
Acyl–Acyl Carrier Protein Desaturases and Plant Biotic Interactions


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Synthesis of Monounsaturated Fatty Acids by Acyl–Acyl Carrier Protein (ACP) Desaturases
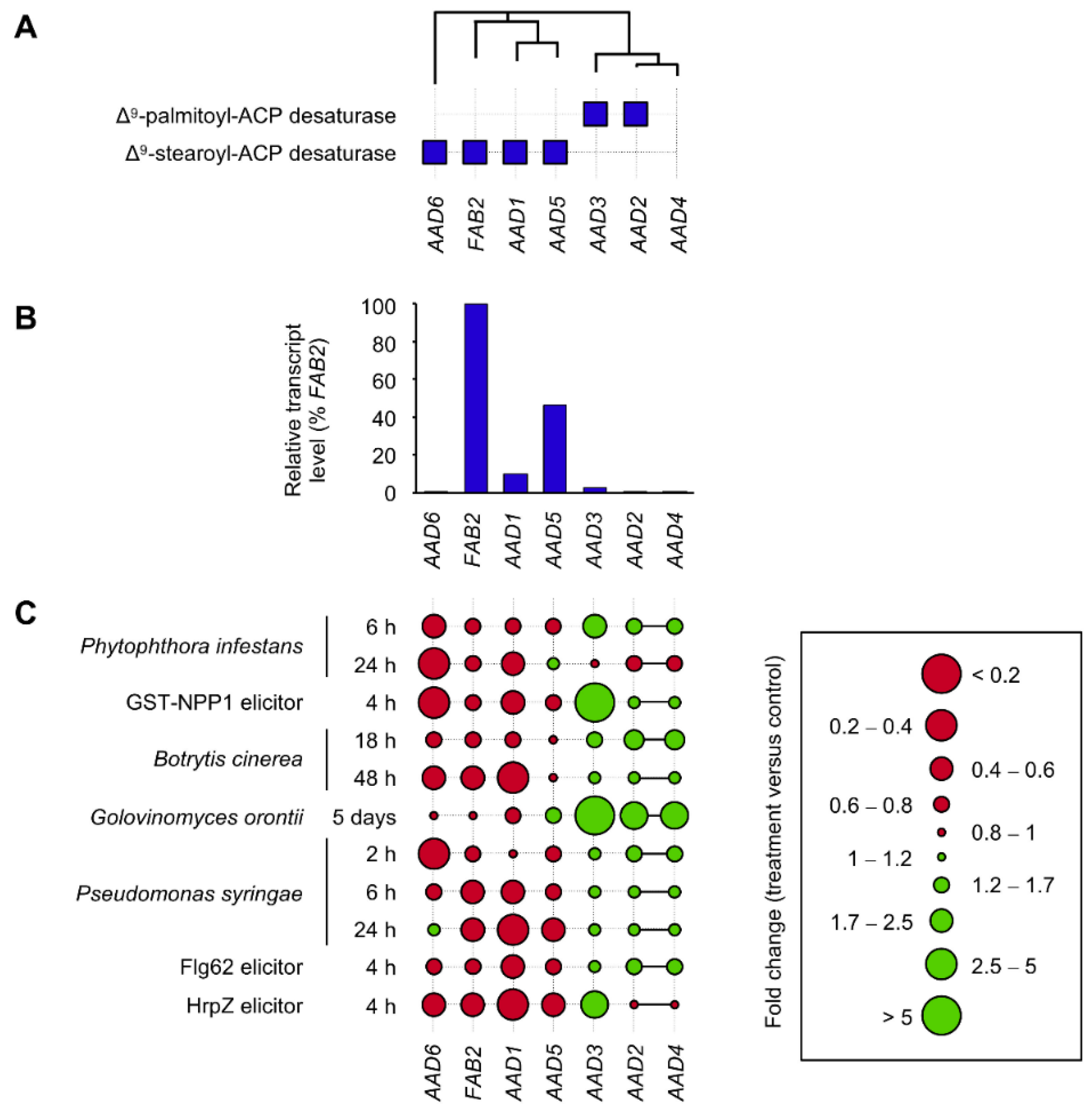
3. Transcriptional Responses of AAD Genes to Biotic Stress in Arabidopsis Leaves
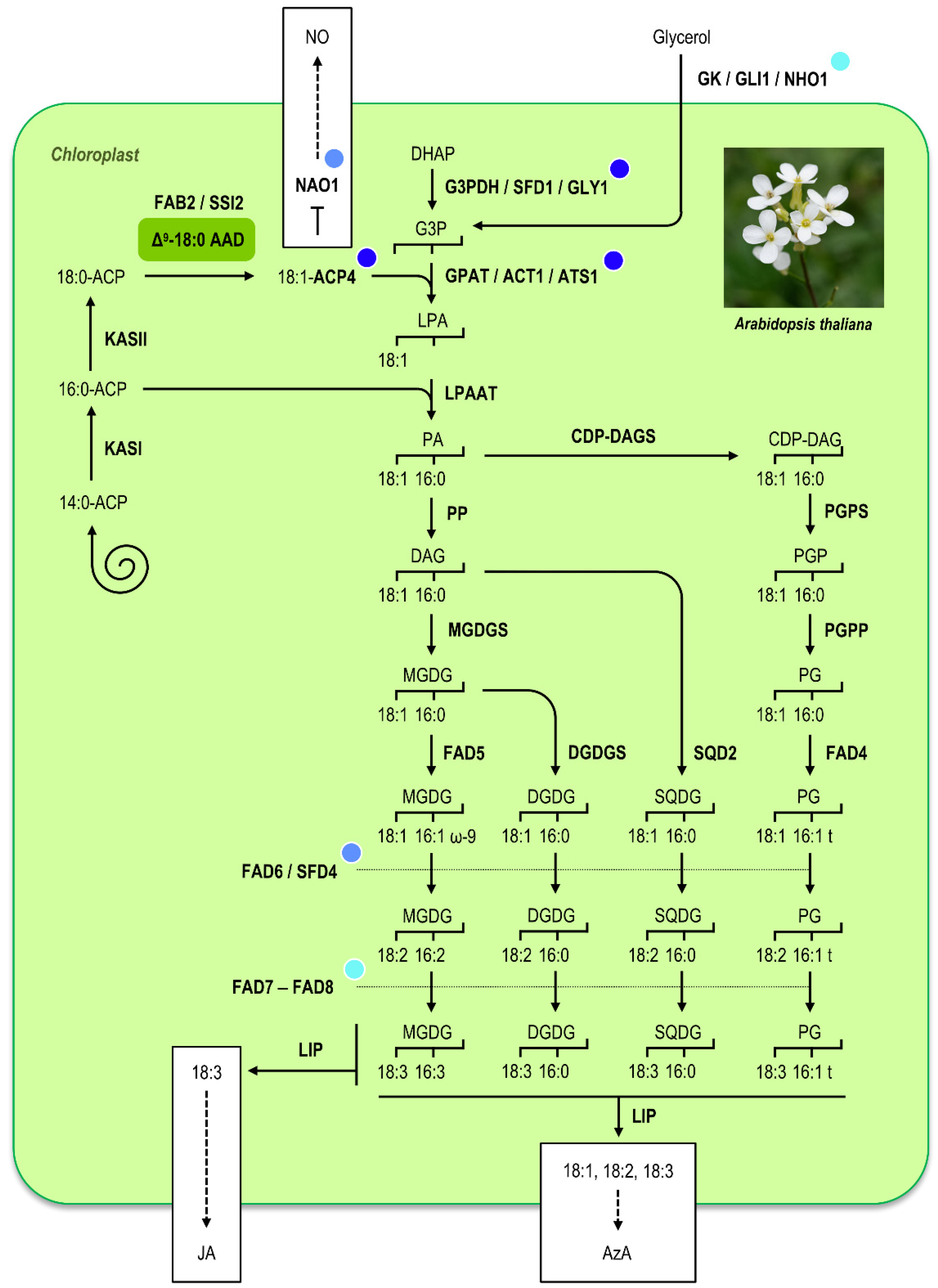
4. Δ9-Stearoyl–ACP Desaturases and Plant–Pathogen Interactions

