
Association between Oncostatin M Expression and Inflammatory Phenotype in Experimental Arthritis Models and Osteoarthritis Patients

 , , and
, , and 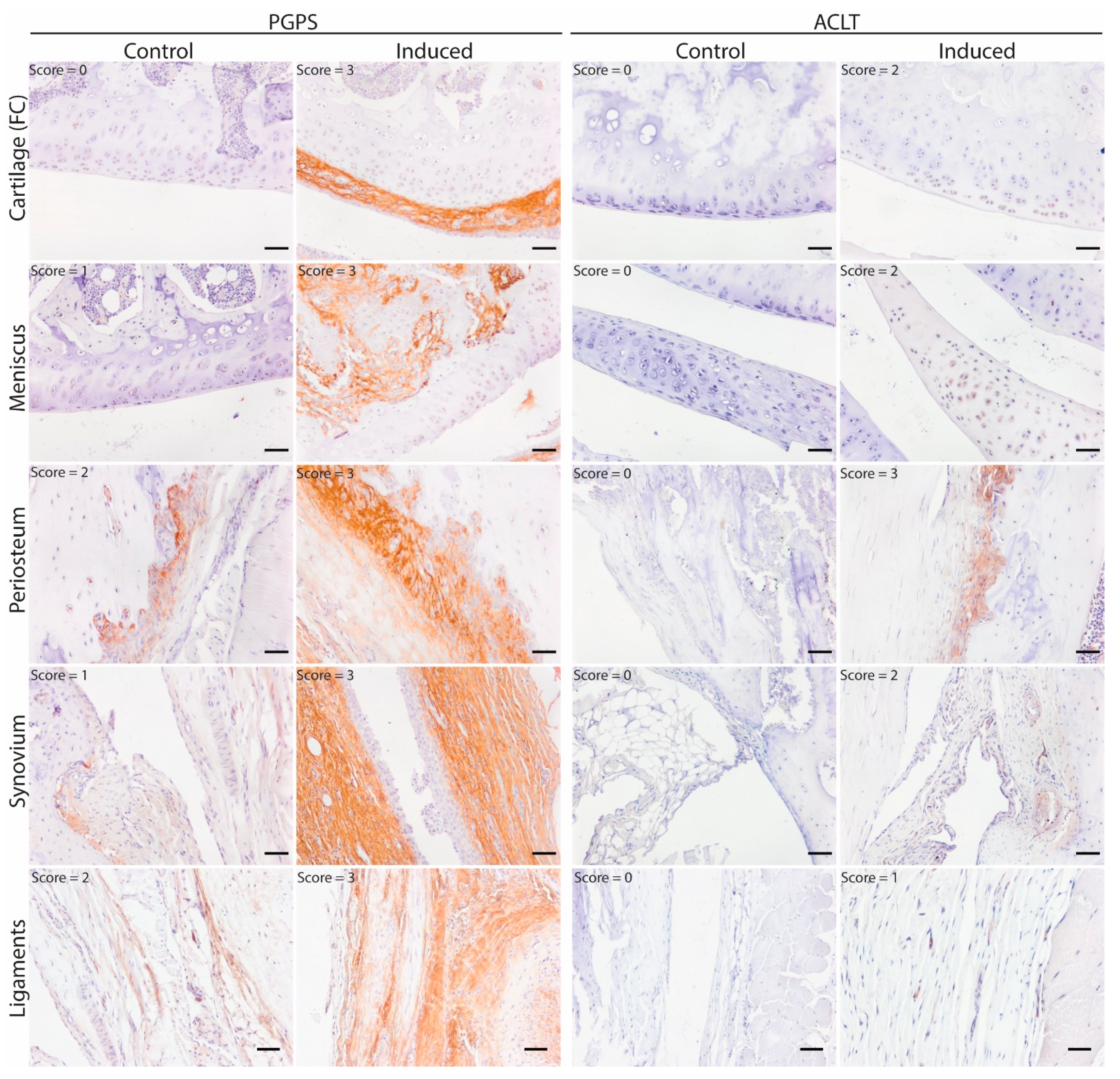{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
2.1. Synovial Fluid Collection and Cytokine Measurements
2.2. Animal Models
2.3. Histology
2.4. OSM Immunohistochemistry
2.5. Micro-CT Analysis
2.6. Statistics
3. Results
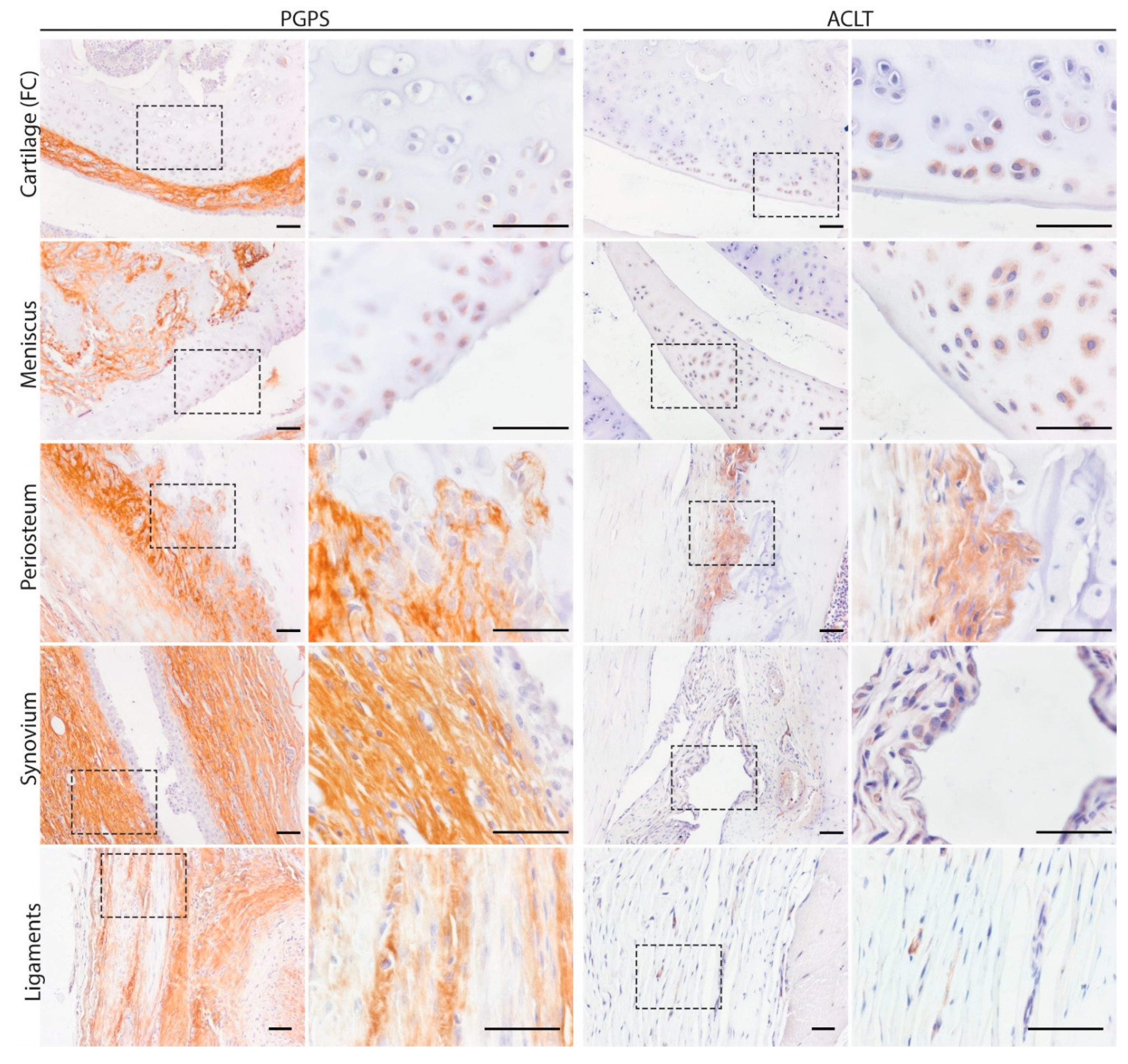
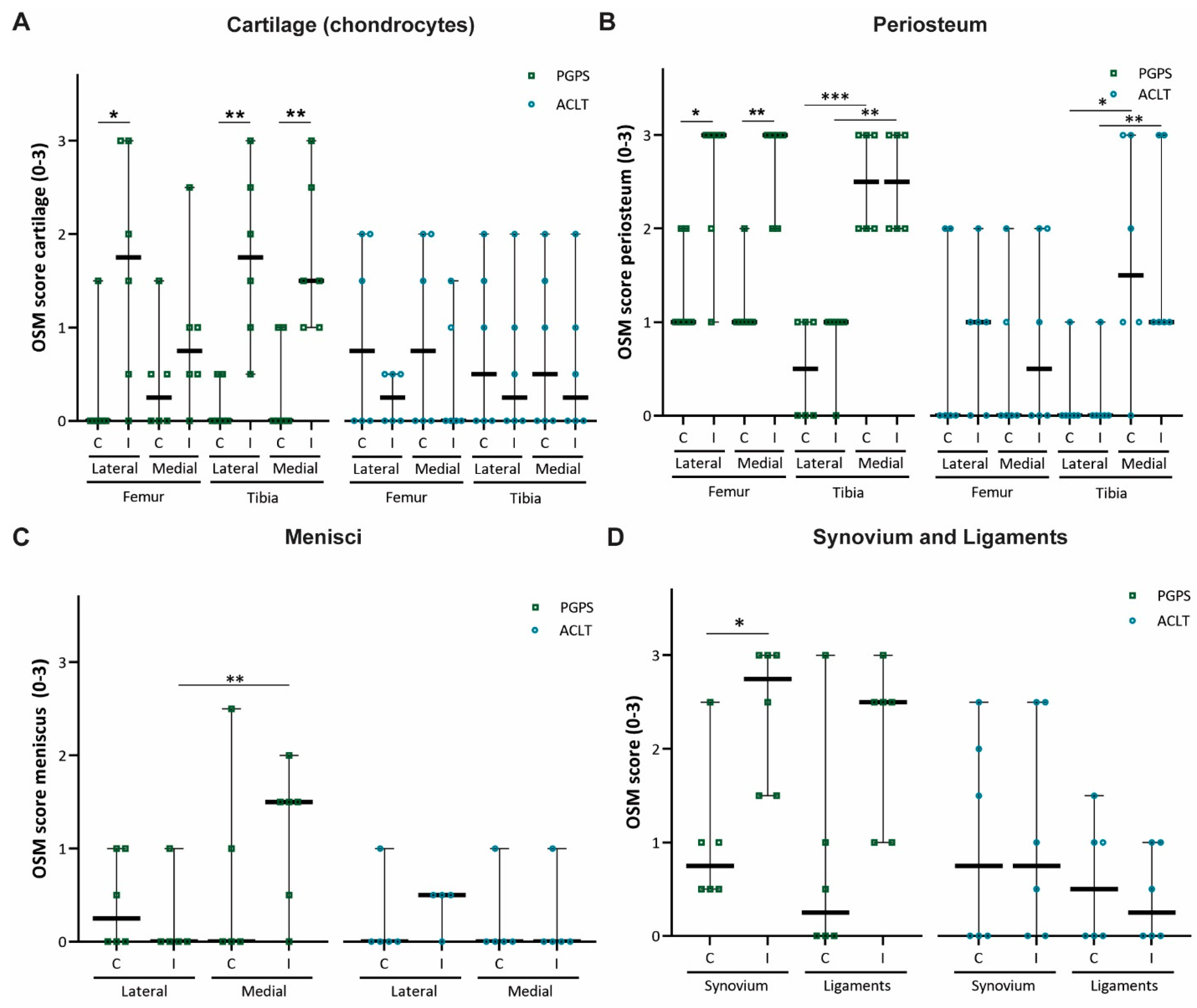
3.1. Mapping and Quantification of OSM Expression in Joint Tissues
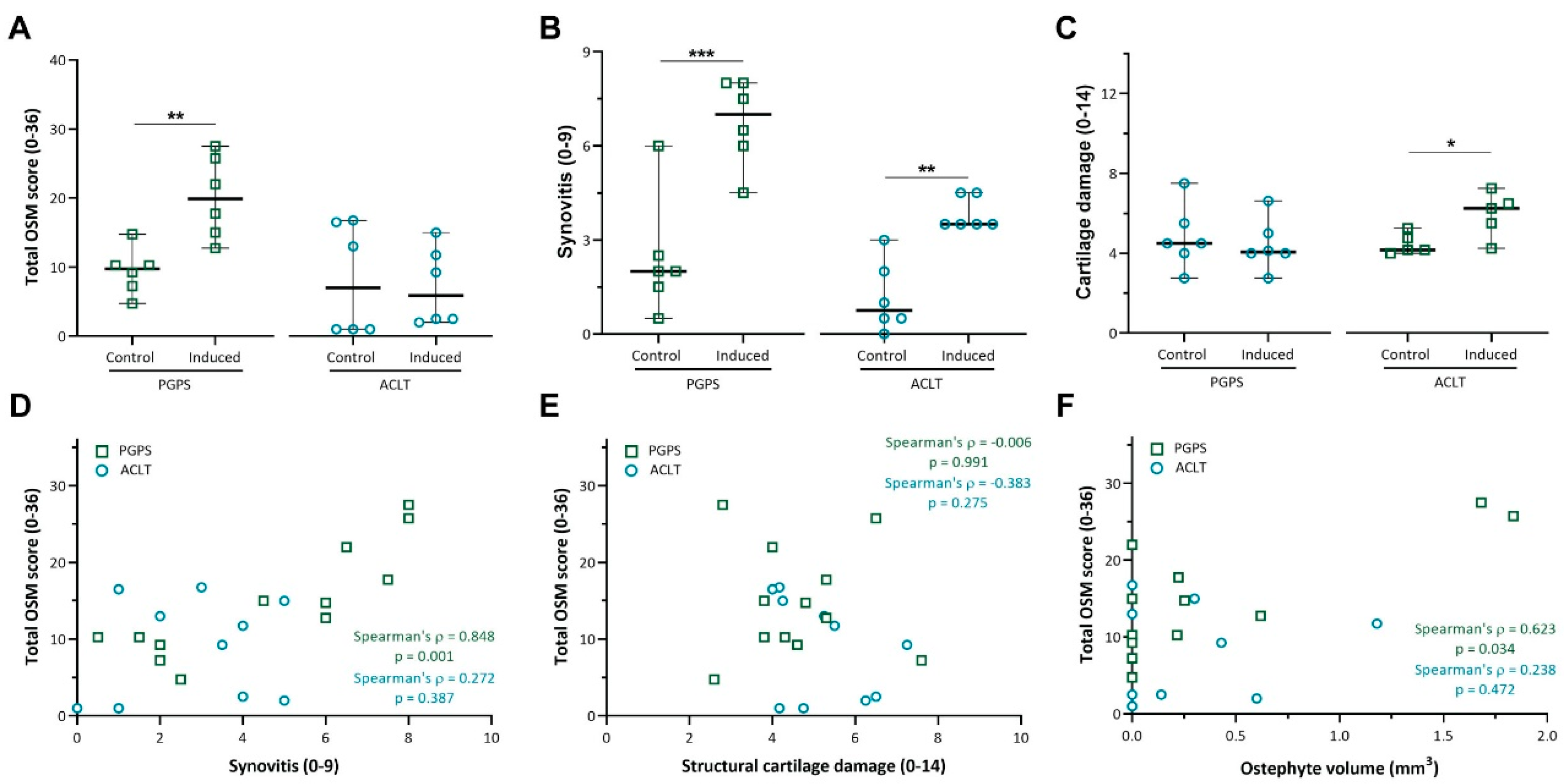
3.2. Expression of OSM Correlates with Synovitis in the PGPS Model of Acute Arthritis
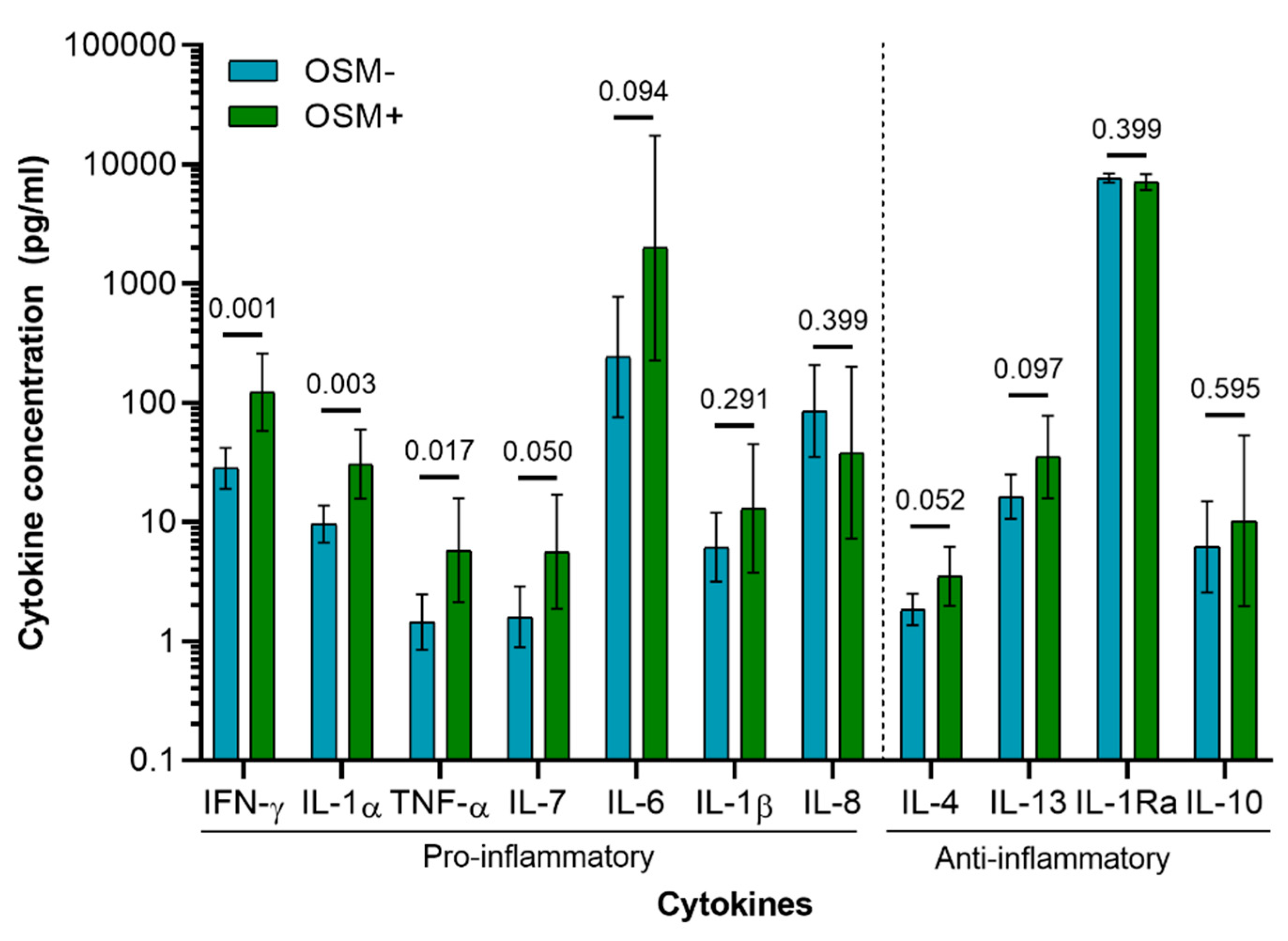
3.3. Cytokine Profile of Osteoarthritis Patients
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Woolf, A.D.; Pfleger, B. Burden of major musculoskeletal conditions. Bull. World Health Organ. 2003, 81, 646–656. [Google Scholar] [PubMed]
- Goldring, M.B.; Goldring, S.R. Osteoarthritis. J. Cell. Physiol. 2007, 213, 626–634. [Google Scholar] [CrossRef]
- Goldring, M.B.; Otero, M. Inflammation in osteoarthritis. Curr. Opin. Rheumatol. 2011, 23, 471–478. [Google Scholar] [CrossRef]
- Bijlsma, J.W.; Berenbaum, F.; Lafeber, F.P. Osteoarthritis: An update with relevance for clinical practice. Lancet 2011, 377, 2115–2126. [Google Scholar] [CrossRef]
- Schue, J.R.; Tawfik, O.; Smith, D.D.; Hinson, G.; Bolce, R.; Wick, J.A.; Lindsley, H.B. Treatment of knee osteoarthritis with intraarticular infliximab may improve knee function and reduce synovial infiltration by macrophages. In Proceedings of the Annual Scientific Meeting of the American College of Rheumatology and Association of Rheumatology Health Professionals, Chicago, IL, USA, 4 November 2011. [Google Scholar]
