
Collagen Assembly at the Cell Surface: Dogmas Revisited

 , , , and
, , , and 
Abstract
:1. Introduction
2. Components of the Interstitial ECM
2.1. The Collagen Family—Fibrillar Collagens
2.2. Fibronectin
2.3. Tenascins and Other ECM Proteins in the Interstitial Matrix Influencing Collagen Organization
2.3.1. Tenascins
2.3.2. Periostin
2.3.3. Stromal Proteoglycans
3. Mechanisms of ECM Assembly
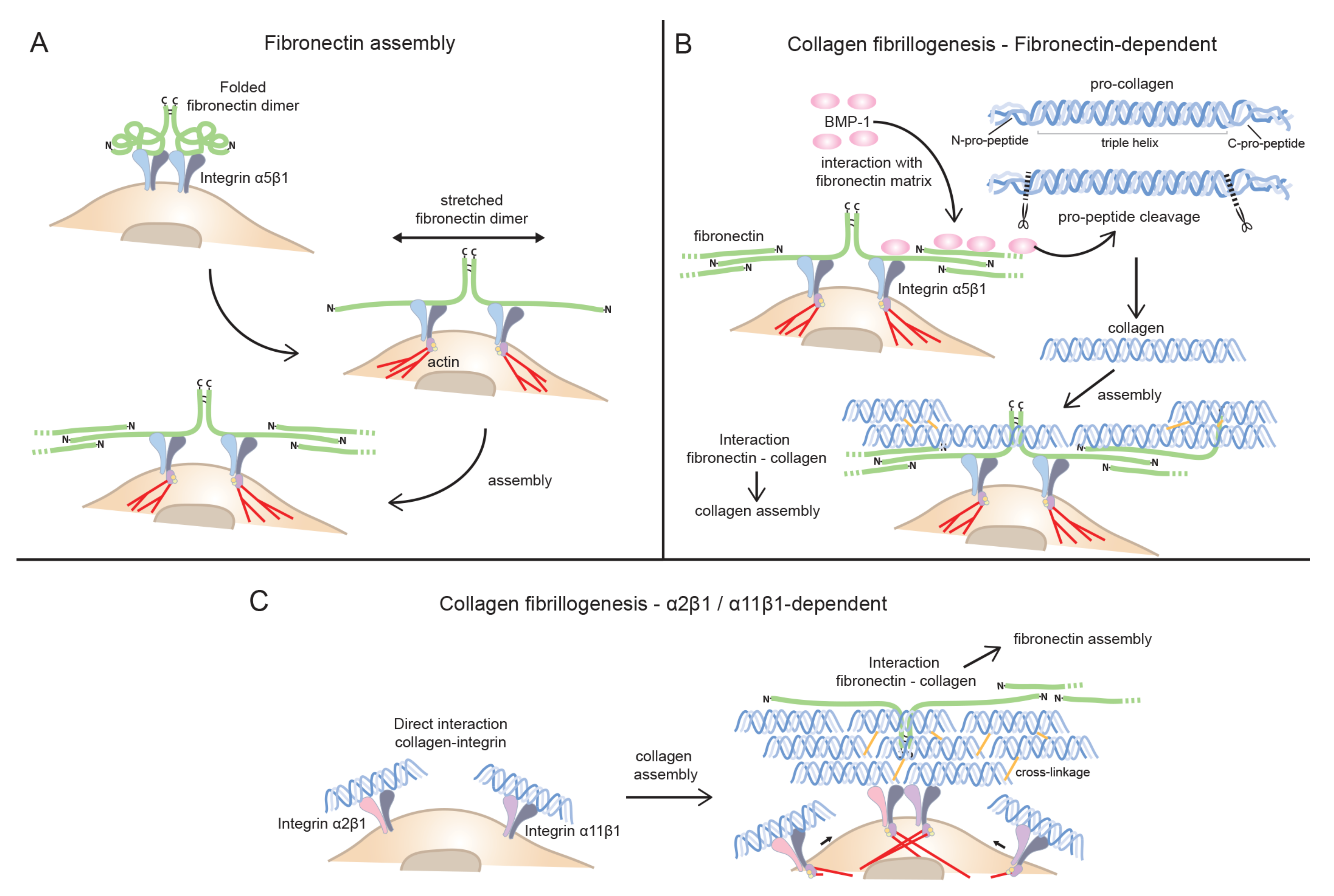
3.1. Fibronectin Fibrillogenesis—Integrin Initiated Assembly, a Central Mechanism for Matrix Assembly in Cell Culture

{kind=link}
{kind=link}
| Protein | Type | Role in Fibronectin (FN) Fibrillogenesis In Vitro | Reference |
|---|---|---|---|
| α5β1 integrin | Membrane receptor | Primary FN assembly receptor. | [102] |
| α4β1 integrin | Membrane receptor | May contribute to FN assembly. | [33] |
| αvβ3 integrin | Membrane receptor | May contribute to FN assembly. | [8,34] |
| Syndecan-1 (SDC1), syndecan-4 (SDC4) | Membrane receptors | SDC1 implicated in FN fibrillogenesis. SDC4 Co-receptor for integrins, but also signaling cross-talk. Heparin can inhibit FN fibrillogenesis. | [77,82,84] |
| AMP-activated protein kinase (AMPK) | Kinase | Repress fibrillogenesis by negatively regulating tensin-dependent integrin activity. | [97] |
| Integrin linked kinase (ILK) | Adapter protein, pseudokinase | Promote fibrillogenesis by stimulating focal adhesion maturation and fibrillar adhesion formation. | [98] |
| RhoA | Small GTPase | Allow formation of α5β1 containing fibrillar adhesions. | [103] |
| RhoJ | Small GTPase | Repress fibrillogenesis by diverting α5β1 integrin into degradative fate. | [100] |
| Cdc42 | Small GTPase | Stimulates FN fibrillogenesis in endothelial cells in vitro. | [101] |
| Tensins | Cytoskeletal protein | Bind to phosphorylated proximal NPXY motif in integrin β1 chain with higher affinity than talin. | [92] |
| Hic-5 | Scaffolding protein | Promote fibrillogenesis by stabilizing tensin-β1 integrin interaction. | [99] |
3.2. Laminin Assembly—Uncontroversial Roles of Sulfatides, Integrins and Dystroglycan
| Ligand | Integrin | Cells | References |
|---|---|---|---|
| Fibronectin | |||
| (matrix assembly in vitro) | α5β1 | Fibroblasts, stromal cells | [102] |
| αvβ3 | Mouse embryonic cells | [8,34] | |
| α4β1 | RAMOS B-Cells, α4 integrin-transfected CHO cells | [33] | |
| Laminins | |||
| (binding/matrix assembly in vitro) | β1 | Mouse embryonic stem cells | [104] |
| α3β1 | Endothelial cells | [105] | |
| α6β1 | Epithelial cells, endothelial cells | [106] | |
| α7β1 | Muscle cells | [107,108] | |
| Collagen Ⅰ/Ⅲ | |||
| (matrix assembly in vitro) | α2β1 | Vascular smooth muscle cells, mouse embryonic cells transfected with collagen-binding integrins | [109,110] |
| α11β1 | Mouse embryonic cells transfected with collagen-binding integrins | [110] | |
| Collagen Ⅴ | |||
| (binding) | α2β1 | Human epithelial cells | [111] |
| α11β1 | Mouse embryonic fibroblasts | [112] |
3.3. Cell Surface—Mediated Collagen Fibril Assembly—Direct or Indirect Integrin Links
3.3.1. Fibronectin as the Main Player in Collagen Fibrillogenesis: Indirect Mode of Assembly
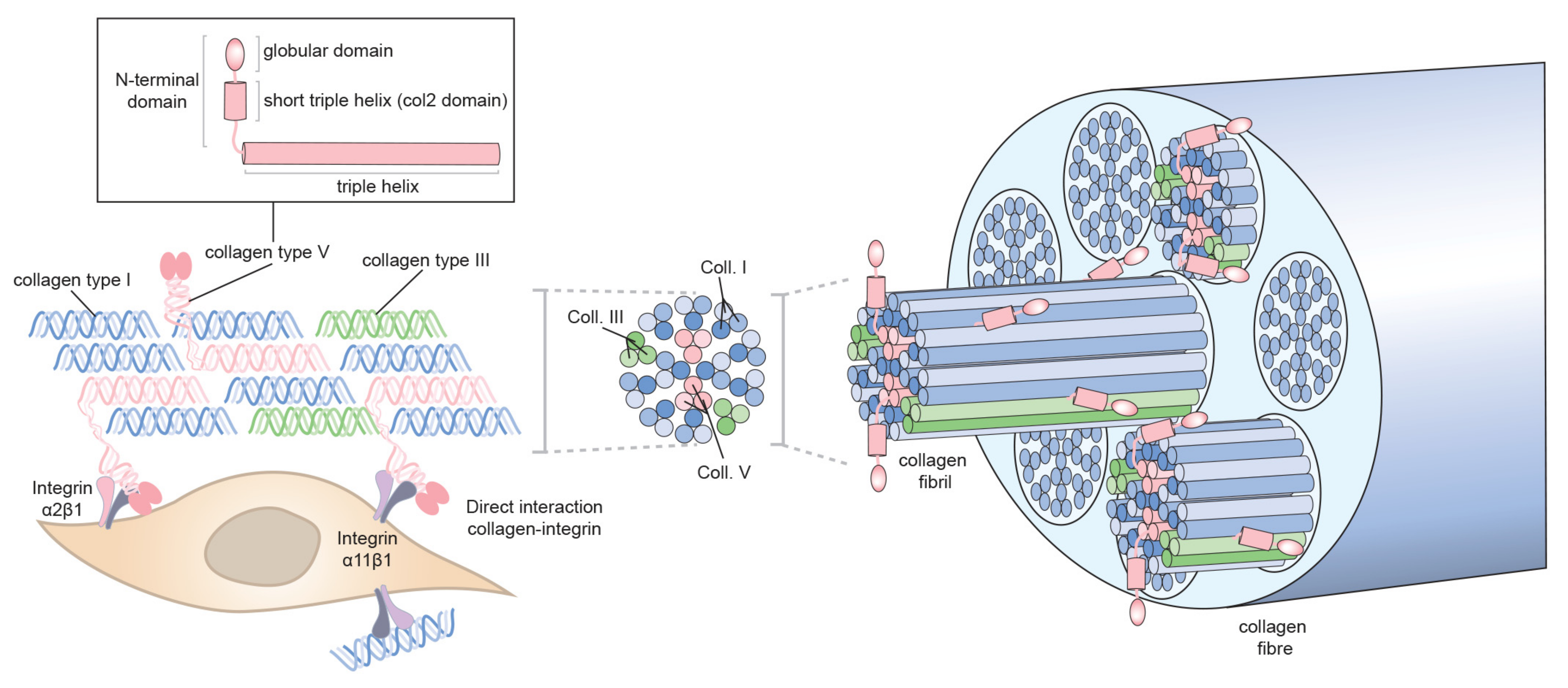
3.3.2. Collagen-binding Integrins in Collagen Fibrillogenesis: Direct Mode of Assembly
3.3.3. Role of Collagen Ⅴ/Ⅺ in Collagen Ⅰ Fibrillogenesis?
4. Who Is Leading the Way—Fibronectin or Collagen—Or a Joint Venture?
- Collagens are assembled and secreted as procollagen molecules, and pro-peptides can be cleaved off extracellularly in close association with cells surface. FN can aid in BMP-1-mediated extracellular collagen Ⅰ pro-peptide cleavage and in this way help to facilitate collagen I fibrillogenesis at the cell surface.
- Binding to cell surface integrins enable cell-directed collagen fibrillogenesis either directly, by collagen-binding integrins or indirectly, by being captured by FN fibrils which have first been assembled by FN receptors.
- The relative contribution of FN and collagen-binding integrins to collagen fibrillogenesis most likely vary in a dynamic manner during physiological and pathological processes.
- Collagens Ⅴ/Ⅺ have a nucleating role in formation of heterotypic fibrils and might be the first collagens to bind to cell surface via collagen-binding integrins.
Funding
Acknowledgments
Conflicts of Interest
References
- Hynes, R.O. The emergence of integrins: A personal and historical perspective. Matrix Biol. 2004, 23, 333–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamkun, J.W.; DeSimone, D.W.; Fonda, D.; Patel, R.S.; Buck, C.; Horwitz, A.F.; Hynes, R.O. Structure of integrin, a glycoprotein involved in the transmembrane linkage between fibronectin and actin. Cell 1986, 46, 271–282. [Google Scholar] [CrossRef]
- Hynes, R.O. Integrins: Versatility, modulation, and signaling in cell adhesion. Cell 1992, 69, 11–25. [Google Scholar] [CrossRef]
- Bissell, M.J.; Aggeler, J. Dynamic reciprocity: How do extracellular matrix and hormones direct gene expression? Prog. Clin. Biol. Res. 1987, 249, 251–262. [Google Scholar]
- Roskelley, C.D.; Bissell, M.J. Dynamic reciprocity revisited: A continuous, bidirectional flow of information between cells and the extracellular matrix regulates mammary epithelial cell function. Biochem. Cell Biol. 1995, 73, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.; Boudreau, A.; Bissell, M.J. Tissue architecture and function: Dynamic reciprocity via extra- and intra-cellular matrices. Cancer Metastasis Rev. 2009, 28, 167–176. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.T.; Rayburn, H.; Hynes, R.O. Embryonic mesodermal defects in alpha 5 integrin-deficient mice. Development 1993, 119, 1093–1105. [Google Scholar]
