
Morpho-Functional Consequences of Swiss Cheese Knockdown in Glia of Drosophila melanogaster


 , , and
, , and
Abstract
:
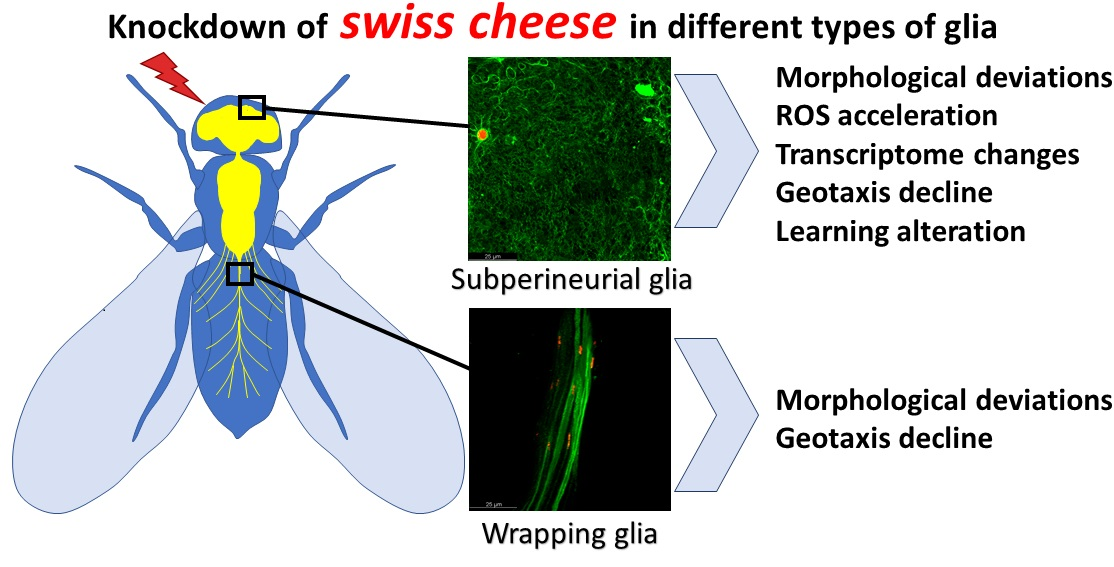{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Drosophila Stocks
2.2. Nervous System Preparation for Confocal Microscopy
2.3. Imaging and Analysis
2.4. Immunohistochemistry
2.5. Survival Analysis
2.6. Preparation and Analysis of Paraffin Sections
2.7. Negative Geotaxis Assay
2.8. Learning Indices Assessment
2.9. Oxidative Particle Measurement
2.10. Oxidized Glutathione Relative Level Assessment
2.11. Transcriptomic Assay
2.12. Results Processing
3. Results
3.1. sws is Expressed in the Central and Peripheral Nervous System in Drosophila Larva and Imago
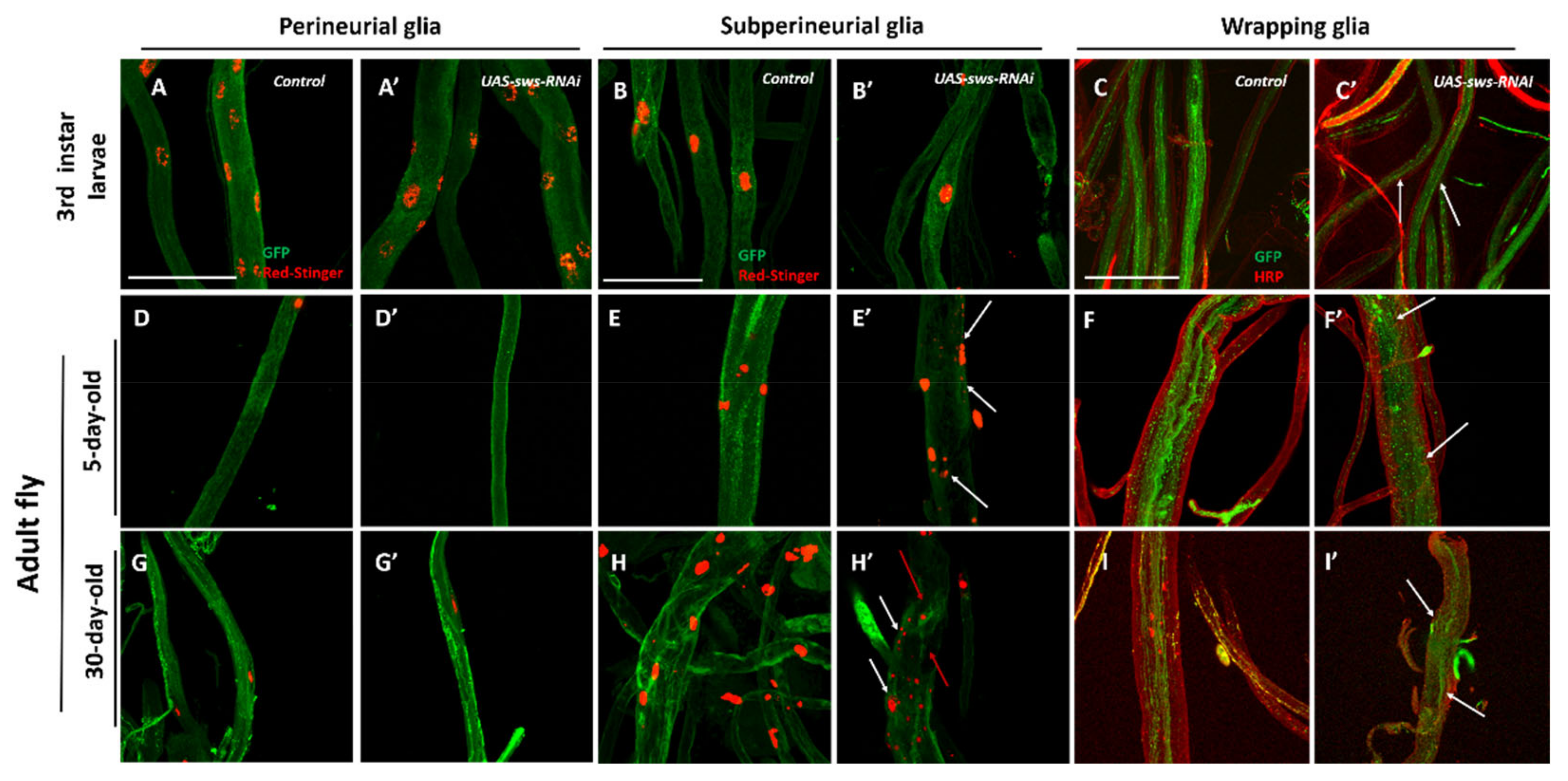
3.2. The sws Gene is Important for the Structural Maintenance of SPG in CNS
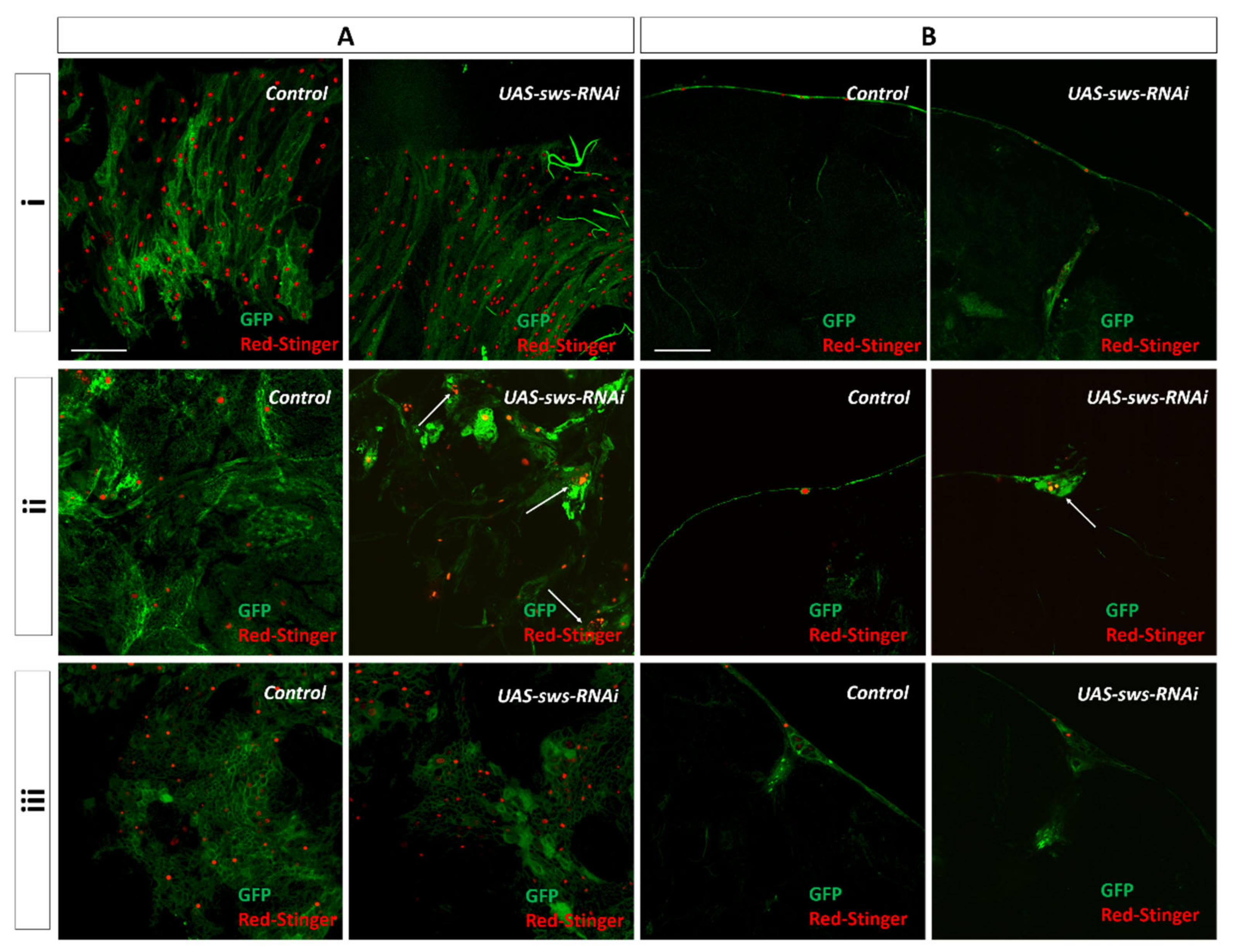
3.3. The sws Gene is Important for the Structural Maintenance of SPG and WG in PNS
3.4. Knockdown of sws in Glia Leads to Degeneration in CNS
3.5. Glial Knockdown of sws is Associated with Impaired Behavior
3.6. Whole-Organism Transcriptome Analysis of Flies with sws KD
3.7. Glial Knockdown of sws is Associated with Increased ROS Levels
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Moser, M.; Stempfl, T.; Li, Y.; Glynn, P.; Büttner, R.; Kretzschmar, D. Cloning and Expression of the Murine Sws/NTE Gene. Mech. Dev. 2000, 90, 279–282. [Google Scholar] [CrossRef]
- Johnson, M.K. The Delayed Neurotoxic Effect of Some Organophosphorus Compounds. Identification of the Phosphorylation Site as an Esterase. Biochem. J. 1969, 114, 711–717. [Google Scholar] [CrossRef] [Green Version]
- Rainier, S.; Bui, M.; Mark, E.; Thomas, D.; Tokarz, D.; Ming, L.; Delaney, C.; Richardson, R.J.; Albers, J.W.; Matsunami, N.; et al. Neuropathy Target Esterase Gene Mutations Cause Motor Neuron Disease. Am. J. Hum. Genet. 2008, 82, 780–785. [Google Scholar] [CrossRef] [Green Version]
- Deik, A.; Johannes, B.; Rucker, J.C.; Sánchez, E.; Brodie, S.E.; Deegan, E.; Landy, K.; Kajiwara, Y.; Scelsa, S.; Saunders-Pullman, R.; et al. Compound Heterozygous PNPLA6 Mutations Cause Boucher–Neuhäuser Syndrome with Late-Onset Ataxia. J. Neurol 2014, 261, 2411–2423. [Google Scholar] [CrossRef] [Green Version]