- Chevalier, X.; Goupille, P.; Beaulieu, A.D.; Burch, F.X.; Bensen, W.G.; Conrozier, T.; Loeuille, D.; Kivitz, A.J.; Silver, D.; Appleton, B.E. Intraarticular injection of anakinra in osteoarthritis of the knee: A multicenter, randomized, double-blind, placebo-controlled study. Arthritis Rheumatol. 2009, 61, 344–352. [Google Scholar] [CrossRef]
- Fleischmann, R.M.; Bliddal, H.; Blanco, F.J.; Schnitzer, T.J.; Peterfy, C.; Chen, S.; Wang, L.; Feng, S.; Conaghan, P.G.; Berenbaum, F.; et al. A phase II trial of lutikizumab, an anti–interleukin-1α/β dual variable domain immunoglobulin, in knee osteoarthritis patients with synovitis. Arthritis Rheumatol. 2019, 71, 1056–1069. [Google Scholar] [CrossRef] [PubMed]
- Richette, P.; François, M.; Vicaut, E.; Fitting, C.; Bardin, T.; Corvol, M.; Savouret, J.-F.; Rannou, F. A high interleukin 1 receptor antagonist/IL-1beta ratio occurs naturally in knee osteoarthritis. J. Rheumatol. 2008, 35, 1650–1654. [Google Scholar]
- Simão, A.P.; Almeida, T.M.D.O.; Mendonça, V.A.; Santos, S.A.; Gomes, W.F.; Coimbra, C.C.; Lacerda, A.C.R. Soluble TNF receptors are produced at sites of inflammation and are inversely associated with self-reported symptoms (WOMAC) in knee osteoarthritis. Rheumatol. Int. 2014, 34, 1759–1763. [Google Scholar] [CrossRef]
- Hui, W.; Bell, M.; Carroll, G. Detection of oncostatin M in synovial fluid from patients with rheumatoid arthritis. Ann. Rheum. Dis. 1997, 56, 184–187. [Google Scholar] [CrossRef] [Green Version]
- Cawston, T.E.; Curry, V.A.; Summers, C.A.; Clark, I.M.; Riley, G.P.; Life, P.F.; Spaull, J.R.; Goldring, M.B.; Koshy, P.J.T.; Rowan, A.D.; et al. The role of oncostatin M in animal and human connective tissue collagen turnover and its localization within the rheumatoid joint. Arthritis Rheum. 1998, 41, 1760–1771. [Google Scholar] [CrossRef]
- Fearon, U.; Mullan, R.; Markham, T.; Connolly, M.; Sullivan, S.; Poole, A.R.; Fitzgerald, O.; Bresnihan, B.; Veale, D.J. Oncostatin M induces angiogenesis and cartilage degradation in rheumatoid arthritis synovial tissue and human cartilage cocultures. Arthritis Rheum. 2006, 54, 3152–3162. [Google Scholar] [CrossRef]