- Yang, J.T.; Hynes, R.O. Fibronectin receptor functions in embryonic cells deficient in alpha 5 beta 1 integrin can be replaced by alpha V integrins. Mol. Biol. Cell 1996, 7, 1737–1748. [Google Scholar] [CrossRef] [Green Version]
- Wewer, U.M.; Engval, E. Merosin/laminin-2 and muscular dystrophy. Neuromuscul. Disord. 1996, 6, 409–418. [Google Scholar] [CrossRef]
- Aumailley, M.; Smyth, N. The role of laminins in basement membrane function. J. Anat. 1998, 193, 1–21. [Google Scholar] [CrossRef]
- Gullberg, D.; Tiger, C.F.; Velling, T. Laminins during muscle development and in muscular dystrophies. Cell Mol. Life Sci. 1999, 56, 442–460. [Google Scholar] [CrossRef]
- Santoro, S.A. Identification of a 160,000 dalton platelet membrane protein that mediates the initial divalent cation-dependent adhesion of platelets to collagen. Cell 1986, 46, 913–920. [Google Scholar] [CrossRef]
- Barnes, M.J.; Knight, C.G.; Farndale, R.W. The collagen-platelet interaction. Curr. Opin. Hematol 1998, 5, 314–320. [Google Scholar] [CrossRef]
- Piersma, B.; Hayward, M.K.; Weaver, V.M. Fibrosis and cancer: A strained relationship. Biochim. Biophys. Acta. Rev. Cancer 2020, 1873, 188356. [Google Scholar] [CrossRef]
- Kubow, K.E.; Vukmirovic, R.; Zhe, L.; Klotzsch, E.; Smith, M.L.; Gourdon, D.; Luna, S.; Vogel, V. Mechanical forces regulate the interactions of fibronectin and collagen I in extracellular matrix. Nat. Commun. 2015, 6, 8026. [Google Scholar] [CrossRef] [Green Version]
- Graham, J.; Raghunath, M.; Vogel, V. Fibrillar fibronectin plays a key role as nucleator of collagen I polymerization during macromolecular crowding-enhanced matrix assembly. Biomater Sci. 2019, 7, 4519–4535. [Google Scholar] [CrossRef] [Green Version]
- Zeltz, C.; Orgel, J.; Gullberg, D. Molecular composition and function of integrin-based collagen glues-introducing COLINBRIs. Biochim. Biophys. Acta. 2014, 1840, 2533–2548. [Google Scholar] [CrossRef] [Green Version]
- Bourgot, I.; Primac, I.; Louis, T.; Noel, A.; Maquoi, E. Reciprocal Interplay Between Fibrillar Collagens and Collagen-Binding Integrins: Implications in Cancer Progression and Metastasis. Front. Oncol. 2020, 10, 1488. [Google Scholar] [CrossRef]
- Zeltz, C.; Gullberg, D. The integrin-collagen connection—A glue for tissue repair? J. Cell Sci. 2016, 129, 653–664. [Google Scholar] [CrossRef] [Green Version]
- Zeltz, C.; Primac, I.; Erusappan, P.; Alam, J.; Noel, A.; Gullberg, D. Cancer-associated fibroblasts in desmoplastic tumors: Emerging role of integrins. Semin. Cancer Biol. 2020, 62, 166–181. [Google Scholar] [CrossRef]
- Emsley, J.; Knight, C.G.; Farndale, R.W.; Barnes, M.J.; Liddington, R.C. Structural basis of collagen recognition by integrin alpha2beta1. Cell 2000, 101, 47–56. [Google Scholar] [CrossRef]
- Woltersdorf, C.; Bonk, M.; Leitinger, B.; Huhtala, M.; Kapyla, J.; Heino, J.; Gil Girol, C.; Niland, S.; Eble, J.A.; Bruckner, P.; et al. The binding capacity of alpha1beta1-, alpha2beta1- and alpha10beta1-integrins depends on non-collagenous surface macromolecules rather than the collagens in cartilage fibrils. Matrix Biol. 2017, 63, 91–105. [Google Scholar] [CrossRef] [Green Version]
- Ricard-Blum, S. The collagen family. Cold Spring Harb. Perspect. Biol. 2011, 3, a004978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.; Ge, G. Complexity of type IV collagens: From network assembly to function. Biol. Chem. 2019, 400, 565–574. [Google Scholar] [CrossRef] [PubMed]
- Yeung, J.; Adili, R.; Stringham, E.N.; Luo, R.; Vizurraga, A.; Rosselli-Murai, L.K.; Stoveken, H.M.; Yu, M.; Piao, X.; Holinstat, M.; et al. GPR56/ADGRG1 is a platelet collagen-responsive GPCR and hemostatic sensor of shear force. Proc. Natl. Acad. Sci. USA 2020, 117, 28275–28286. [Google Scholar] [CrossRef]
- Gondelaud, F.; Ricard-Blum, S. Structures and interactions of syndecans. Febs. J. 2019, 286, 2994–3007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vuoriluoto, K.; Hognas, G.; Meller, P.; Lehti, K.; Ivaska, J. Syndecan-1 and -4 differentially regulate oncogenic K-ras dependent cell invasion into collagen through alpha2beta1 integrin and MT1-MMP. Matrix Biol. 2011, 30, 207–217. [Google Scholar] [CrossRef]
- Kadler, K.E.; Hill, A.; Canty-Laird, E.G. Collagen fibrillogenesis: Fibronectin, integrins, and minor collagens as organizers and nucleators. Curr. Opin. Cell Biol. 2008, 20, 495–501. [Google Scholar] [CrossRef]
- Hynes, R. Molecular biology of fibronectin. Annu Rev. Cell Biol. 1985, 1, 67–90. [Google Scholar] [CrossRef]
- Pankov, R.; Yamada, K.M. Fibronectin at a glance. J. Cell Sci. 2002, 115, 3861–3863. [Google Scholar] [CrossRef] [Green Version]
- White, E.S.; Baralle, F.E.; Muro, A.F. New insights into form and function of fibronectin splice variants. J. Pathol. 2008, 216, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sechler, J.L.; Corbett, S.A.; Schwarzbauer, J.E. Modulatory roles for integrin activation and the synergy site of fibronectin during matrix assembly. Mol. Biol. Cell 1997, 8, 2563–2573. [Google Scholar] [CrossRef] [Green Version]
- Sechler, J.L.; Cumiskey, A.M.; Gazzola, D.M.; Schwarzbauer, J.E. A novel RGD-independent fibronectin assembly pathway initiated by alpha4beta1 integrin binding to the alternatively spliced V region. J. Cell Sci. 2000, 113 Pt 8, 1491–1498. [Google Scholar]