- Synofzik, M.; Gonzalez, M.A.; Lourenco, C.M.; Coutelier, M.; Haack, T.B.; Rebelo, A.; Hannequin, D.; Strom, T.M.; Prokisch, H.; Kernstock, C.; et al. PNPLA6 Mutations Cause Boucher-Neuhäuser and Gordon Holmes Syndromes as Part of a Broad Neurodegenerative Spectrum. Brain 2014, 137, 69–77. [Google Scholar] [CrossRef]
- Zaccheo, O.; Dinsdale, D.; Meacock, P.A.; Glynn, P. Neuropathy Target Esterase and Its Yeast Homologue Degrade Phosphatidylcholine to Glycerophosphocholine in Living Cells*. J. Biol. Chem. 2004, 279, 24024–24033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Da Cruz, A.B.; Wentzell, J.; Kretzschmar, D. Swiss Cheese, a Protein Involved in Progressive Neurodegeneration, Acts as a Noncanonical Regulatory Subunit for PKA-C3. J. Neurosci. 2008, 28, 10885–10892. [Google Scholar] [CrossRef] [PubMed]
- Wentzell, J.S.; Cassar, M.; Kretzschmar, D. Organophosphate-Induced Changes in the PKA Regulatory Function of Swiss Cheese/NTE Lead to Behavioral Deficits and Neurodegeneration. Plos One 2014, 9, e87526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kretzschmar, D.; Hasan, G.; Sharma, S.; Heisenberg, M.; Benzer, S. The Swiss Cheese Mutant Causes Glial Hyperwrapping and Brain Degeneration in Drosophila. J. Neurosci. 1997, 17, 7425–7432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mühlig-Versen, M.; Cruz, A.B. da; Tschäpe, J.-A.; Moser, M.; Büttner, R.; Athenstaedt, K.; Glynn, P.; Kretzschmar, D. Loss of Swiss Cheese/Neuropathy Target Esterase Activity Causes Disruption of Phosphatidylcholine Homeostasis and Neuronal and Glial Death in Adult Drosophila. J. Neurosci. 2005, 25, 2865–2873. [Google Scholar] [CrossRef] [Green Version]
- Dutta, S.; Rieche, F.; Eckl, N.; Duch, C.; Kretzschmar, D. Glial Expression of Swiss Cheese (SWS), the Drosophila Orthologue of Neuropathy Target Esterase (NTE), Is Required for Neuronal Ensheathment and Function. Dis. Models Mech. 2016, 9, 283–294. [Google Scholar] [CrossRef] [Green Version]
- Carlson, S.D.; Juang, J.-L.; Hilgers, S.L.; Garment, M.B. Blood Barriers of the Insect. Annu. Rev. Entomol. 2000, 45, 151–174. [Google Scholar] [CrossRef]