- Hui, W.; E Cawston, T.; Richards, C.D.; Rowan, A.D. A model of inflammatory arthritis highlights a role for oncostatin M in pro-inflammatory cytokine-induced bone destruction via RANK/RANKL. Arthritis Res. Ther. 2005, 7, R57–R64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durigova, M.; Roughley, P.; Mort, J. Mechanism of proteoglycan aggregate degradation in cartilage stimulated with oncostatin M. Osteoarthr. Cartil. 2008, 16, 98–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richards, C.D. The enigmatic cytokine oncostatin M and roles in disease. ISRN Inflamm. 2013, 2013, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Rowan, A.D.; Hui, W.; Cawston, T.E.; Richards, C.D. Adenoviral gene transfer of interleukin-1 in combination with oncostatin M induces significant joint damage in a murine model. Am. J. Pathol. 2003, 162, 1975–1984. [Google Scholar] [CrossRef] [Green Version]
- Hui, W.; Rowan, A.D.; Richards, C.D.; Cawston, T.E. Oncostatin M in combination with tumor necrosis factor? induces cartilage damage and matrix metalloproteinase expression in vitro and in vivo. Arthritis Rheum. 2003, 48, 3404–3418. [Google Scholar] [CrossRef]
- Manicourt, D.-H.; Poilvache, P.; Egeren, A.V.; Devogelaer, J.-P.; Lenz, M.-E.; Thonar, E.J.M.A. Synovial fluid levels of tumor necrosis factor α and oncostatin M correlate with levels of markers of the degradation of crosslinked collagen and cartilage aggrecan in rheumatoid arthritis but not in osteoarthritis. Arthritis Rheum. 2000, 43, 281–288. [Google Scholar] [CrossRef]
- Beekhuizen, M.; Van Osch, G.; Bot, A.; Hoekstra, M.; Saris, D.; Dhert, W.; Creemers, L. Inhibition of oncostatin M in osteoarthritic synovial fluid enhances GAG production in osteoarthritic cartilage repair. Eur. Cells Mater. 2013, 26, 80–90. [Google Scholar] [CrossRef]