- Wennerberg, K.; Lohikangas, L.; Gullberg, D.; Pfaff, M.; Johansson, S.; Fassler, R. Beta 1 integrin-dependent and -independent polymerization of fibronectin. J. Cell Biol. 1996, 132, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Doyle, A.D.; Shinsato, Y.; Wang, S.; Bodendorfer, M.A.; Zheng, M.; Yamada, K.M. Basement Membrane Regulates Fibronectin Organization Using Sliding Focal Adhesions Driven by a Contractile Winch. Dev. Cell 2020, 52, 631–646.e634. [Google Scholar] [CrossRef]
- Hahn, L.H.; Yamada, K.M. Identification and isolation of a collagen-binding fragment of the adhesive glycoprotein fibronectin. Proc. Natl. Acad. Sci. USA 1979, 76, 1160–1163. [Google Scholar] [CrossRef] [Green Version]
- Erat, M.C.; Slatter, D.A.; Lowe, E.D.; Millard, C.J.; Farndale, R.W.; Campbell, I.D.; Vakonakis, I. Identification and structural analysis of type I collagen sites in complex with fibronectin fragments. Proc. Natl. Acad. Sci. USA 2009, 106, 4195–4200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erat, M.C.; Schwarz-Linek, U.; Pickford, A.R.; Farndale, R.W.; Campbell, I.D.; Vakonakis, I. Implications for collagen binding from the crystallographic structure of fibronectin 6FnI1-2FnII7FnI. J. Biol. Chem. 2010, 285, 33764–33770. [Google Scholar] [CrossRef] [Green Version]
- Astrof, S.; Crowley, D.; George, E.L.; Fukuda, T.; Sekiguchi, K.; Hanahan, D.; Hynes, R.O. Direct test of potential roles of EIIIA and EIIIB alternatively spliced segments of fibronectin in physiological and tumor angiogenesis. Mol. Cell Biol. 2004, 24, 8662–8670. [Google Scholar] [CrossRef] [Green Version]
- Singh, P.; Reimer, C.L.; Peters, J.H.; Stepp, M.A.; Hynes, R.O.; Van De Water, L. The spatial and temporal expression patterns of integrin alpha9beta1 and one of its ligands, the EIIIA segment of fibronectin, in cutaneous wound healing. J. Investig. Derm. 2004, 123, 1176–1181. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharyya, S.; Tamaki, Z.; Wang, W.; Hinchcliff, M.; Hoover, P.; Getsios, S.; White, E.S.; Varga, J. FibronectinEDA promotes chronic cutaneous fibrosis through Toll-like receptor signaling. Sci. Transl. Med. 2014, 6, 232ra250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rybinski, B.; Franco-Barraza, J.; Cukierman, E. The wound healing, chronic fibrosis, and cancer progression triad. Physiol. Genom. 2014, 46, 223–244. [Google Scholar] [CrossRef]
- Rybak, J.N.; Roesli, C.; Kaspar, M.; Villa, A.; Neri, D. The extra-domain A of fibronectin is a vascular marker of solid tumors and metastases. Cancer Res. 2007, 67, 10948–10957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsumoto, E.; Yoshida, T.; Kawarada, Y.; Sakakura, T. Expression of fibronectin isoforms in human breast tissue: Production of extra domain A+/extra domain B+ by cancer cells and extra domain A+ by stromal cells. Jpn J. Cancer Res. 1999, 90, 320–325. [Google Scholar] [CrossRef]
- Pujuguet, P.; Hammann, A.; Moutet, M.; Samuel, J.L.; Martin, F.; Martin, M. Expression of fibronectin ED-A+ and ED-B+ isoforms by human and experimental colorectal cancer. Contribution of cancer cells and tumor-associated myofibroblasts. Am. J. Pathol. 1996, 148, 579–592. [Google Scholar] [PubMed]
- Manabe, R.; Ohe, N.; Maeda, T.; Fukuda, T.; Sekiguchi, K. Modulation of cell-adhesive activity of fibronectin by the alternatively spliced EDA segment. J. Cell Biol. 1997, 139, 295–307. [Google Scholar] [CrossRef] [Green Version]
- Shinde, A.V.; Bystroff, C.; Wang, C.; Vogelezang, M.G.; Vincent, P.A.; Hynes, R.O.; Van De Water, L. Identification of the peptide sequences within the EIIIA (EDA) segment of fibronectin that mediate integrin alpha9beta1-dependent cellular activities. J. Biol. Chem. 2008, 283, 2858–2870. [Google Scholar] [CrossRef] [Green Version]
- Kohan, M.; Muro, A.F.; White, E.S.; Berkman, N. EDA-containing cellular fibronectin induces fibroblast differentiation through binding to alpha4beta7 integrin receptor and MAPK/Erk 1/2-dependent signaling. Faseb. J. 2010, 24, 4503–4512. [Google Scholar] [CrossRef]
- Gopal, S.; Veracini, L.; Grall, D.; Butori, C.; Schaub, S.; Audebert, S.; Camoin, L.; Baudelet, E.; Radwanska, A.; Beghelli-de la Forest Divonne, S.; et al. Fibronectin-guided migration of carcinoma collectives. Nat. Commun. 2017, 8, 14105. [Google Scholar] [CrossRef] [PubMed]
- Kwon, A.; Chae, I.H.; You, E.; Kim, S.H.; Ahn, S.Y.; Lee, O.J.; Park, Z.Y.; Rhee, S.; Huh, Y.H.; Song, W.K. Extra domain A-containing fibronectin expression in Spin90-deficient fibroblasts mediates cancer-stroma interaction and promotes breast cancer progression. J. Cell Physiol. 2020, 235, 4494–4507. [Google Scholar] [CrossRef]
- Mosher, D.F.; Adams, J.C. Adhesion-modulating/matricellular ECM protein families: A structural, functional and evolutionary appraisal. Matrix Biol. 2012, 31, 155–161. [Google Scholar] [CrossRef]
- Gerarduzzi, C.; Hartmann, U.; Leask, A.; Drobetsky, E. The Matrix Revolution: Matricellular Proteins and Restructuring of the Cancer Microenvironment. Cancer Res. 2020, 80, 2705–2717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiquet-Ehrismann, R.; Tucker, R.P. Tenascins and the importance of adhesion modulation. Cold Spring Harb. Perspect. Biol. 2011, 3. [Google Scholar] [CrossRef] [Green Version]