- Daneman, R.; Barres, B.A. The Blood-Brain Barrier— Lessons from Moody Flies. Cell 2005, 123, 9–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbott, N.J.; Rönnbäck, L.; Hansson, E. Astrocyte–Endothelial Interactions at the Blood–Brain Barrier. Nat. Rev. Neurosci. 2006, 7, 41–53. [Google Scholar] [CrossRef]
- Stork, T.; Engelen, D.; Krudewig, A.; Silies, M.; Bainton, R.J.; Klämbt, C. Organization and Function of the Blood–Brain Barrier in Drosophila. J. Neurosci. 2008, 28, 587–597. [Google Scholar] [CrossRef] [Green Version]
- Desalvo, M.K.; Mayer, N.; Mayer, F.; Bainton, R.J. Physiologic and Anatomic Characterization of the Brain Surface Glia Barrier of Drosophila. Glia 2011, 59, 1322–1340. [Google Scholar] [CrossRef] [Green Version]
- DeSalvo, M.K.; Hindle, S.J.; Rusan, Z.M.; Orng, S.; Eddison, M.; Halliwill, K.; Bainton, R.J. The Drosophila Surface Glia Transcriptome: Evolutionary Conserved Blood-Brain Barrier Processes. Front. Neurosci. 2014, 8. [Google Scholar] [CrossRef] [Green Version]
- Kremer, M.C.; Jung, C.; Batelli, S.; Rubin, G.M.; Gaul, U. The Glia of the Adult Drosophila Nervous System: Glia Anatomy in Adult Drosophila Nervous System. Glia 2017, 65, 606–638. [Google Scholar] [CrossRef] [Green Version]
- Awasaki, T.; Lai, S.-L.; Ito, K.; Lee, T. Organization and Postembryonic Development of Glial Cells in the Adult Central Brain of Drosophila. J. Neurosci. 2008, 28, 13742–13753. [Google Scholar] [CrossRef] [PubMed]
- Unhavaithaya, Y.; Orr-Weaver, T.L. Polyploidization of Glia in Neural Development Links Tissue Growth to Blood-Brain Barrier Integrity. Genes Dev. 2012, 26, 31–36. [Google Scholar] [CrossRef] [Green Version]
- Zülbahar, S.; Sieglitz, F.; Kottmeier, R.; Altenhein, B.; Rumpf, S.; Klämbt, C. Differential Expression of Öbek Controls Ploidy in the Drosophila Blood-Brain Barrier. Development 2018, 145, dev164111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juang, J.-L.; Carlson, S.D. A Blood-Brain Barrier without Tight Junctions in the Fly Central Nervous System in the Early Postembryonic Stage. Cell Tissue Res. 1992, 270, 95–103. [Google Scholar] [CrossRef]
- Baumgartner, S.; Littleton, J.T.; Broadie, K.; Bhat, M.A.; Harbecke, R.; Lengyel, J.A.; Chiquet-Ehrismann, R.; Prokop, A.; Bellen, H.J. A Drosophila Neurexin Is Required for Septate Junction and Blood-Nerve Barrier Formation and Function. Cell 1996, 87, 1059–1068. [Google Scholar] [CrossRef]