- Tsuchida, A.I.; Beekhuizen, M.; Hart, M.C.; Radstake, T.R.D.J.; Dhert, W.J.A.; Saris, D.B.F.; Van Osch, G.J.V.M.; Creemers, L.B. Cytokine profiles in the joint depend on pathology, but are different between synovial fluid, cartilage tissue and cultured chondrocytes. Arthritis Res. Ther. 2014, 16, 441. [Google Scholar] [CrossRef] [Green Version]
- Heymann, D.; Rousselle, A.V. gp130 Cytokine family and bone cells. Cytokine 2000, 12, 1455–1468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Hooge, A.S.; van de Loo, F.A.; Bennink, M.B.; de Jong, D.S.; Arntz, O.J.; Lubberts, E.; Richards, C.D.; vandDen Berg, W.B. Adenoviral transfer of murine oncostatin M elicits periosteal bone apposition in knee joints of mice, despite synovial inflammation and up-regulated expression of interleukin-6 and receptor activator of nuclear factor-kappa B ligand. Am. J. Pathol. 2002, 160, 1733–1743. [Google Scholar]
- Walker, E.C.; McGregor, N.E.; Poulton, I.J.; Solano, M.; Pompolo, S.; Fernandes, T.J.; Constable, M.J.; Nicholson, G.C.; Zhang, J.-G.; Nicola, N.A.; et al. Oncostatin M promotes bone formation independently of resorption when signaling through leukemia inhibitory factor receptor in mice. J. Clin. Investig. 2010, 120, 582–592. [Google Scholar] [CrossRef] [Green Version]
- Cross, A.; Edwards, S.W.; Bucknall, R.C.; Moots, R.J. Secretion of oncostatin M by neutrophils in rheumatoid arthritis. Arthritis Rheum. 2004, 50, 1430–1436. [Google Scholar] [CrossRef]
- Brown, T.J.; Lioubin, M.N.; Marquardt, H. Purification and characterization of cytostatic lymphokines produced by activated human T lymphocytes. Synergistic antiproliferative activity of transforming growth factor beta 1, interferon-gamma, and oncostatin M for human melanoma cells. J. Immunol. 1987, 139, 2977–2983. [Google Scholar]
- Salamon, P.; Shoham, N.G.; Puxeddu, I.; Paitan, Y.; Levi-Schaffer, F.; Mekori, Y.A. Human mast cells release oncostatin M on contact with activated T cells: Possible biologic relevance. J. Allergy Clin. Immunol. 2008, 121, 448–455.e5. [Google Scholar] [CrossRef] [PubMed]
