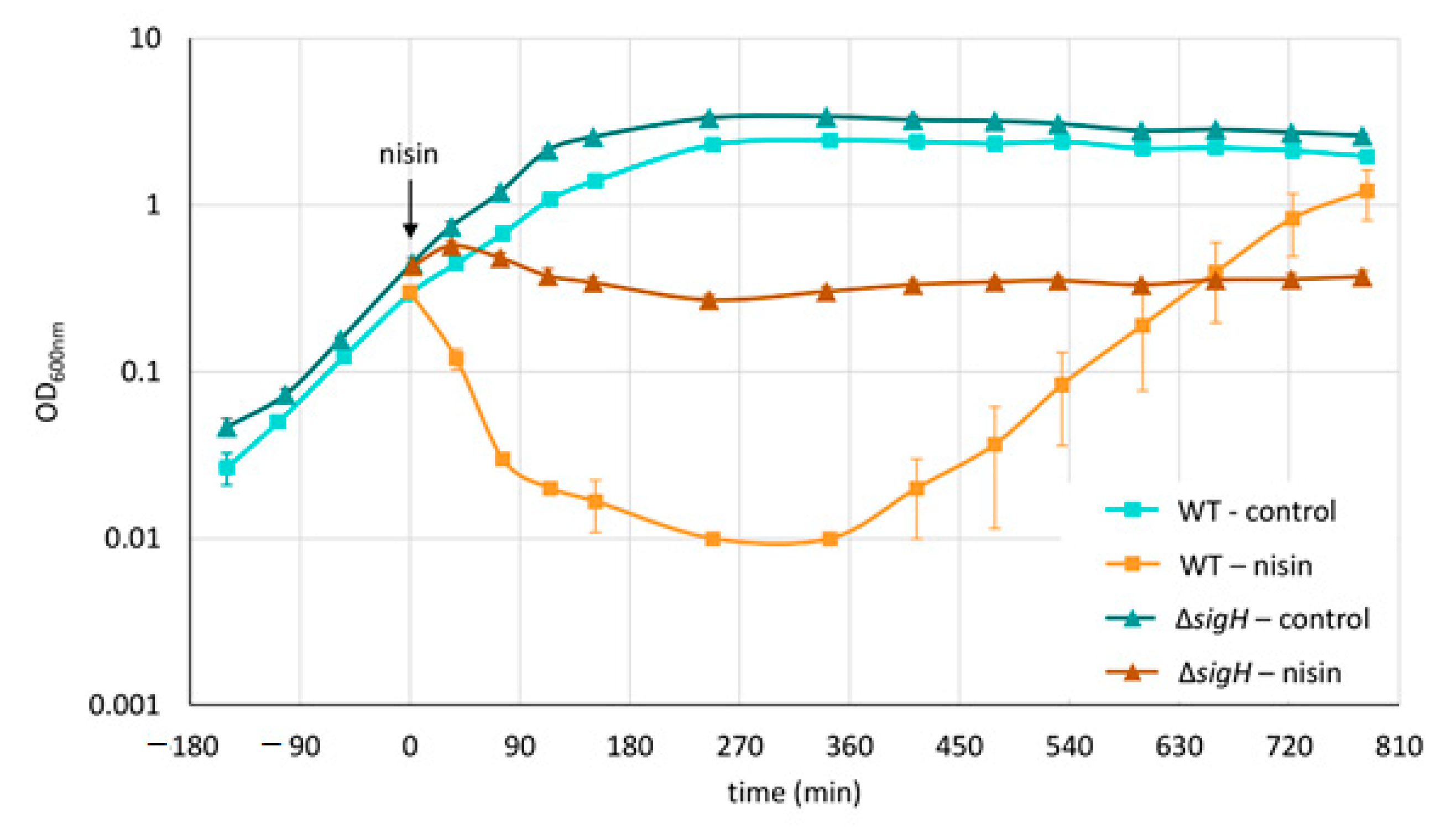
3.1. Phenotypic and Proteomic Adaptation
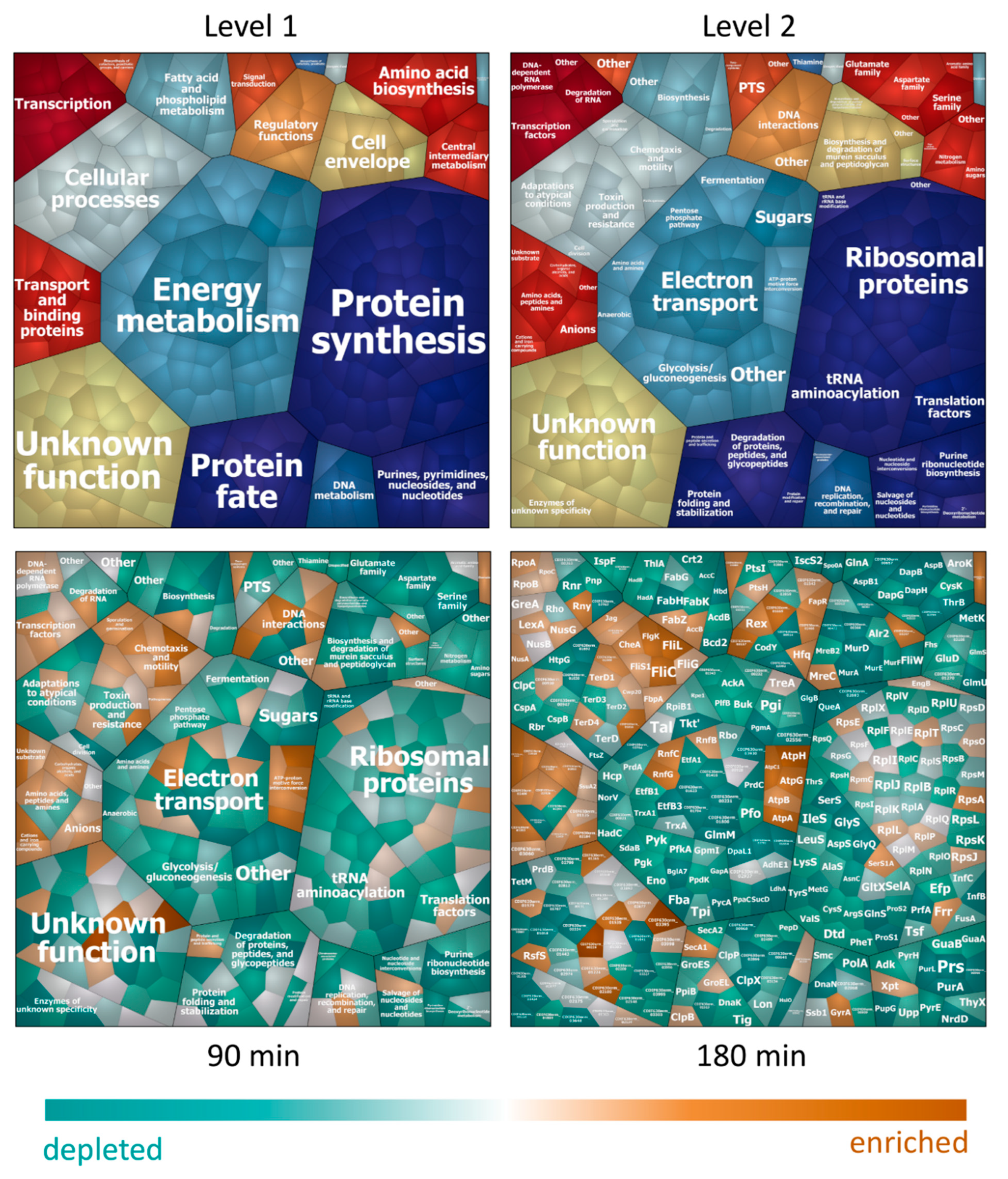
3.2. Adaptation in the Cytosol
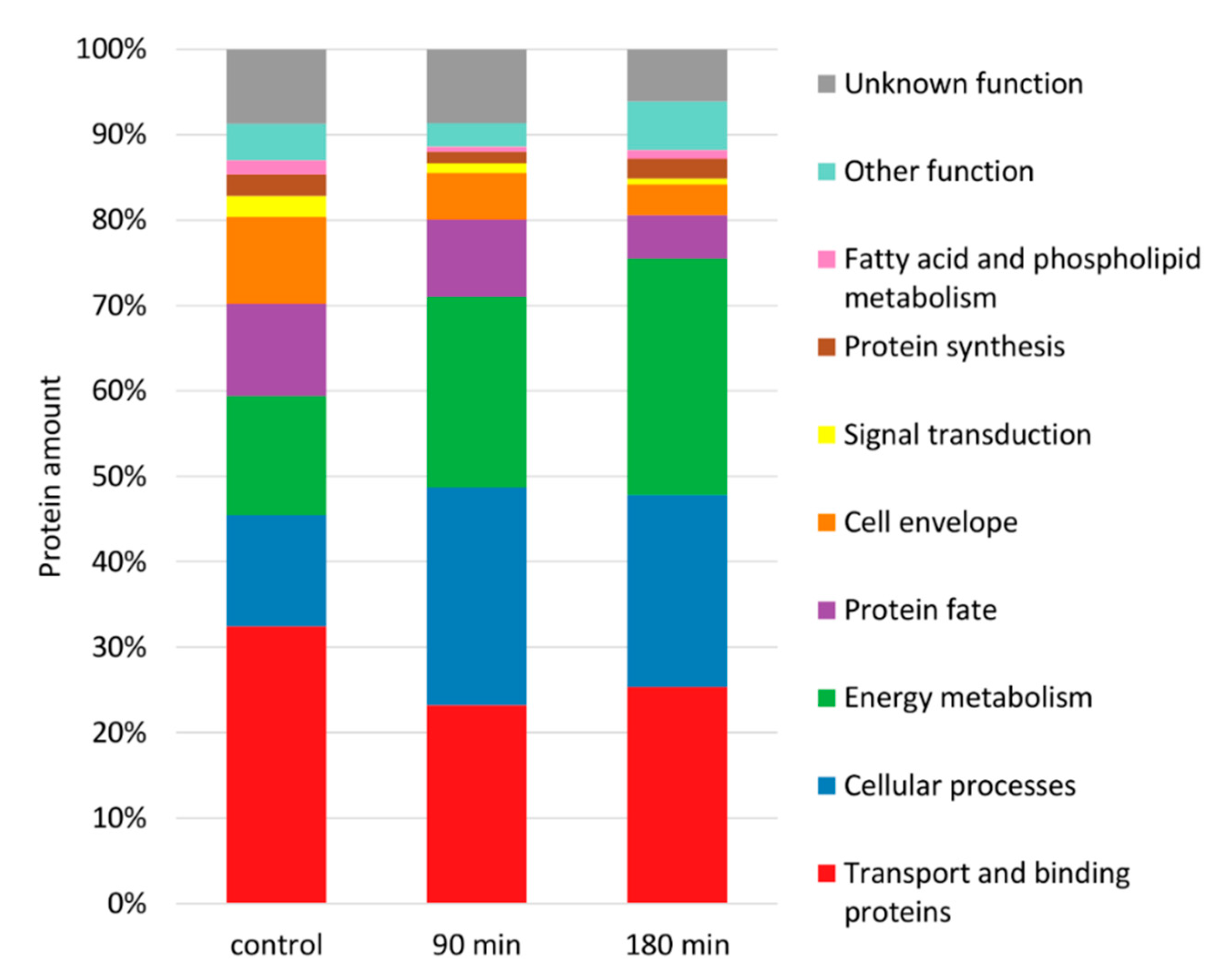
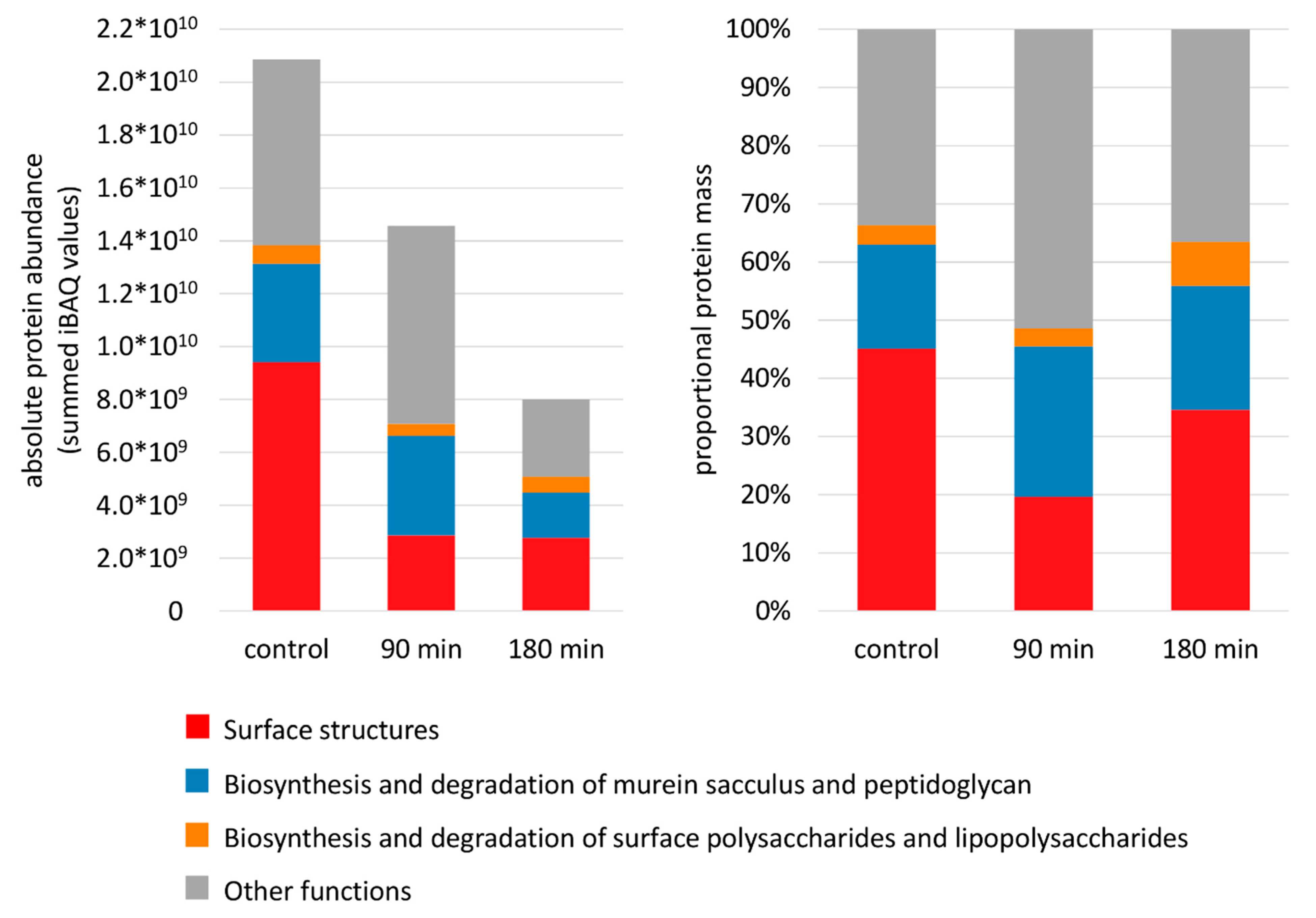
3.3. Composition of the Membrane Proteome during Adaptation to Nisin
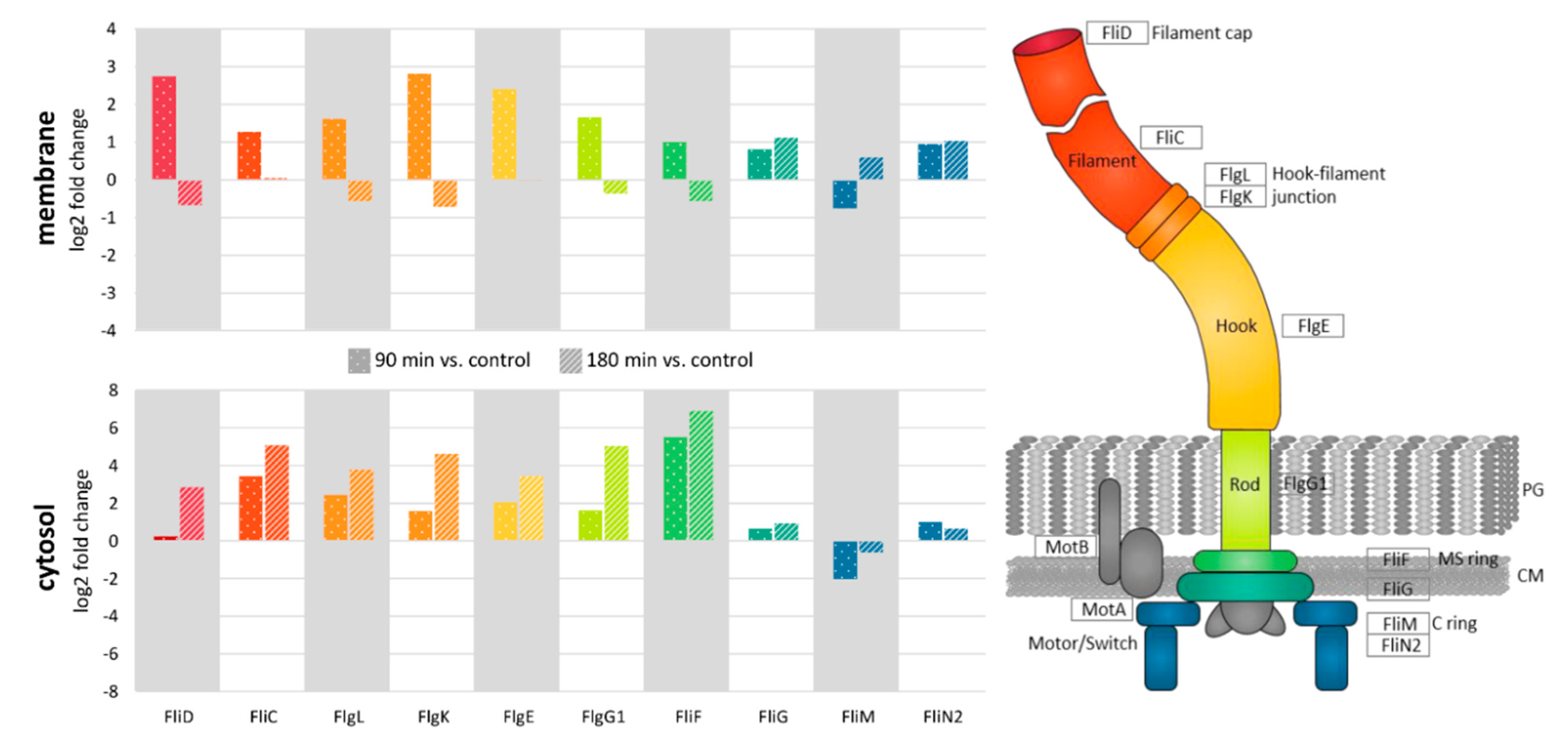
3.4. Relocalization of Proteins in Response to Nisin
4. Discussion
4.1. Differences between Early Phase and Fully Established Adaptation or Resistance
4.2. Role of Flagella in Virulence and Resistance
4.3. Lipid II as Target for Antimicrobials
4.4. Bactericidal Activities of Various Membrane-Active Agents Against C. difficile
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Drider, D.; Rebuffat, S. Prokaryotic Antimicrobial Peptides: From Genes to Applications; Springer: New York, NY, USA, 2011. [Google Scholar]
- Dischinger, J.; Basi Chipalu, S.; Bierbaum, G. Lantibiotics: Promising Candidates for Future Applications in Health Care. Int. J. Med. Microbiol. 2014, 304, 51–62. [Google Scholar] [CrossRef]
- Bower, C.K.; Parker, J.E.; Higgins, A.Z.; Oest, M.E.; Wilson, J.T.; Valentine, B.A.; Bothwell, M.K.; McGuire, J. Protein Antimicrobial Barriers to Bacterial Adhesion: In Vitro and in Vivo Evaluation of Nisin-Treated Implantable Materials. Coll. Surf. B Biointerfaces 2002, 25, 81–90. [Google Scholar] [CrossRef]
- Dicks, L.M.T.; Heunis, T.D.J.; van Staden, D.A.; Brand, A.; Noll, K.S.; Chikindas, M.L. Medical and personal care applications of bacteriocins produced by lactic acid bacteria. In Prokaryotic Antimicrobial Peptides: From Genes to Applications; Drider, D., Rebuffat, S., Eds.; Springer: New York, NY, USA, 2011; pp. 391–421. [Google Scholar]
- García, P.; Rodríguez, L.; Rodríguez, A.; Martínez, B. Food Biopreservation: Promising Strategies Using Bacteriocins, Bacteriophages and Endolysins. Trends Food Sci. Technol. 2010, 21, 373–382. [Google Scholar] [CrossRef] [Green Version]
- Delves-Broughton, J.; Blackburn, P.; Evans, R.J.; Hugenholtz, J. Applications of the Bacteriocin, Nisin. Antonie Van Leeuwenhoek 1996, 69, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, B.P.; Wei, J.; Greenberg, K.; Novick, R. Activity of Nisin against Streptococcus pneumoniae, in Vitro, and in a Mouse Infection Model. J. Antimicrob. Chemother. 1998, 42, 277–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brumfitt, W.; Salton, M.R.J.; Hamilton-Miller, J.M.T. Nisin, Alone and Combined with Peptidoglycan-Modulating Antibiotics: Activity against Methicillin-Resistant Staphylococcus aureus and Vancomycin-Resistant Enterococci. J. Antimicrob. Chemother. 2002, 50, 731–734. [Google Scholar] [CrossRef] [Green Version]
- Dosler, S.; Gerceker, A.A. In Vitro Activities of Nisin Alone or in Combination with Vancomycin and Ciprofloxacin against Methicillin-Resistant and Methicillin-Susceptible Staphylococcus aureus Strains. Chemotherapy 2011, 57, 511–516. [Google Scholar] [CrossRef] [PubMed]