- Bhat, M.A.; Rios, J.C.; Lu, Y.; Garcia-Fresco, G.P.; Ching, W.; Martin, M.St.; Li, J.; Einheber, S.; Chesler, M.; Rosenbluth, J.; et al. Axon-Glia Interactions and the Domain Organization of Myelinated Axons Requires Neurexin IV/Caspr/Paranodin. Neuron 2001, 30, 369–383. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, S.; Pillai, A.M.; Paik, R.; Li, J.; Bhat, M.A. Axonal Ensheathment and Septate Junction Formation in the Peripheral Nervous System of Drosophila. J. Neurosci. 2006, 26, 3319–3329. [Google Scholar] [CrossRef] [Green Version]
- Kis, V.; Barti, B.; Lippai, M.; Sass, M. Specialized Cortex Glial Cells Accumulate Lipid Droplets in Drosophila Melanogaster. PLoS ONE 2015, 10, e0131250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Von Hilchen, C.M.; Beckervordersandforth, R.M.; Rickert, C.; Technau, G.M.; Altenhein, B. Identity, Origin, and Migration of Peripheral Glial Cells in the Drosophila Embryo. Mech. Dev. 2008, 125, 337–352. [Google Scholar] [CrossRef] [PubMed]
- Kottmeier, R.; Bittern, J.; Schoofs, A.; Scheiwe, F.; Matzat, T.; Pankratz, M.; Klämbt, C. Wrapping Glia Regulates Neuronal Signaling Speed and Precision in the Peripheral Nervous System of Drosophila. Nat. Commun. 2020, 11, 4491. [Google Scholar] [CrossRef]
- Matzat, T.; Sieglitz, F.; Kottmeier, R.; Babatz, F.; Engelen, D.; Klämbt, C. Axonal Wrapping in the Drosophila PNS Is Controlled by Glia-Derived Neuregulin Homolog Vein. Development 2015, 142, 1336–1345. [Google Scholar] [CrossRef] [Green Version]
- Kanda, H.; Shimamura, R.; Koizumi-Kitajima, M.; Okano, H. Degradation of Extracellular Matrix by Matrix Metalloproteinase 2 Is Essential for the Establishment of the Blood-Brain Barrier in Drosophila. iScience 2019, 16, 218–229. [Google Scholar] [CrossRef] [Green Version]
- Bainton, R.J.; Tsai, L.T.-Y.; Schwabe, T.; DeSalvo, M.; Gaul, U.; Heberlein, U. Moody Encodes Two GPCRs That Regulate Cocaine Behaviors and Blood-Brain Barrier Permeability in Drosophila. Cell 2005, 123, 145–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kretzschmar, D. Swiss Cheese et Allii, Some of the First Neurodegenerative Mutants Isolated in Drosophila. J. Neurogenet. 2009, 23, 34–41. [Google Scholar] [CrossRef] [Green Version]