- Zuliani-Alvarez, L.; Marzeda, A.M.; Deligne, C.; Schwenzer, A.; McCann, F.E.; Marsden, B.D.; Piccinini, A.M.; Midwood, K.S. Mapping tenascin-C interaction with toll-like receptor 4 reveals a new subset of endogenous inflammatory triggers. Nat. Commun. 2017, 8, 1595. [Google Scholar] [CrossRef] [PubMed]
- Spenle, C.; Loustau, T.; Murdamoothoo, D.; Erne, W.; Beghelli-de la Forest Divonne, S.; Veber, R.; Petti, L.; Bourdely, P.; Morgelin, M.; Brauchle, E.M.; et al. Tenascin-C Orchestrates an Immune-Suppressive Tumor Microenvironment in Oral Squamous Cell Carcinoma. Cancer Immunol. Res. 2020, 8, 1122–1138. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, S.; Wang, W.; Morales-Nebreda, L.; Feng, G.; Wu, M.; Zhou, X.; Lafyatis, R.; Lee, J.; Hinchcliff, M.; Feghali-Bostwick, C.; et al. Tenascin-C drives persistence of organ fibrosis. Nat. Commun. 2016, 7, 11703. [Google Scholar] [CrossRef] [PubMed]
- Katoh, D.; Kozuka, Y.; Noro, A.; Ogawa, T.; Imanaka-Yoshida, K.; Yoshida, T. Tenascin-C induces phenotypic changes in fibroblasts to myofibroblasts with high contractility through the integrin alphavbeta1/TGF-beta/SMAD signaling axis in human breast cancer. Am. J. Pathol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, K.; Saga, Y.; Ikemura, T.; Sakakura, T.; Chiquet-Ehrismann, R. The distribution of tenascin-X is distinct and often reciprocal to that of tenascin-C. J. Cell Biol. 1994, 125, 483–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiquet-Ehrismann, R.; Chiquet, M. Tenascins: Regulation and putative functions during pathological stress. J. Pathol. 2003, 200, 488–499. [Google Scholar] [CrossRef]
- Minamitani, T.; Ikuta, T.; Saito, Y.; Takebe, G.; Sato, M.; Sawa, H.; Nishimura, T.; Nakamura, F.; Takahashi, K.; Ariga, H.; et al. Modulation of collagen fibrillogenesis by tenascin-X and type VI collagen. Exp. Cell Res. 2004, 298, 305–315. [Google Scholar] [CrossRef]
- Egging, D.; van den Berkmortel, F.; Taylor, G.; Bristow, J.; Schalkwijk, J. Interactions of human tenascin-X domains with dermal extracellular matrix molecules. Arch. Derm. Res. 2007, 298, 389–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bristow, J.; Carey, W.; Egging, D.; Schalkwijk, J. Tenascin-X, collagen, elastin, and the Ehlers-Danlos syndrome. Am. J. Med. Genet. C Semin. Med. Genet. 2005, 139C, 24–30. [Google Scholar] [CrossRef] [Green Version]
- Elefteriou, F.; Exposito, J.Y.; Garrone, R.; Lethias, C. Cell adhesion to tenascin-X mapping of cell adhesion sites and identification of integrin receptors. Eur. J. Biochem. 1999, 263, 840–848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alcaraz, L.B.; Exposito, J.Y.; Chuvin, N.; Pommier, R.M.; Cluzel, C.; Martel, S.; Sentis, S.; Bartholin, L.; Lethias, C.; Valcourt, U. Tenascin-X promotes epithelial-to-mesenchymal transition by activating latent TGF-beta. J. Cell Biol. 2014, 205, 409–428. [Google Scholar] [CrossRef] [Green Version]
- Rios, H.; Koushik, S.V.; Wang, H.; Wang, J.; Zhou, H.M.; Lindsley, A.; Rogers, R.; Chen, Z.; Maeda, M.; Kruzynska-Frejtag, A.; et al. periostin null mice exhibit dwarfism, incisor enamel defects, and an early-onset periodontal disease-like phenotype. Mol. Cell Biol. 2005, 25, 11131–11144. [Google Scholar] [CrossRef] [Green Version]
- Conway, S.J.; Izuhara, K.; Kudo, Y.; Litvin, J.; Markwald, R.; Ouyang, G.; Arron, J.R.; Holweg, C.T.; Kudo, A. The role of periostin in tissue remodeling across health and disease. Cell Mol. Life Sci. 2014, 71, 1279–1288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elliott, C.G.; Wang, J.; Guo, X.; Xu, S.w.; Eastwood, M.; Guan, J.; Leask, A.; Conway, S.J.; Hamilton, D.W. Periostin modulates myofibroblast differentiation during full-thickness cutaneous wound repair. J. Cell Sci. 2012, 125, 121–132. [Google Scholar] [CrossRef] [Green Version]
- Shimazaki, M.; Nakamura, K.; Kii, I.; Kashima, T.; Amizuka, N.; Li, M.; Saito, M.; Fukuda, K.; Nishiyama, T.; Kitajima, S.; et al. Periostin is essential for cardiac healing after acute myocardial infarction. J. Exp. Med. 2008, 205, 295–303. [Google Scholar] [CrossRef] [PubMed]
- Lorts, A.; Schwanekamp, J.A.; Baudino, T.A.; McNally, E.M.; Molkentin, J.D. Deletion of periostin reduces muscular dystrophy and fibrosis in mice by modulating the transforming growth factor-beta pathway. Proc. Natl. Acad. Sci. USA 2012, 109, 10978–10983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Norris, R.A.; Damon, B.; Mironov, V.; Kasyanov, V.; Ramamurthi, A.; Moreno-Rodriguez, R.; Trusk, T.; Potts, J.D.; Goodwin, R.L.; Davis, J.; et al. Periostin regulates collagen fibrillogenesis and the biomechanical properties of connective tissues. J. Cell Biochem. 2007, 101, 695–711. [Google Scholar] [CrossRef] [Green Version]
- Egbert, M.; Ruetze, M.; Sattler, M.; Wenck, H.; Gallinat, S.; Lucius, R.; Weise, J.M. The matricellular protein periostin contributes to proper collagen function and is downregulated during skin aging. J. Derm. Sci. 2014, 73, 40–48. [Google Scholar] [CrossRef]
- Couchman, J.R.; Gopal, S.; Lim, H.C.; Norgaard, S.; Multhaupt, H.A. Syndecans: From peripheral coreceptors to mainstream regulators of cell behaviour. Int. J. Exp. Pathol. 2015, 96, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Multhaupt, H.A.; Leitinger, B.; Gullberg, D.; Couchman, J.R. Extracellular matrix component signaling in cancer. Adv. Drug Deliv. Rev. 2016, 97, 28–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, Y.; Chung, H.; Jung, H.; Couchman, J.R.; Oh, E.S. Syndecans as cell surface receptors: Unique structure equates with functional diversity. Matrix Biol. 2011, 30, 93–99. [Google Scholar] [CrossRef]