- Wiedemann, I.; Breukink, E.; van Kraaij, C.; Kuipers, O.P.; Bierbaum, G.; de Kruijff, B.; Sahl, H.-G. Specific Binding of Nisin to the Peptidoglycan Precursor Lipid II Combines Pore Formation and Inhibition of Cell Wall Biosynthesis for Potent Antibiotic Activity. J. Biol. Chem. 2001, 276, 1772–1779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brötz, H.; Josten, M.; Wiedemann, I.; Schneider, U.; Götz, F.; Bierbaum, G.; Sahl, H.-G. Role of Lipid-Bound Peptidoglycan Precursors in the Formation of Pores by Nisin, Epidermin and Other Lantibiotics. Mol. Microbiol. 1998, 30, 317–327. [Google Scholar] [CrossRef] [Green Version]
- Hsu, S.-T.D.; Breukink, E.; Tischenko, E.; Lutters, M.A.G.; de Kruijff, B.; Kaptein, R.; Bonvin, A.M.J.J.; van Nuland, N.A.J. The Nisin-Lipid II Complex Reveals a Pyrophosphate Cage That Provides a Blueprint for Novel Antibiotics. Nat. Struct. Mol. Biol. 2004, 11, 963–967. [Google Scholar] [CrossRef] [Green Version]
- Bonev, B.B.; Breukink, E.; Swiezewska, E.; De Kruijff, B.; Watts, A. Targeting Extracellular Pyrophosphates Underpins the High Selectivity of Nisin. FASEB J. 2004, 18, 1862–1869. [Google Scholar] [CrossRef]
- Hasper, H.E.; Kramer, N.E.; Smith, J.L.; Hillman, J.D.; Zachariah, C.; Kuipers, O.P.; de Kruijff, B.; Breukink, E. An Alternative Bactericidal Mechanism of Action for Lantibiotic Peptides That Target Lipid II. Science 2006, 313, 1636–1637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Draper, L.A.; Cotter, P.D.; Hill, C.; Ross, R.P. Lantibiotic Resistance. Microbiol. Mol. Biol. Rev. MMBR 2015, 79, 171–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasper, H.E.; de Kruijff, B.; Breukink, E. Assembly and Stability of Nisin-Lipid II Pores. Biochemistry 2004, 43, 11567–11575. [Google Scholar] [CrossRef]
- Sahl, H.-G.; Bierbaum, G. Lantibiotics: Mode of Action, Biosynthesis and Bioengineering. Curr. Pharm. Biotechnol. 2009, 10, 2–18. [Google Scholar]
- Le Lay, C.; Dridi, L.; Bergeron, M.G.; Ouellette, M.; Fliss, I.L. Nisin Is an Effective Inhibitor of Clostridium difficile Vegetative Cells and Spore Germination. J. Med. Microbiol. 2016, 65, 169–175. [Google Scholar] [CrossRef]
- Slimings, C.; Riley, T.V. Antibiotics and Hospital-Acquired Clostridium Difficile Infection: Update of Systematic Review and Meta-Analysis. J. Antimicrob. Chemother. 2014, 69, 881–891. [Google Scholar] [CrossRef]
- Bartlett, J.G. Narrative Review: The New Epidemic of Clostridium difficile-Associated Enteric Disease. Ann. Intern. Med. 2006, 145, 758–764. [Google Scholar] [CrossRef]
- Rupnik, M.; Wilcox, M.H.; Gerding, D.N. Clostridium difficile Infection: New Developments in Epidemiology and Pathogenesis. Nat. Rev. Microbiol. 2009, 7, 526–536. [Google Scholar] [CrossRef]