- Gargano, J.W.; Martin, I.; Bhandari, P.; Grotewiel, M.S. Rapid Iterative Negative Geotaxis (RING): A New Method for Assessing Age-Related Locomotor Decline in Drosophila. Exp. Gerontol. 2005, 40, 386–395. [Google Scholar] [CrossRef]
- Quinn, W.G.; Harris, W.A.; Benzer, S. Conditioned Behavior in Drosophila Melanogaster. PNAS 1974, 71, 708–712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tully, T.; Quinn, W.G. Classical Conditioning and Retention in Normal and MutantDrosophila Melanogaster. J. Comp. Physiol. 1985, 157, 263–277. [Google Scholar] [CrossRef]
- Pérez-Severiano, F.; Rodríguez-Pérez, M.; Pedraza-Chaverrí, J.; Maldonado, P.D.; Medina-Campos, O.N.; Ortíz-Plata, A.; Sánchez-García, A.; Villeda-Hernández, J.; Galván-Arzate, S.; Aguilera, P.; et al. S-Allylcysteine, a Garlic-Derived Antioxidant, Ameliorates Quinolinic Acid-Induced Neurotoxicity and Oxidative Damage in Rats. Neurochem. Int. 2004, 45, 1175–1183. [Google Scholar] [CrossRef]
- Albrecht, S.C.; Barata, A.G.; Großhans, J.; Teleman, A.A.; Dick, T.P. In Vivo Mapping of Hydrogen Peroxide and Oxidized Glutathione Reveals Chemical and Regional Specificity of Redox Homeostasis. Cell Metab. 2011, 14, 819–829. [Google Scholar] [CrossRef] [Green Version]
- Barata, A.G.; Dick, T.P. Chapter Four - In Vivo Imaging of H2O2 Production in Drosophila. Methods in Enzymol. 2013, 526, 61–82. [Google Scholar]
- Reimand, J.; Isserlin, R.; Voisin, V.; Kucera, M.; Tannus-Lopes, C.; Rostamianfar, A.; Wadi, L.; Meyer, M.; Wong, J.; Xu, C.; et al. Pathway Enrichment Analysis and Visualization of Omics Data Using g:Profiler, GSEA, Cytoscape and EnrichmentMap. Nat. Protoc. 2019, 14, 482–517. [Google Scholar] [CrossRef] [PubMed]
- Ryabova, E.; Matiytsiv, N.; Trush, O.; Mohylyak, I.; Kislik, G.; Melentev, P.; Sarantseva, S. Swiss Cheese, Drosophila Ortholog of Hereditary Spastic Paraplegia Gene NTE, Maintains Neuromuscular Junction Development and Microtubule Network. In Drosophila melanogaster—Model for Recent Advances in Genetics and Therapeutics; Perveen, F.K., Ed.; IntechOpen: London, UK, 2018. [Google Scholar] [CrossRef] [Green Version]