- Chronopoulos, A.; Thorpe, S.D.; Cortes, E.; Lachowski, D.; Rice, A.J.; Mykuliak, V.V.; Rog, T.; Lee, D.A.; Hytonen, V.P.; Del Rio Hernandez, A.E. Syndecan-4 tunes cell mechanics by activating the kindlin-integrin-RhoA pathway. Nat. Mater. 2020, 19, 669–678. [Google Scholar] [CrossRef] [PubMed]
- Ihrcke, N.S.; Platt, J.L. Shedding of heparan sulfate proteoglycan by stimulated endothelial cells: Evidence for proteolysis of cell-surface molecules. J. Cell Physiol. 1996, 168, 625–637. [Google Scholar] [CrossRef]
- Manon-Jensen, T.; Multhaupt, H.A.; Couchman, J.R. Mapping of matrix metalloproteinase cleavage sites on syndecan-1 and syndecan-4 ectodomains. Febs. J. 2013, 280, 2320–2331. [Google Scholar] [CrossRef]
- Choi, S.; Kim, J.Y.; Park, J.H.; Lee, S.T.; Han, I.O.; Oh, E.S. The matrix metalloproteinase-7 regulates the extracellular shedding of syndecan-2 from colon cancer cells. Biochem. Biophys. Res. Commun. 2012, 417, 1260–1264. [Google Scholar] [CrossRef] [PubMed]
- Manon-Jensen, T.; Itoh, Y.; Couchman, J.R. Proteoglycans in health and disease: The multiple roles of syndecan shedding. FEBS J. 2010, 277, 3876–3889. [Google Scholar] [CrossRef]
- Stepp, M.A.; Daley, W.P.; Bernstein, A.M.; Pal-Ghosh, S.; Tadvalkar, G.; Shashurin, A.; Palsen, S.; Jurjus, R.A.; Larsen, M. Syndecan-1 regulates cell migration and fibronectin fibril assembly. Exp. Cell Res. 2010, 316, 2322–2339. [Google Scholar] [CrossRef] [Green Version]
- Vuoriluoto, K.; Jokinen, J.; Kallio, K.; Salmivirta, M.; Heino, J.; Ivaska, J. Syndecan-1 supports integrin alpha2beta1-mediated adhesion to collagen. Exp. Cell Res. 2008, 314, 3369–3381. [Google Scholar] [CrossRef] [PubMed]
- Sottile, J.; Hocking, D.C.; Langenbach, K.J. Fibronectin polymerization stimulates cell growth by RGD-dependent and -independent mechanisms. J. Cell Sci. 2000, 113 Pt 23, 4287–4299. [Google Scholar]
- Gubbiotti, M.A.; Vallet, S.D.; Ricard-Blum, S.; Iozzo, R.V. Decorin interacting network: A comprehensive analysis of decorin-binding partners and their versatile functions. Matrix Biol. 2016, 55, 7–21. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Birk, D.E. The regulatory roles of small leucine-rich proteoglycans in extracellular matrix assembly. FEBS J. 2013, 280, 2120–2137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, D.; Smith, L.R.; Khandekar, G.; Patel, P.; Yu, C.K.; Zhang, K.; Chen, C.S.; Han, L.; Wells, R.G. Distinct effects of different matrix proteoglycans on collagen fibrillogenesis and cell-mediated collagen reorganization. Sci. Rep. 2020, 10, 19065. [Google Scholar] [CrossRef]
- Oldberg, A.; Kalamajski, S.; Salnikov, A.V.; Stuhr, L.; Morgelin, M.; Reed, R.K.; Heldin, N.E.; Rubin, K. Collagen-binding proteoglycan fibromodulin can determine stroma matrix structure and fluid balance in experimental carcinoma. Proc. Natl. Acad. Sci. USA 2007, 104, 13966–13971. [Google Scholar] [CrossRef] [Green Version]
- Seidler, D.G.; Faiyaz-Ul-Haque, M.; Hansen, U.; Yip, G.W.; Zaidi, S.H.; Teebi, A.S.; Kiesel, L.; Gotte, M. Defective glycosylation of decorin and biglycan, altered collagen structure, and abnormal phenotype of the skin fibroblasts of an Ehlers-Danlos syndrome patient carrying the novel Arg270Cys substitution in galactosyltransferase I (beta4GalT-7). J. Mol. Med. 2006, 84, 583–594. [Google Scholar] [CrossRef]
- Hildebrand, A.; Romaris, M.; Rasmussen, L.M.; Heinegard, D.; Twardzik, D.R.; Border, W.A.; Ruoslahti, E. Interaction of the small interstitial proteoglycans biglycan, decorin and fibromodulin with transforming growth factor beta. Biochem. J. 1994, 302 Pt 2, 527–534. [Google Scholar] [CrossRef]
- Boyd, N.A.; Bradwell, A.R.; Thompson, R.A. Quantitation of vitronectin in serum: Evaluation of its usefulness in routine clinical practice. J. Clin. Pathol. 1993, 46, 1042–1045. [Google Scholar] [CrossRef] [Green Version]
- Burgi, J.; Abrami, L.; Castanon, I.; Abriata, L.A.; Kunz, B.; Yan, S.E.; Lera, M.; Unger, S.; Superti-Furga, A.; Peraro, M.D.; et al. Ligand Binding to the Collagen VI Receptor Triggers a Talin-to-RhoA Switch that Regulates Receptor Endocytosis. Dev. Cell 2020, 53, 418–430.e414. [Google Scholar] [CrossRef]
- Schwarzbauer, J.E.; DeSimone, D.W. Fibronectins, their fibrillogenesis, and in vivo functions. Cold Spring Harb Perspect Biol. 2011, 3. [Google Scholar] [CrossRef] [Green Version]
- Pankov, R.; Cukierman, E.; Katz, B.Z.; Matsumoto, K.; Lin, D.C.; Lin, S.; Hahn, C.; Yamada, K.M. Integrin dynamics and matrix assembly: Tensin-dependent translocation of alpha(5)beta(1) integrins promotes early fibronectin fibrillogenesis. J. Cell Biol. 2000, 148, 1075–1090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCleverty, C.J.; Lin, D.C.; Liddington, R.C. Structure of the PTB domain of tensin1 and a model for its recruitment to fibrillar adhesions. Protein Sci. 2007, 16, 1223–1229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Legate, K.R.; Fassler, R. Mechanisms that regulate adaptor binding to beta-integrin cytoplasmic tails. J. Cell Sci. 2009, 122, 187–198. [Google Scholar] [CrossRef] [Green Version]
- Zamir, E.; Katz, B.Z.; Aota, S.; Yamada, K.M.; Geiger, B.; Kam, Z. Molecular diversity of cell-matrix adhesions. J. Cell Sci. 1999, 112 Pt 11, 1655–1669. [Google Scholar]