- Viswanathan, V.K.; Mallozzi, M.J.; Vedantam, G. Clostridium Difficile Infection: An Overview of the Disease and Its Pathogenesis, Epidemiology and Interventions. Gut Microbes 2010, 1, 234–242. [Google Scholar] [CrossRef] [Green Version]
- Perego, M.; Glaser, P.; Minutello, A.; Strauch, M.A.; Leopold, K.; Fischer, W. Incorporation of D-Alanine into Lipoteichoic Acid and Wall Teichoic Acid in Bacillus subtilis. Identification of Genes and Regulation. J. Biol. Chem. 1995, 270, 15598–15606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neuhaus, F.C.; Baddiley, J. A Continuum of Anionic Charge: Structures and Functions of D-Alanyl-Teichoic Acids in Gram-Positive Bacteria. Microbiol. Mol. Biol. Rev. MMBR 2003, 67, 686–723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McBride, S.M.; Sonenshein, A.L. The dlt Operon Confers Resistance to Cationic Antimicrobial Peptides in Clostridium difficile. Microbiology 2011, 157, 1457–1465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demel, R.A.; Peelen, T.; Siezen, R.J.; De Kruijff, B.; Kuipers, O.P. Nisin Z, Mutant Nisin Z and Lacticin 481 Interactions with Anionic Lipids Correlate with Antimicrobial Activity. A Monolayer Study. Eur. J. Biochem. 1996, 235, 267–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ming, X.; Daeschel, M.A. Correlation of Cellular Phospholipid Content with Nisin Resistance of Listeria monocytogenes Scott A. J. Food Prot. 1995, 58, 416–420. [Google Scholar] [CrossRef] [PubMed]
- Ming, X.; Daeschel, M.A. Nisin Resistance of Foodborne Bacteria and the Specific Resistance Responses of Listeria monocytogenes Scott A. J. Food Prot. 1993, 56, 944–948. [Google Scholar] [CrossRef] [PubMed]
- Martínez, B.; Rodríguez, A. Antimicrobial Susceptibility of Nisin Resistant Listeria monocytogenes of Dairy Origin. FEMS Microbiol. Lett. 2005, 252, 67–72. [Google Scholar] [CrossRef] [Green Version]
- Staubitz, P.; Neumann, H.; Schneider, T.; Wiedemann, I.; Peschel, A. MprF-Mediated Biosynthesis of Lysylphosphatidylglycerol, an Important Determinant in Staphylococcal Defensin Resistance. FEMS Microbiol. Lett. 2004, 231, 67–71. [Google Scholar] [CrossRef] [Green Version]
- Suárez, J.M.; Edwards, A.N.; McBride, S.M. The Clostridium difficile cpr Locus Is Regulated by a Noncontiguous Two-Component System in Response to Type A and B Lantibiotics. J. Bacteriol. 2013, 195, 2621–2631. [Google Scholar] [CrossRef] [Green Version]
- Clemens, R.; Zaschke-Kriesche, J.; Khosa, S.; Smits, S.H.J. Insight into Two ABC Transporter Families Involved in Lantibiotic Resistance. Front. Mol. Biosci. 2018, 4, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reunanen, J.; Saris, P.E.J. Survival of Nisin Activity in Intestinal Environment. Biotechnol. Lett. 2009, 31, 1229–1232. [Google Scholar] [CrossRef]
- Shin, J.M.; Gwak, J.W.; Kamarajan, P.; Fenno, J.C.; Rickard, A.H.; Kapila, Y.L. Biomedical Applications of Nisin. J. Appl. Microbiol. 2016, 120, 1449–1465. [Google Scholar] [CrossRef] [Green Version]