- Schwabe, T.; Bainton, R.J.; Fetter, R.D.; Heberlein, U.; Gaul, U. GPCR Signaling Is Required for Blood-Brain Barrier Formation in Drosophila. Cell 2005, 123, 133–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coutinho-Budd, J.C.; Sheehan, A.E.; Freeman, M.R. The Secreted Neurotrophin Spätzle 3 Promotes Glial Morphogenesis and Supports Neuronal Survival and Function. Genes Dev. 2017, 31, 2023–2038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khwaja, F.S.; Quann, E.J.; Pattabiraman, N.; Wynne, S.; Djakiew, D. Carprofen Induction of P75NTR-Dependent Apoptosis via the P38 Mitogen-Activated Protein Kinase Pathway in Prostate Cancer Cells. Mol Cancer 2008, 7, 3539–3545. [Google Scholar] [CrossRef] [Green Version]
- Nhiem, N.X.; Thu, V.K.; Van Kiem, P.; Van Minh, C.; Tai, B.H.; Quang, T.H.; Cuong, N.X.; Yen, P.H.; Boo, H.-J.; Kang, J.-I.; et al. Cytotoxic Oleane-Type Triterpene Saponins from Glochidion Eriocarpum. Arch. Pharm. Res. 2012, 35, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Singhania, A.; Grueber, W.B. Development of the Embryonic and Larval Peripheral Nervous System of Drosophila. Wires Dev. Biol. 2014, 3, 193–210. [Google Scholar] [CrossRef] [Green Version]
- Subramanian, A.; Siefert, M.; Banerjee, S.; Vishal, K.; Bergmann, K.A.; Curts, C.C.M.; Dorr, M.; Molina, C.; Fernandes, J. Remodeling of Peripheral Nerve Ensheathment during the Larval-to-Adult Transition in Drosophila. Dev. Neurobiol. 2017, 77, 1144–1160. [Google Scholar] [CrossRef]
- Kounatidis, I.; Chtarbanova, S. Role of Glial Immunity in Lifespan Determination: A Drosophila Perspective. Front. Immunol. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Angelova, P.R.; Abramov, A.Y. Role of Mitochondrial ROS in the Brain: From Physiology to Neurodegeneration. FEBS Lett. 2018, 592, 692–702. [Google Scholar] [CrossRef]
- Barres, B.A. Five Electrophysiological Properties of Glial Cells. Ann. N. Y. Acad. Sci. 1991, 633, 248–254. [Google Scholar] [CrossRef]
- Pfrieger, F.W.; Barres, B.A. What the Fly’s Glia Tell the Fly’s Brain. Cell 1995, 83, 671–674. [Google Scholar] [CrossRef] [Green Version]
- Barres, B.A.; Raff, M.C. Axonal Control of Oligodendrocyte Development. J. Cell Biol. 1999, 147, 1123–1128. [Google Scholar] [CrossRef]
- Eroglu, C.; Barres, B.A. Regulation of Synaptic Connectivity by Glia. Nature 2010, 468, 223–231. [Google Scholar] [CrossRef]
- Allen, N.J.; Lyons, D.A. Glia as Architects of Central Nervous System Formation and Function. Science 2018, 362, 181–185. [Google Scholar] [CrossRef] [Green Version]