- De Lange-Brokaar, B.J.E.; Ioan-Facsinay, A.; van Osch, G.J.V.M.; Zuurmond, A.M.; Schoones, J.; Toes, R.E.M.; Huizinga, T.W.J.; Kloppenburg, M. Synovial inflammation, immune cells and their cytokines in osteoarthritis: A review. Osteoarthr. Cartil. 2012, 20, 1484–1499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cromartie, W.J.; Craddock, J.G.; Schwab, J.H.; Anderle, S.K.; Yang, C.H. Arthritis in rats after systemic injection of streptococcal cells or cell walls. J. Exp. Med. 1977, 146, 1585–1602. [Google Scholar] [CrossRef] [Green Version]
- Lampropoulou-Adamidou, K.; Lelovas, P.; Karadimas, E.V.; Liakou, C.; Triantafillopoulos, I.K.; Dontas, I.; Papaioannou, N.A. Useful animal models for the research of osteoarthritis. Eur. J. Orthop. Surg. Traumatol. 2014, 24, 263–271. [Google Scholar] [CrossRef]
- Rudnik-Jansen, I.; Schrijver, K.; Woike, N.; Tellegen, A.; Versteeg, S.; Emans, P.; Mihov, G.; Thies, J.; Eijkelkamp, N.; Tryfonidou, M.; et al. Intra-articular injection of triamcinolone acetonide releasing biomaterial microspheres inhibits pain and inflammation in an acute arthritis model. Drug Deliv. 2019, 26, 226–236. [Google Scholar] [CrossRef] [Green Version]
- Rudnik-Jansen, I.; Tellegen, A.R.; Pouran, B.; Schrijver, K.; Meij, B.P.; Emans, P.J.; De Gendt, E.; Thomas, R.E.; Kik, M.J.; De Visser, H.M.; et al. Local controlled release of corticosteroids extends surgically induced joint instability by inhibiting tissue healing. Br. J. Pharmacol. 2019, 176, 4050–4064. [Google Scholar] [CrossRef] [PubMed]
- Mankin, H.J.; Lippiello, L. Biochemical and metabolic abnormalities in articular cartilage from osteo-arthritic human hips. J. Bone Jt. Surg. Am. 1970, 52, 424–434. [Google Scholar] [CrossRef]