- Kindrachuk, J.; Jenssen, H.; Elliott, M.; Nijnik, A.; Magrangeas-Janot, L.; Pasupuleti, M.; Thorson, L.; Ma, S.; Easton, D.M.; Bains, M.; et al. Manipulation of Innate Immunity by a Bacterial Secreted Peptide: Lantibiotic Nisin Z Is Selectively Immunomodulatory. Innate Immun. 2013, 19, 315–327. [Google Scholar] [CrossRef]
- De Pablo, M.A.; Gaforio, J.J.; Gallego, A.M.; Ortega, E.; Gálvez, A.M.; Alvarez de Cienfuegos López, G. Evaluation of Immunomodulatory Effects of Nisin-Containing Diets on Mice. FEMS Immunol. Med. Microbiol. 1999, 24, 35–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Begde, D.; Bundale, S.; Mashitha, P.; Rudra, J.; Nashikkar, N.; Upadhyay, A. Immunomodulatory Efficacy of Nisin-a Bacterial Lantibiotic Peptide. J. Pept. Sci. Off. Publ. Eur. Pept. Soc. 2011, 17, 438–444. [Google Scholar] [CrossRef]
- Singh, A.P.; Prabha, V.; Rishi, P. Value Addition in the Efficacy of Conventional Antibiotics by Nisin against Salmonella. PLoS ONE 2013, 8, e76844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gudmundsson, G.H.; Agerberth, B. Neutrophil Antibacterial Peptides, Multifunctional Effector Molecules in the Mammalian Immune System. J. Immunol. Methods 1999, 232, 45–54. [Google Scholar] [CrossRef]
- Müller, C.A.; Autenrieth, I.B.; Peschel, A. Intestinal Epithelial Barrier and Mucosal Immunity. Cell. Mol. Life Sci. 2005, 62, 1297. [Google Scholar] [CrossRef] [PubMed]
- Abts, A.; Mavaro, A.; Stindt, J.; Bakkes, P.J.; Metzger, S.; Driessen, A.J.M.; Smits, S.H.J.; Schmitt, L. Easy and Rapid Purification of Highly Active Nisin. Int. J. Pept. 2011, 2011, 1751455. [Google Scholar] [CrossRef] [PubMed]
- Abts, A.; Montalban-Lopez, M.; Kuipers, O.P.; Smits, S.H.; Schmitt, L. NisC Binds the FxLx Motif of the Nisin Leader Peptide. Biochemistry 2013, 52, 5387–5395. [Google Scholar] [CrossRef]
- Hussain, H.A.; Roberts, A.P.; Mullany, P. Generation of an Erythromycin-Sensitive Derivative of Clostridium difficile Strain 630 (630ΔeErm) and Demonstration That the Conjugative Transposon Tn916ΔE Enters the Genome of This Strain at Multiple Sites. J. Med. Microbiol. 2005, 54, 137–141. [Google Scholar] [CrossRef] [PubMed]
- Saujet, L.; Monot, M.; Dupuy, B.; Soutourina, O.; Martin-Verstraete, I. The Key Sigma Factor of Transition Phase, SigH, Controls Sporulation, Metabolism, and Virulence Factor Expression in Clostridium difficile. J. Bacteriol. 2011, 193, 3186–3196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Eymann, C.; Dreisbach, A.; Albrecht, D.; Bernhardt, J.; Becher, D.; Gentner, S.; Tam, L.T.; Büttner, K.; Buurman, G.; Scharf, C.; et al. A Comprehensive Proteome Map of Growing Bacillus subtilis Cells. Proteomics 2004, 4, 2849–2876. [Google Scholar] [CrossRef]