- Freeman, M.R. Drosophila Central Nervous System Glia. Cold Spring Harb. Perspect. Biol. 2015, 7, a020552. [Google Scholar] [CrossRef] [Green Version]
- Sasse, S.; Neuert, H.; Klämbt, C. Differentiation of Drosophila Glial Cells. Wires Dev. Biol. 2015, 4, 623–636. [Google Scholar] [CrossRef] [PubMed]
- Yildirim, K.; Petri, J.; Kottmeier, R.; Klämbt, C. Drosophila Glia: Few Cell Types and Many Conserved Functions. Glia 2019, 67, 5–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hindle, S.J.; Bainton, R.J. Barrier Mechanisms in the Drosophila Blood-Brain Barrier. Front. Neurosci. 2014, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Limmer, S.; Weiler, A.; Volkenhoff, A.; Babatz, F.; Klämbt, C. The Drosophila Blood-Brain Barrier: Development and Function of a Glial Endothelium. Front. Neurosci. 2014, 8. [Google Scholar] [CrossRef] [Green Version]
- Von Hilchen, C.M.; Bustos, A.E.; Giangrande, A.; Technau, G.M.; Altenhein, B. Predetermined Embryonic Glial Cells Form the Distinct Glial Sheaths of the Drosophila Peripheral Nervous System. Development 2013, 140, 3657–3668. [Google Scholar] [CrossRef] [Green Version]
- Navarro, J.A.; Ohmann, E.; Sanchez, D.; Botella, J.A.; Liebisch, G.; Moltó, M.D.; Ganfornina, M.D.; Schmitz, G.; Schneuwly, S. Altered Lipid Metabolism in a Drosophila Model of Friedreich’s Ataxia. Hum. Mol. Genet. 2010, 19, 2828–2840. [Google Scholar] [CrossRef] [Green Version]
- Bailey, A.P.; Koster, G.; Guillermier, C.; Hirst, E.M.A.; MacRae, J.I.; Lechene, C.P.; Postle, A.D.; Gould, A.P. Antioxidant Role for Lipid Droplets in a Stem Cell Niche of Drosophila. Cell 2015, 163, 340–353. [Google Scholar] [CrossRef] [Green Version]
- Cabirol-Pol, M.-J.; Khalil, B.; Rival, T.; Faivre-Sarrailh, C.; Besson, M.T. Glial Lipid Droplets and Neurodegeneration in a Drosophila Model of Complex I Deficiency. Glia 2018, 66, 874–888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sunderhaus, E.R.; Law, A.D.; Kretzschmar, D. ER Responses Play a Key Role in Swiss-Cheese/Neuropathy Target Esterase-Associated Neurodegeneration. Neurobiol. Dis. 2019, 130, 104520. [Google Scholar] [CrossRef] [PubMed]