- Krenn, V.; Morawietz, L.; Burmester, G.-R.; Kinne, R.W.; Mueller-Ladner, U.; Muller, B.; Haupl, T. Synovitis score: Discrimination between chronic low-grade and high-grade synovitis. Histopathology 2006, 49, 358–364. [Google Scholar] [CrossRef] [PubMed]
- Koo, T.K.; Li, M.Y. A guideline of selecting and reporting intraclass correlation coefficients for reliability research. J. Chiropr. Med. 2016, 15, 155–163. [Google Scholar] [CrossRef] [Green Version]
- Roosa, S.M.M.; Liu, Y.; Turner, C.H. Gene expression patterns in bone following mechanical loading. J. Bone Min. Res. 2011, 26, 100–112. [Google Scholar] [CrossRef] [Green Version]
- Pasta, D.J.; Miriam, G. Cisternas: Estimating standard errors for class variables in generalized linear models using PROC IML. In Proceedings of the SAS Conference Proceedings: SAS Users Group International 28, Seattle, WA, USA, 30 March 2003. [Google Scholar]
- Wright, H.L.; Moots, R.J.; Edwards, S.W. The multifactorial role of neutrophils in rheumatoid arthritis. Nat. Rev. Rheumatol. 2014, 10, 593–601. [Google Scholar] [CrossRef]
- Taşoğlu, Ö.; Bölük, H.; Onat, Ş.Ş.; Taşoğlu, I.; Özgirgin, N. Is blood neutrophil-lymphocyte ratio an independent predictor of knee osteoarthritis severity? Clin. Rheumatol. 2016, 35, 1579–1583. [Google Scholar] [CrossRef] [PubMed]
- Walker, E.C.; Johnson, R.W.; Hu, Y.; Brennan, H.J.; Poulton, I.J.; Zhang, J.-G.; Jenkins, B.J.; Smyth, G.K.; Nicola, N.A.; Sims, N.A. Murine oncostatin M acts via leukemia inhibitory factor receptor to phosphorylate signal transducer and activator of transcription 3 (STAT3) but not STAT1, an effect that protects bone mass. J. Biol. Chem. 2016, 291, 21703–21716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldring, S.R. The effects of inflammatory arthritis on bone remodeling. Arthritis Res. Ther. 2005, 7, S3–S4. [Google Scholar] [CrossRef]
- Siebelt, M.; Groen, H.C.; Koelewijn, S.J.; De Blois, E.; Sandker, M.; Waarsing, J.H.; Müller, C.; Van Osch, G.J.; De Jong, M.; Weinans, H. Increased physical activity severely induces osteoarthritic changes in knee joints with papain induced sulfate-glycosaminoglycan depleted cartilage. Arthritis Res. Ther. 2014, 16, R32. [Google Scholar] [CrossRef] [Green Version]