- Bonn, F.; Bartel, J.; Büttner, K.; Hecker, M.; Otto, A.; Becher, D. Picking Vanished Proteins from the Void: How to Collect and Ship/Share Extremely Dilute Proteins in a Reproducible and Highly Efficient Manner. Anal. Chem. 2014, 86, 7421–7427. [Google Scholar] [CrossRef] [PubMed]
- Vizcaíno, J.A.; Csordas, A.; del-Toro, N.; Dianes, J.A.; Griss, J.; Lavidas, I.; Mayer, G.; Perez-Riverol, Y.; Reisinger, F.; Ternent, T.; et al. 2016 Update of the PRIDE Database and Its Related Tools. Nucleic Acids Res. 2016, 44, D447–D456. [Google Scholar] [CrossRef] [PubMed]
- Cox, J.; Hein, M.Y.; Luber, C.A.; Paron, I.; Nagaraj, N.; Mann, M. Accurate Proteome-Wide Label-Free Quantification by Delayed Normalization and Maximal Peptide Ratio Extraction, Termed Maxlfq. Mol. Cell. Proteom. 2014, 13, 2513–2526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dannheim, H.; Riedel, T.; Neumann-Schaal, M.; Bunk, B.; Schober, I.; Spröer, C.; Chibani, C.M.; Gronow, S.; Liesegang, H.; Overmann, J.; et al. Manual Curation and Reannotation of the Genomes of Clostridium difficile 630Δerm and Clostridium difficile 630. J. Med. Microbiol. 2017, 66, 286–293. [Google Scholar] [CrossRef]
- Schwanhäusser, B.; Busse, D.; Li, N.; Dittmar, G.; Schuchhardt, J.; Wolf, J.; Chen, W.; Selbach, M. Global Quantification of Mammalian Gene Expression Control. Nature 2011, 473, 337–342. [Google Scholar] [CrossRef] [Green Version]
- Tyanova, S.; Cox, J. Perseus: A bioinformatics platform for integrative analysis of proteomics data in cancer research. In Cancer Systems Biology: Methods and Protocols; von Stechow, L., Ed.; Springer: New York, NY, USA, 2018; pp. 133–148. [Google Scholar]
- Sievers, S.; Metzendorf, N.G.; Dittmann, S.; Troitzsch, D.; Gast, V.; Tröger, S.M.; Wolff, C.; Zühlke, D.; Hirschfeld, C.; Schlüter, R.; et al. Differential View on the Bile Acid Stress Response of Clostridioides difficile. Front. Microbiol. 2019, 10, 258. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Cherian, P.T.; Lee, R.E.; Hurdle, J.G. The Membrane as a Target for Controlling Hypervirulent Clostridium difficile Infections. J. Antimicrob. Chemother. 2013, 68, 806–815. [Google Scholar] [CrossRef] [PubMed]
- Muntel, J.; Fromion, V.; Goelzer, A.; Maaβ, S.; Mäder, U.; Büttner, K.; Hecker, M.; Becher, D. Comprehensive Absolute Quantification of the Cytosolic Proteome of Bacillus subtilis by Data Independent, Parallel Fragmentation in Liquid Chromatography/Mass Spectrometry (LC/MSE). Mol. Cell. Proteom. MCP 2014, 13, 1008–1019. [Google Scholar] [CrossRef] [Green Version]
- Goelzer, A.; Muntel, J.; Chubukov, V.; Jules, M.; Prestel, E.; Nölker, R.; Mariadassou, M.; Aymerich, S.; Hecker, M.; Noirot, P.; et al. Quantitative Prediction of Genome-Wide Resource Allocation in Bacteria. Metab. Eng. 2015, 32, 232–243. [Google Scholar] [CrossRef] [PubMed]
- Okusu, H.; Ma, D.; Nikaido, H. AcrAB Efflux Pump Plays a Major Role in the Antibiotic Resistance Phenotype of Escherichia coli Multiple-Antibiotic-Resistance (mar) Mutants. J. Bacteriol. 1996, 178, 306–308. [Google Scholar] [CrossRef] [Green Version]