- Verheggen, I.C.M.; de Jong, J.J.A.; van Boxtel, M.P.J.; Gronenschild, E.H.B.M.; Palm, W.M.; Postma, A.A.; Jansen, J.F.A.; Verhey, F.R.J.; Backes, W.H. Increase in Blood–Brain Barrier Leakage in Healthy, Older Adults. GeroScience 2020, 42, 1183–1193. [Google Scholar] [CrossRef] [PubMed]
- Nation, D.A.; Sweeney, M.D.; Montagne, A.; Sagare, A.P.; D’Orazio, L.M.; Pachicano, M.; Sepehrband, F.; Nelson, A.R.; Buennagel, D.P.; Harrington, M.G.; et al. Blood–Brain Barrier Breakdown Is an Early Biomarker of Human Cognitive Dysfunction. Nat. Med. 2019, 25, 270–276. [Google Scholar] [CrossRef]
- Zlokovic, B.V. The Blood-Brain Barrier in Health and Chronic Neurodegenerative Disorders. Neuron 2008, 57, 178–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melentev, P.A.; Agranovich, O.E.; Sarantseva, S.V. Human diseases associated with NTE gene. Ecol. Genet. 2020, 18, 229–242. [Google Scholar] [CrossRef]
- McFerrin, J.; Patton, B.L.; Sunderhaus, E.R.; Kretzschmar, D. NTE/PNPLA6 Is Expressed in Mature Schwann Cells and Is Required for Glial Ensheathment of Remak Fibers. Glia 2017, 65, 804–816. [Google Scholar] [CrossRef] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ryabova, E.V.; Melentev, P.A.; Komissarov, A.E.; Surina, N.V.; Ivanova, E.A.; Matiytsiv, N.; Shcherbata, H.R.; Sarantseva, S.V. Morpho-Functional Consequences of Swiss Cheese Knockdown in Glia of Drosophila melanogaster. Cells 2021, 10, 529. https://doi.org/10.3390/cells10030529
Ryabova EV, Melentev PA, Komissarov AE, Surina NV, Ivanova EA, Matiytsiv N, Shcherbata HR, Sarantseva SV. Morpho-Functional Consequences of Swiss Cheese Knockdown in Glia of Drosophila melanogaster. Cells. 2021; 10(3):529. https://doi.org/10.3390/cells10030529
Chicago/Turabian StyleRyabova, Elena V., Pavel A. Melentev, Artem E. Komissarov, Nina V. Surina, Ekaterina A. Ivanova, Natalia Matiytsiv, Halyna R. Shcherbata, and Svetlana V. Sarantseva. 2021. "Morpho-Functional Consequences of Swiss Cheese Knockdown in Glia of Drosophila melanogaster" Cells 10, no. 3: 529. https://doi.org/10.3390/cells10030529
APA StyleRyabova, E. V., Melentev, P. A., Komissarov, A. E., Surina, N. V., Ivanova, E. A., Matiytsiv, N., Shcherbata, H. R., & Sarantseva, S. V. (2021). Morpho-Functional Consequences of Swiss Cheese Knockdown in Glia of Drosophila melanogaster. Cells, 10(3), 529. https://doi.org/10.3390/cells10030529