- Filla, M.S.; Dimeo, K.D.; Tong, T.; Peters, D.M. Disruption of fibronectin matrix affects type IV collagen, fibrillin and laminin deposition into extracellular matrix of human trabecular meshwork (HTM) cells. Exp. Eye Res. 2017, 165, 7–19. [Google Scholar] [CrossRef] [PubMed]
- Georgiadou, M.; Lilja, J.; Jacquemet, G.; Guzman, C.; Rafaeva, M.; Alibert, C.; Yan, Y.; Sahgal, P.; Lerche, M.; Manneville, J.B.; et al. AMPK negatively regulates tensin-dependent integrin activity. J. Cell Biol. 2017, 216, 1107–1121. [Google Scholar] [CrossRef] [Green Version]
- Stanchi, F.; Grashoff, C.; Nguemeni Yonga, C.F.; Grall, D.; Fassler, R.; Van Obberghen-Schilling, E. Molecular dissection of the ILK-PINCH-parvin triad reveals a fundamental role for the ILK kinase domain in the late stages of focal-adhesion maturation. J. Cell Sci. 2009, 122, 1800–1811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goreczny, G.J.; Forsythe, I.J.; Turner, C.E. Hic-5 regulates fibrillar adhesion formation to control tumor extracellular matrix remodeling through interaction with tensin1. Oncogene 2018, 37, 1699–1713. [Google Scholar] [CrossRef]
- Sundararaman, A.; Fukushima, Y.; Norman, J.C.; Uemura, A.; Mellor, H. RhoJ Regulates alpha5beta1 Integrin Trafficking to Control Fibronectin Remodeling during Angiogenesis. Curr. Biol. 2020, 30, 2146–2155.e2145. [Google Scholar] [CrossRef] [PubMed]
- Sundararaman, A.; Mellor, H. A functional antagonism between RhoJ and Cdc42 regulates fibronectin remodelling during angiogenesis. Small Gtpases. 2020, 1–5. [Google Scholar] [CrossRef]
- Fogerty, F.J.; Mosher, D.F. Mechanisms for organization of fibronectin matrix. Cell Differ. Dev. 1990, 32, 439–450. [Google Scholar] [CrossRef]
- Danen, E.H.; Sonneveld, P.; Brakebusch, C.; Fassler, R.; Sonnenberg, A. The fibronectin-binding integrins alpha5beta1 and alphavbeta3 differentially modulate RhoA-GTP loading, organization of cell matrix adhesions, and fibronectin fibrillogenesis. J. Cell Biol. 2002, 159, 1071–1086. [Google Scholar] [CrossRef] [Green Version]
- Lohikangas, L.; Gullberg, D.; Johansson, S. Assembly of laminin polymers is dependent on beta1-integrins. Exp. Cell Res. 2001, 265, 135–144. [Google Scholar] [CrossRef]
- Gonzalez, A.M.; Gonzales, M.; Herron, G.S.; Nagavarapu, U.; Hopkinson, S.B.; Tsuruta, D.; Jones, J.C. Complex interactions between the laminin alpha 4 subunit and integrins regulate endothelial cell behavior in vitro and angiogenesis in vivo. Proc. Natl. Acad. Sci. USA 2002, 99, 16075–16080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henry, M.D.; Satz, J.S.; Brakebusch, C.; Costell, M.; Gustafsson, E.; Fassler, R.; Campbell, K.P. Distinct roles for dystroglycan, beta1 integrin and perlecan in cell surface laminin organization. J. Cell Sci. 2001, 114, 1137–1144. [Google Scholar]
- Mayer, U. Integrins: Redundant or important players in skeletal muscle? J. Biol. Chem. 2003, 278, 14587–14590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gawlik, K.I.; Durbeej, M. Deletion of integrin alpha7 subunit does not aggravate the phenotype of laminin alpha2 chain-deficient mice. Sci. Rep. 2015, 5, 13916. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Van Den Diepstraten, C.; D’Souza, S.J.; Chan, B.M.; Pickering, J.G. Vascular smooth muscle cells orchestrate the assembly of type I collagen via alpha2beta1 integrin, RhoA, and fibronectin polymerization. Am. J. Pathol. 2003, 163, 1045–1056. [Google Scholar] [CrossRef]
- Velling, T.; Risteli, J.; Wennerberg, K.; Mosher, D.F.; Johansson, S. Polymerization of type I and III collagens is dependent on fibronectin and enhanced by integrins a11b1 and a2b1. J. Biol. Chem. 2002, 277, 37377–37381. [Google Scholar] [CrossRef] [Green Version]
- Ruggiero, F.; Comte, J.; Cabanas, C.; Garrone, R. Structural requirements for alpha 1 beta 1 and alpha 2 beta 1 integrin mediated cell adhesion to collagen V. J. Cell Sci. 1996, 109 Pt 7, 1865–1874. [Google Scholar]
- Popova, S.N.; Rodriguez-Sanchez, B.; Liden, A.; Betsholtz, C.; Van Den Bos, T.; Gullberg, D. The mesenchymal alpha11beta1 integrin attenuates PDGF-BB-stimulated chemotaxis of embryonic fibroblasts on collagens. Dev. Biol. 2004, 270, 427–442. [Google Scholar] [CrossRef]
- Yurchenco, P.D. Integrating Activities of Laminins that Drive Basement Membrane Assembly and Function. Curr. Top. Membr. 2015, 76, 1–30. [Google Scholar] [CrossRef]
- Takada, Y.; Hemler, M.E. The primary structure of the VLA-2/collagen receptor a2 subunit (platelet GPIa): Homology to other integrins and the presence of a possible collagen-binding domain. J. Cell Biol. 1989, 109, 397–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elices, M.J.; Hemler, M.E. The human integrin VLA-2 is a collagen receptor on some cells and a collagen/laminin receptor on others. Proc. Natl. Acad. Sci. USA 1989, 86, 9906–9910. [Google Scholar] [CrossRef] [Green Version]
- Trelstad, R.L.; Hayashi, K. Tendon collagen fibrillogenesis: Intracellular subassemblies and cell surface changes associated with fibril growth. Dev. Biol. 1979, 71, 228–242. [Google Scholar] [CrossRef]
- Birk, D.E.; Trelstad, R.L. Extracellular compartments in matrix morphogenesis: Collagen fibril, bundle, and lamellar formation by corneal fibroblasts. J. Cell Biol. 1984, 99, 2024–2033. [Google Scholar] [CrossRef] [PubMed]
- Canty, E.G.; Lu, Y.; Meadows, R.S.; Shaw, M.K.; Holmes, D.F.; Kadler, K.E. Coalignment of plasma membrane channels and protrusions (fibripositors) specifies the parallelism of tendon. J. Cell Biol. 2004, 165, 553–563. [Google Scholar] [CrossRef] [Green Version]