- Goldman, J.D.; White, D.G.; Levy, S.B. Multiple Antibiotic Resistance (mar) Locus Protects Escherichia coli from Rapid Cell Killing by Fluoroquinolones. Antimicrob. Agents Chemother. 1996, 40, 1266–1269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, J.; Deng, W.; Yang, W.; Luo, H.; Duan, X.; Xie, L.; Li, P.; Wang, R.; Fu, T.; Abdalla, A.E.; et al. Mycobacterium tuberculosis Rv1152 Is a Novel GntR Family Transcriptional Regulator Involved in Intrinsic Vancomycin Resistance and Is a Potential Vancomycin Adjuvant Target. Sci. Rep. 2016, 6, 28002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, J.; Zhao, L.; Yang, M. A GntR Family Transcription Factor Positively Regulates Mycobacterial Isoniazid Resistance by Controlling the Expression of a Putative Permease. BMC Microbiol. 2015, 15, 214. [Google Scholar] [CrossRef] [Green Version]
- Maaß, S.; Otto, A.; Albrecht, D.; Riedel, K.; Trautwein-Schult, A.; Becher, D. Proteomic Signatures of Clostridium difficile Stressed with Metronidazole, Vancomycin, or Fidaxomicin. Cells 2018, 7, 213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nie, X.; Yang, B.; Zhang, L.; Gu, Y.; Yang, S.; Jiang, W.; Yang, C. PTS Regulation Domain-Containing Transcriptional Activator CelR and Sigma Factor σ54 Control Cellobiose Utilization in Clostridium acetobutylicum. Mol. Microbiol. 2016, 100, 289–302. [Google Scholar] [CrossRef] [Green Version]
- Gaupp, R.; Wirf, J.; Wonnenberg, B.; Biegel, T.; Eisenbeis, J.; Graham, J.; Herrmann, M.; Lee, C.Y.; Beisswenger, C.; Wolz, C.; et al. RpiRc Is a Pleiotropic Effector of Virulence Determinant Synthesis and Attenuates Pathogenicity in Staphylococcus Aureus. Infect. Immun. 2016, 84, 2031–2041. [Google Scholar] [CrossRef] [Green Version]
- Ganeshapillai, J.; Vinogradov, E.; Rousseau, J.; Weese, J.S.; Monteiro, M.A. Clostridium difficile Cell-Surface Polysaccharides Composed of Pentaglycosyl and Hexaglycosyl Phosphate Repeating Units. Carbohydr. Res. 2008, 343, 703–710. [Google Scholar] [CrossRef] [PubMed]
- Jarrad, A.M.; Blaskovich, M.A.T.; Prasetyoputri, A.; Karoli, T.; Hansford, K.A.; Cooper, M.A. Detection and Investigation of Eagle Effect Resistance to Vancomycin in Clostridium difficile with an ATP-Bioluminescence Assay. Front. Microbiol. 2018, 9, 1420. [Google Scholar] [CrossRef] [PubMed]
- Steil, L.; Hoffmann, T.; Budde, I.; Völker, U.; Bremer, E. Genome-Wide Transcriptional Profiling Analysis of Adaptation of Bacillus subtilis to High Salinity. J. Bacteriol. 2003, 185, 6358–6370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hecker, M.; Pané-Farré, J.; Völker, U. SigB-Dependent General Stress Response in Bacillus subtilis and Related Gram-Positive Bacteria. Annu. Rev. Microbiol. 2007, 61, 215–236. [Google Scholar] [CrossRef]