- Moshtagh, P.R.; Korthagen, N.M.; Plomp, S.G.; Pouran, B.; Castelein, R.M.; Zadpoor, A.A.; Weinans, H. Early signs of bone and cartilage changes induced by treadmill exercise in rats. JBMR Plus 2018, 2, 134–142. [Google Scholar] [CrossRef] [Green Version]
- Van Lent, P.L.; Blom, A.B.; van der Kraan, P.; Holthuysen, A.E.; Vitters, E.; van Rooijen, N.; Smeets, R.L.; Nabbe, K.C.; van den Berg, W.B. Crucial role of synovial lining macrophages in the promotion of transforming growth factor beta-mediated osteophyte formation. Arthritis Rheum. 2004, 50, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Blom, A.B.; van Lent, P.L.; Holthuysen, A.E.; van der Kraan, P.M.; Roth, J.; van Rooijen, N.; van den Berg, W.B. Synovial lining macrophages mediate osteophyte formation during experimental osteoarthritis. Osteoarthr. Cartil. 2004, 12, 627–635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rudnik-Jansen, I.; Woike, N.; De Jong, S.; Versteeg, S.; Kik, M.; Emans, P.; Mihov, G.; Thies, J.; Eijkelkamp, N.; Tryfonidou, M.; et al. Applicability of a modified rat model of acute arthritis for long-term testing of drug delivery systems. Pharmaceutics 2019, 11, 70. [Google Scholar] [CrossRef] [Green Version]
- Murata, K.; Kanemura, N.; Kokubun, T.; Fujino, T.; Morishita, Y.; Onitsuka, K.; Fujiwara, S.; Nakajima, A.; Shimizu, D.; Takayanagi, K. Controlling joint instability delays the degeneration of articular cartilage in a rat model. Osteoarthr. Cartil. 2017, 25, 297–308. [Google Scholar] [CrossRef] [Green Version]
- Bigoni, M.; Sacerdote, P.; Turati, M.; Franchi, S.; Gandolla, M.; Gaddi, D.; Moretti, S.; Munegato, D.; Augusti, C.A.; Bresciani, E.; et al. Acute and late changes in intraarticular cytokine levels following anterior cruciate ligament injury. J. Orthop. Res. 2012, 31, 315–321. [Google Scholar] [CrossRef]