- Kalson, N.S.; Lu, Y.; Taylor, S.H.; Starborg, T.; Holmes, D.F.; Kadler, K.E. A structure-based extracellular matrix expansion mechanism of fibrous tissue growth. Elife 2015, 4. [Google Scholar] [CrossRef] [PubMed]
- Saunders, J.T.; Schwarzbauer, J.E. Fibronectin matrix as a scaffold for procollagen proteinase binding and collagen processing. Mol. Biol. Cell 2019, 30, 2218–2226. [Google Scholar] [CrossRef]
- Primac, I.; Maquoi, E.; Blacher, S.; Heljasvaara, R.; Van Deun, J.; Smeland, H.Y.; Canale, A.; Louis, T.; Stuhr, L.; Sounni, N.E.; et al. Stromal integrin alpha11 regulates PDGFR-beta signaling and promotes breast cancer progression. J. Clin. Investig. 2019, 130, 4609–4628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navab, R.; Strumpf, D.; To, C.; Pasko, E.; Kim, K.S.; Park, C.J.; Hai, J.; Liu, J.; Jonkman, J.; Barczyk, M.; et al. Integrin alpha11beta1 regulates cancer stromal stiffness and promotes tumorigenicity and metastasis in non-small cell lung cancer. Oncogene 2016, 35, 1899–1908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulz, J.N.; Zeltz, C.; Sorensen, I.W.; Barczyk, M.; Carracedo, S.; Hallinger, R.; Niehoff, A.; Eckes, B.; Gullberg, D. Reduced granulation tissue and wound strength in the absence of alpha11beta1 integrin. J. Investig. Derm. 2015, 135, 1435–1444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munksgaard Thoren, M.; Chmielarska Masoumi, K.; Krona, C.; Huang, X.; Kundu, S.; Schmidt, L.; Forsberg-Nilsson, K.; Floyd Keep, M.; Englund, E.; Nelander, S.; et al. Integrin alpha10, a Novel Therapeutic Target in Glioblastoma, Regulates Cell Migration, Proliferation, and Survival. Cancers 2019, 11, 587. [Google Scholar] [CrossRef] [Green Version]
- Franco-Barraza, J.; Francescone, R.; Luong, T.; Shah, N.; Madhani, R.; Cukierman, G.; Dulaimi, E.; Devarajan, K.; Egleston, B.L.; Nicolas, E.; et al. Matrix-regulated integrin alphavbeta5 maintains alpha5beta1-dependent desmoplastic traits prognostic of neoplastic recurrence. Elife 2017, 6. [Google Scholar] [CrossRef]
- Ledger, P.W.; Uchida, N.; Tanzer, M.L. Immunocytochemical localization of procollagen and fibronectin in human fibroblasts: Effects of the monovalent ionophore, monensin. J. Cell Biol. 1980, 87, 663–671. [Google Scholar] [CrossRef] [PubMed]
- McDonald, J.A.; Kelley, D.G.; Broekelmann, T.J. Role of fibronectin in collagen deposition: Fab’ to the gelatin-binding domain of fibronectin inhibits both fibronectin and collagen organization in fibroblast extracellular matrix. J. Cell Biol. 1982, 92, 485–492. [Google Scholar] [CrossRef]
- Prockop, D.J.; Fertala, A. Inhibition of the self-assembly of collagen I into fibrils with synthetic peptides. Demonstration that assembly is driven by specific binding sites on the monomers. J. Biol. Chem. 1998, 273, 15598–15604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dzamba, B.J.; Wu, H.; Jaenisch, R.; Peters, D.M. Fibronectin binding site in type I collagen regulates fibronectin fibril formation. J. Cell Biol. 1993, 121, 1165–1172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sottile, J.; Shi, F.; Rublyevska, I.; Chiang, H.Y.; Lust, J.; Chandler, J. Fibronectin-dependent collagen I deposition modulates the cell response to fibronectin. Am. J. Physiol. Cell Physiol. 2007, 293, C1934–C1946. [Google Scholar] [CrossRef] [Green Version]
- Tomasini-Johansson, B.R.; Kaufman, N.R.; Ensenberger, M.G.; Ozeri, V.; Hanski, E.; Mosher, D.F. A 49-residue peptide from adhesin F1 of Streptococcus pyogenes inhibits fibronectin matrix assembly. J. Biol. Chem. 2001, 276, 23430–23439. [Google Scholar] [CrossRef] [Green Version]
- Sottile, J.; Hocking, D.C.; Swiatek, P.J. Fibronectin matrix assembly enhances adhesion-dependent cell growth. J. Cell Sci. 1998, 111 Pt 19, 2933–2943. [Google Scholar]
- Holtkotter, O.; Nieswandt, B.; Smyth, N.; Muller, W.; Hafner, M.; Schulte, V.; Krieg, T.; Eckes, B. Integrin alpha 2-deficient mice develop normally, are fertile, but display partially defective platelet interaction with collagen. J. Biol. Chem. 2002, 277, 10789–10794. [Google Scholar] [CrossRef] [Green Version]
- Sottile, J.; Hocking, D.C. Fibronectin polymerization regulates the composition and stability of extracellular matrix fibrils and cell-matrix adhesions. Mol. Biol. Cell 2002, 13, 3546–3559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hocking, D.C.; Sottile, J.; Langenbach, K.J. Stimulation of integrin-mediated cell contractility by fibronectin polymerization. J. Biol. Chem. 2000, 275, 10673–10682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moriya, K.; Bae, E.; Honda, K.; Sakai, K.; Sakaguchi, T.; Tsujimoto, I.; Kamisoyama, H.; Keene, D.R.; Sasaki, T.; Sakai, T. A fibronectin-independent mechanism of collagen fibrillogenesis in adult liver remodeling. Gastroenterology 2011, 140, 1653–1663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, K.; Pritchett, J.; Llewellyn, J.; Mullan, A.F.; Athwal, V.S.; Dobie, R.; Harvey, E.; Zeef, L.; Farrow, S.; Streuli, C.; et al. PAK proteins and YAP-1 signalling downstream of integrin beta-1 in myofibroblasts promote liver fibrosis. Nat. Commun. 2016, 7, 12502. [Google Scholar] [CrossRef] [PubMed]
- Gardner, H. Integrin alpha1beta1. Adv. Exp. Med. Biol. 2014, 819, 21–39. [Google Scholar] [CrossRef]