- Hecker, M.; Reder, A.; Fuchs, S.; Pagels, M.; Engelmann, S. Physiological Proteomics and Stress/Starvation Responses in Bacillus subtilis and Staphylococcus aureus. Res. Microbiol. 2009, 160, 245–258. [Google Scholar] [CrossRef] [PubMed]
- Maaß, S.; Wachlin, G.; Bernhardt, J.; Eymann, C.; Fromion, V.; Riedel, K.; Becher, D.; Hecker, M. Highly Precise Quantification of Protein Molecules per Cell during Stress and Starvation Responses in Bacillus subtilis. Mol. Cell. Proteom. 2014, 13, 2260–2276. [Google Scholar]
- Logan, S.M. Flagellar Glycosylation—A New Component of the Motility Repertoire? Microbiol. Soc. 2006, 152, 1249–1262. [Google Scholar] [CrossRef] [Green Version]
- Twine, S.M.; Reid, C.W.; Aubry, A.; McMullin, D.R.; Fulton, K.M.; Austin, J.; Logan, S.M. Motility and Flagellar Glycosylation in Clostridium difficile. J. Bacteriol. 2009, 191, 7050–7062. [Google Scholar] [CrossRef] [Green Version]
- Duan, Q.; Zhou, M.; Zhu, L.; Zhu, G. Flagella and Bacterial Pathogenicity. J. Basic Microbiol. 2013, 53, 1–8. [Google Scholar] [CrossRef]
- Aubry, A.; Hussack, G.; Chen, W.; KuoLee, R.; Twine, S.M.; Fulton, K.M.; Foote, S.; Carrillo, C.D.; Tanha, J.; Logan, S.M. Modulation of Toxin Production by the Flagellar Regulon in Clostridium difficile. Infect. Immun. 2012, 80, 3521–3532. [Google Scholar] [CrossRef] [Green Version]
- Tasteyre, A.; Barc, M.-C.; Collignon, A.; Boureau, H.; Karjalainen, T. Role of FliC and FliD Flagellar Proteins of Clostridium difficile in Adherence and Gut Colonization. Infect. Immun. 2001, 69, 7937–7940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dingle, T.C.; Mulvey, G.L.; Armstrong, G.D. Mutagenic Analysis of the Clostridium difficile Flagellar Proteins, FliC and FliD, and Their Contribution to Virulence in Hamsters. Infect. Immun. 2011, 79, 4061–4067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baban, S.T.; Kuehne, S.A.; Barketi-Klai, A.; Cartman, S.T.; Kelly, M.L.; Hardie, K.R.; Kansau, I.; Collignon, A.; Minton, N.P. The Role of Flagella in Clostridium difficile Pathogenesis: Comparison between a Non-Epidemic and an Epidemic Strain. PLoS ONE 2013, 8, e73026. [Google Scholar] [CrossRef]
- Breukink, E.; Wiedemann, I.; van Kraaij, C.; Kuipers, O.P.; Sahl, H.-G.; Kruijff, B. de Use of the Cell Wall Precursor Lipid II by a Pore-Forming Peptide Antibiotic. Science 1999, 286, 2361–2364. [Google Scholar] [CrossRef] [Green Version]
- Van Heusden, H.E.; de Kruijff, B.; Breukink, E. Lipid II Induces a Transmembrane Orientation of the Pore-Forming Peptide Lantibiotic Nisin. Biochemistry 2002, 41, 12171–12178. [Google Scholar] [CrossRef]
- Breukink, E.; van Heusden, H.E.; Vollmerhaus, P.J.; Swiezewska, E.; Brunner, L.; Walker, S.; Heck, A.J.R.; de Kruijff, B. Lipid II Is an Intrinsic Component of the Pore Induced by Nisin in Bacterial Membranes. J. Biol. Chem. 2003, 278, 19898–19903. [Google Scholar] [CrossRef] [Green Version]