- Marks, P.H.; Donaldson, M.L.C. Inflammatory cytokine profiles associated with chondral damage in the anterior cruciate ligament–deficient knee. Arthrosc. J. Arthrosc. Relat. Surg. 2005, 21, 1342–1347. [Google Scholar] [CrossRef] [PubMed]
- Kraus, V.B.; Birmingham, J.; Stabler, T.V.; Feng, S.; Taylor, D.C.; Moorman, C.T.; Garrett, W.E.; Toth, A.P. Effects of intraarticular IL1-Ra for acute anterior cruciate ligament knee injury: A randomized controlled pilot trial (NCT00332254). Osteoarthr. Cartil. 2012, 20, 271–278. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garcia, J.P.; Utomo, L.; Rudnik-Jansen, I.; Du, J.; Zuithoff, N.P.A.; Krouwels, A.; van Osch, G.J.V.M.; Creemers, L.B. Association between Oncostatin M Expression and Inflammatory Phenotype in Experimental Arthritis Models and Osteoarthritis Patients. Cells 2021, 10, 508. https://doi.org/10.3390/cells10030508
Garcia JP, Utomo L, Rudnik-Jansen I, Du J, Zuithoff NPA, Krouwels A, van Osch GJVM, Creemers LB. Association between Oncostatin M Expression and Inflammatory Phenotype in Experimental Arthritis Models and Osteoarthritis Patients. Cells. 2021; 10(3):508. https://doi.org/10.3390/cells10030508
Chicago/Turabian StyleGarcia, Joao Pedro, Lizette Utomo, Imke Rudnik-Jansen, Jie Du, Nicolaas P.A. Zuithoff, Anita Krouwels, Gerjo J.V.M. van Osch, and Laura B. Creemers. 2021. "Association between Oncostatin M Expression and Inflammatory Phenotype in Experimental Arthritis Models and Osteoarthritis Patients" Cells 10, no. 3: 508. https://doi.org/10.3390/cells10030508
APA StyleGarcia, J. P., Utomo, L., Rudnik-Jansen, I., Du, J., Zuithoff, N. P. A., Krouwels, A., van Osch, G. J. V. M., & Creemers, L. B. (2021). Association between Oncostatin M Expression and Inflammatory Phenotype in Experimental Arthritis Models and Osteoarthritis Patients. Cells, 10(3), 508. https://doi.org/10.3390/cells10030508