- Gardner, H.; Kreidberg, J.; Koteliansky, V.; Jaenisch, R. Deletion of integrin a1 by homologous recombination permits normal murine development but gives rise to a specific deficit in cell adhesion. Dev. Biol. 1996, 175, 301–313. [Google Scholar] [CrossRef]
- Camper, L.; Holmvall, K.; Wangnerud, C.; Aszodi, A.; Lundgren-Akerlund, E. Distribution of the collagen-binding integrin alpha10beta1 during mouse development. Cell Tissue Res. 2001, 306, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Kyostila, K.; Lappalainen, A.K.; Lohi, H. Canine chondrodysplasia caused by a truncating mutation in collagen-binding integrin alpha subunit 10. PLoS ONE 2013, 8, e75621. [Google Scholar] [CrossRef] [Green Version]
- Vogel, W.; Gish, G.D.; Alves, F.; Pawson, T. The discoidin domain receptor tyrosine kinases are activated by collagen. Mol. Cell 1997, 1, 13–23. [Google Scholar] [CrossRef]
- Coelho, N.M.; Arora, P.D.; van Putten, S.; Boo, S.; Petrovic, P.; Lin, A.X.; Hinz, B.; McCulloch, C.A. Discoidin Domain Receptor 1 Mediates Myosin-Dependent Collagen Contraction. Cell Rep. 2017, 18, 1774–1790. [Google Scholar] [CrossRef] [Green Version]
- Bayer, S.V.; Grither, W.R.; Brenot, A.; Hwang, P.Y.; Barcus, C.E.; Ernst, M.; Pence, P.; Walter, C.; Pathak, A.; Longmore, G.D. DDR2 controls breast tumor stiffness and metastasis by regulating integrin mediated mechanotransduction in CAFs. Elife 2019, 8. [Google Scholar] [CrossRef]
- Zoppi, N.; Gardella, R.; De Paepe, A.; Barlati, S.; Colombi, M. Human fibroblasts with mutations in COL5A1 and COL3A1 genes do not organize collagens and fibronectin in the extracellular matrix, down-regulate alpha2beta1 integrin, and recruit alphavbeta3 Instead of alpha5beta1 integrin. J. Biol. Chem. 2004, 279, 18157–18168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blaschke, U.K.; Eikenberry, E.F.; Hulmes, D.J.; Galla, H.J.; Bruckner, P. Collagen XI nucleates self-assembly and limits lateral growth of cartilage fibrils. J. Biol. Chem. 2000, 275, 10370–10378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansen, U.; Bruckner, P. Macromolecular specificity of collagen fibrillogenesis: Fibrils of collagens I and XI contain a heterotypic alloyed core and a collagen I sheath. J. Biol. Chem. 2003, 278, 37352–37359. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Lacerda, D.A.; Warman, M.L.; Beier, D.R.; Yoshioka, H.; Ninomiya, Y.; Oxford, J.T.; Morris, N.P.; Andrikopoulos, K.; Ramirez, F.; et al. A fibrillar collagen gene, Col11a1, is essential for skeletal morphogenesis. Cell 1995, 80, 423–430. [Google Scholar] [CrossRef] [Green Version]
- Wenstrup, R.J.; Florer, J.B.; Brunskill, E.W.; Bell, S.M.; Chervoneva, I.; Birk, D.E. Type V collagen controls the initiation of collagen fibril assembly. J. Biol. Chem. 2004, 279, 53331–53337. [Google Scholar] [CrossRef] [Green Version]
- Wenstrup, R.J.; Florer, J.B.; Davidson, J.M.; Phillips, C.L.; Pfeiffer, B.J.; Menezes, D.W.; Chervoneva, I.; Birk, D.E. Murine model of the Ehlers-Danlos syndrome. col5a1 haploinsufficiency disrupts collagen fibril assembly at multiple stages. J. Biol. Chem. 2006, 281, 12888–12895. [Google Scholar] [CrossRef] [Green Version]
- Sun, M.; Chen, S.; Adams, S.M.; Florer, J.B.; Liu, H.; Kao, W.W.; Wenstrup, R.J.; Birk, D.E. Collagen V is a dominant regulator of collagen fibrillogenesis: Dysfunctional regulation of structure and function in a corneal-stroma-specific Col5a1-null mouse model. J. Cell Sci. 2011, 124, 4096–4105. [Google Scholar] [CrossRef] [Green Version]
- Chanut-Delalande, H.; Fichard, A.; Bernocco, S.; Garrone, R.; Hulmes, D.J.; Ruggiero, F. Control of heterotypic fibril formation by collagen Ⅴ is determined by chain stoichiometry. J. Biol. Chem. 2001, 276, 24352–24359. [Google Scholar] [CrossRef] [Green Version]
- Espana, E.M.; Birk, D.E. Composition, structure and function of the corneal stroma. Exp. Eye Res. 2020, 198, 108137. [Google Scholar] [CrossRef] [PubMed]
- Nicolae, C.; Ko, Y.P.; Miosge, N.; Niehoff, A.; Studer, D.; Enggist, L.; Hunziker, E.B.; Paulsson, M.; Wagener, R.; Aszodi, A. Abnormal collagen fibrils in cartilage of matrilin-1/matrilin-3-deficient mice. J. Biol. Chem. 2007, 282, 22163–22175. [Google Scholar] [CrossRef] [Green Version]
- Halasz, K.; Kassner, A.; Morgelin, M.; Heinegard, D. COMP acts as a catalyst in collagen fibrillogenesis. J. Biol. Chem. 2007, 282, 31166–31173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomer, D.; Munshi, S.; Alexander, B.E.; French, B.; Vedula, P.; House, A.; Guvendiren, M.; Kashina, A.; Schwarzbauer, J.E.; Astrof, S. New mechanism of of fibronectin fibril assembly revealed by live imaging and super-resolution microscopy. bioRXiv 2020. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Musiime, M.; Chang, J.; Hansen, U.; Kadler, K.E.; Zeltz, C.; Gullberg, D. Collagen Assembly at the Cell Surface: Dogmas Revisited. Cells 2021, 10, 662. https://doi.org/10.3390/cells10030662
Musiime M, Chang J, Hansen U, Kadler KE, Zeltz C, Gullberg D. Collagen Assembly at the Cell Surface: Dogmas Revisited. Cells. 2021; 10(3):662. https://doi.org/10.3390/cells10030662
Chicago/Turabian StyleMusiime, Moses, Joan Chang, Uwe Hansen, Karl E. Kadler, Cédric Zeltz, and Donald Gullberg. 2021. "Collagen Assembly at the Cell Surface: Dogmas Revisited" Cells 10, no. 3: 662. https://doi.org/10.3390/cells10030662
APA StyleMusiime, M., Chang, J., Hansen, U., Kadler, K. E., Zeltz, C., & Gullberg, D. (2021). Collagen Assembly at the Cell Surface: Dogmas Revisited. Cells, 10(3), 662. https://doi.org/10.3390/cells10030662