- Stogios, P.J.; Savchenko, A. Molecular Mechanisms of Vancomycin Resistance. Protein Sci. Publ. Protein Soc. 2020, 29, 654–669. [Google Scholar] [CrossRef]
- Hessling, B.; Bonn, F.; Otto, A.; Herbst, F.-A.; Rappen, G.-M.; Bernhardt, J.; Hecker, M.; Becher, D. Global Proteome Analysis of Vancomycin Stress in Staphylococcus aureus. Int. J. Med. Microbiol. 2013, 303, 624–634. [Google Scholar] [CrossRef]
- Leeds, J.A.; Sachdeva, M.; Mullin, S.; Barnes, S.W.; Ruzin, A. In Vitro Selection, via Serial Passage, of Clostridium difficile Mutants with Reduced Susceptibility to Fidaxomicin or Vancomycin. J. Antimicrob. Chemother. 2014, 69, 41–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hurdle, J.G.; O’Neill, A.J.; Chopra, I.; Lee, R.E. Targeting Bacterial Membrane Function: An Underexploited Mechanism for Treating Persistent Infections. Nat. Rev. Microbiol. 2011, 9, 62–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belley, A.; Neesham-Grenon, E.; McKay, G.; Arhin, F.F.; Harris, R.; Beveridge, T.; Parr, T.R.; Moeck, G. Oritavancin Kills Stationary-Phase and Biofilm Staphylococcus aureus Cells in Vitro. Antimicrob. Agents Chemother. 2009, 53, 918–925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silverman, J.A.; Perlmutter, N.G.; Shapiro, H.M. Correlation of Daptomycin Bactericidal Activity and Membrane Depolarization in Staphylococcus aureus. Antimicrob. Agents Chemother. 2003, 47, 2538–2544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, H.; Huynh, Q.; Bärlehner, D.; Heerklotz, H. Additive and Synergistic Membrane Permeabilization by Antimicrobial (Lipo)Peptides and Detergents. Biophys. J. 2014, 106, 2115–2125. [Google Scholar] [CrossRef] [PubMed] [Green Version]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maaß, S.; Bartel, J.; Mücke, P.-A.; Schlüter, R.; Sura, T.; Zaschke-Kriesche, J.; Smits, S.H.J.; Becher, D. Proteomic Adaptation of Clostridioides difficile to Treatment with the Antimicrobial Peptide Nisin. Cells 2021, 10, 372. https://doi.org/10.3390/cells10020372
Maaß S, Bartel J, Mücke P-A, Schlüter R, Sura T, Zaschke-Kriesche J, Smits SHJ, Becher D. Proteomic Adaptation of Clostridioides difficile to Treatment with the Antimicrobial Peptide Nisin. Cells. 2021; 10(2):372. https://doi.org/10.3390/cells10020372
Chicago/Turabian StyleMaaß, Sandra, Jürgen Bartel, Pierre-Alexander Mücke, Rabea Schlüter, Thomas Sura, Julia Zaschke-Kriesche, Sander H. J. Smits, and Dörte Becher. 2021. "Proteomic Adaptation of Clostridioides difficile to Treatment with the Antimicrobial Peptide Nisin" Cells 10, no. 2: 372. https://doi.org/10.3390/cells10020372
APA StyleMaaß, S., Bartel, J., Mücke, P. -A., Schlüter, R., Sura, T., Zaschke-Kriesche, J., Smits, S. H. J., & Becher, D. (2021). Proteomic Adaptation of Clostridioides difficile to Treatment with the Antimicrobial Peptide Nisin. Cells, 10(2), 372. https://doi.org/10.3390/cells10020372