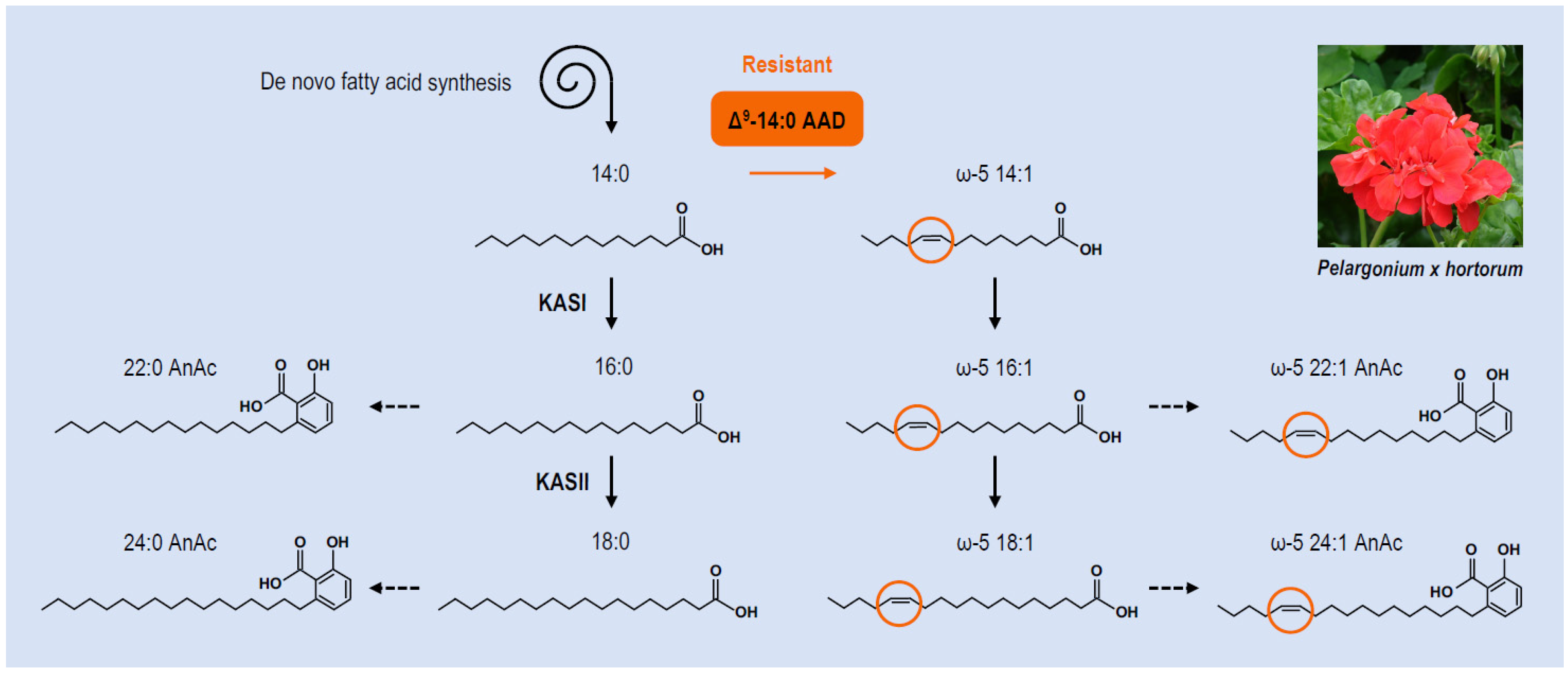
5. Synthesis of ω-Anacardic Acids and Resistance to Pests in Geranium
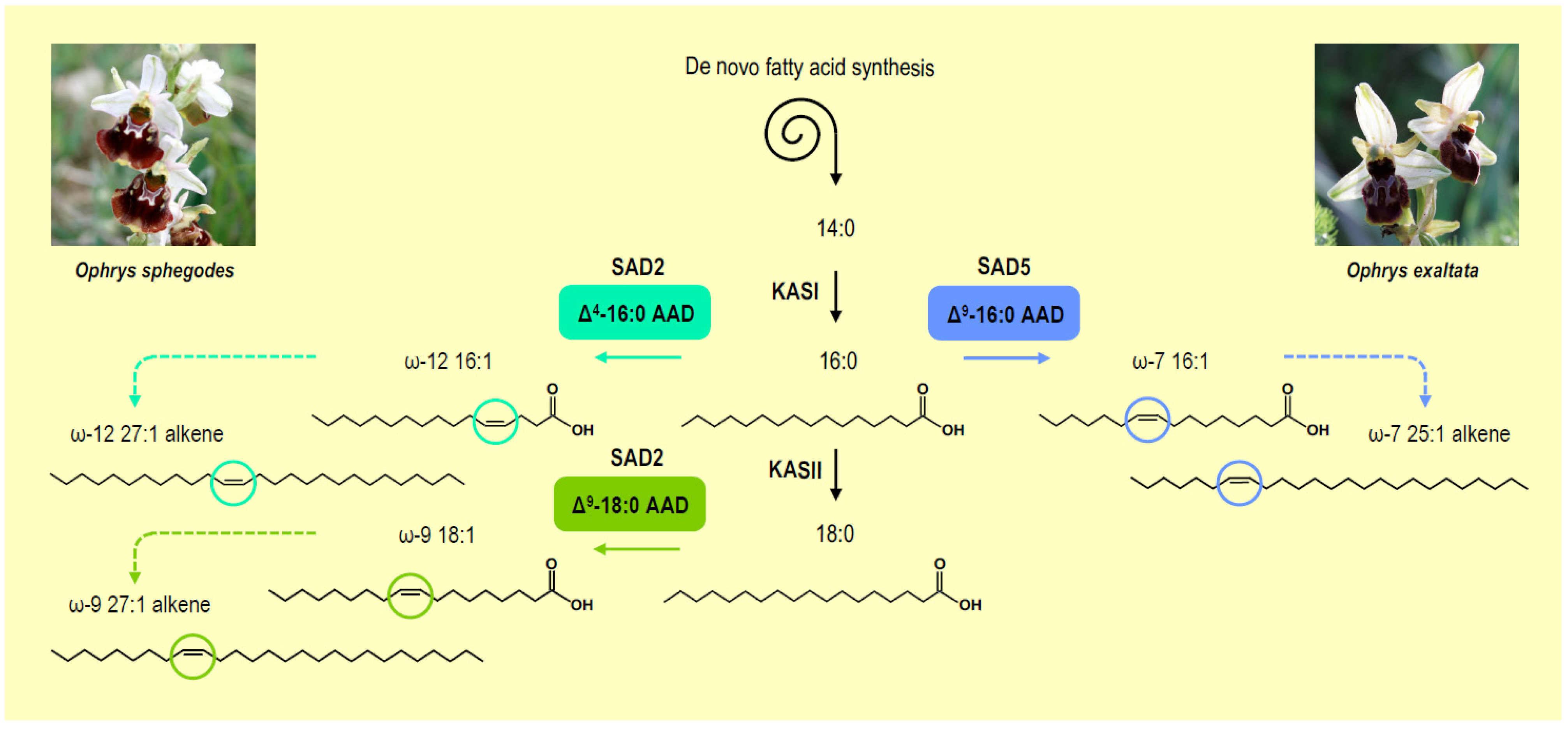
6. Synthesis of Alkenes and Attraction of Pollinators in Orchids
7. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Mamode Cassim, A.; Gouguet, P.; Gronnier, J.; Laurent, N.; Germain, V.; Grison, M.; Boutté, Y.; Gerbeau-Pissot, P.; Simon-Plas, F.; Mongrand, S. Plant lipids: Key players of plasma membrane organization and function. Prog. Lipid Res. 2019, 73, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Baud, S. Seeds as oil factories. Plant Reprod. 2018, 3, 213–235. [Google Scholar] [CrossRef]
- Miray, R.; Kazaz, S.; To, A.; Baud, S. Molecular control of oil metabolism in the endosperm of seeds. Int. J. Mol. Sci. 2021, 22, 1621. [Google Scholar] [CrossRef]
- Kunst, L.; Samuels, L. Plant cuticles shine: Advances in wax biosynthesis and export. Curr. Opin. Plant Biol. 2009, 12, 721–727. [Google Scholar] [CrossRef] [PubMed]
- Nandi, A.; Krothapalli, K.; Buseman, C.M.; Li, M.; Welti, R.; Enyedi, A.; Shah, J. Arabidopsis sfd mutants affect plastidic lipid composition and suppress dwarfing, cell death, and the enhanced disease resistance phenotypes resulting from the deficiency of a fatty acid desaturase. Plant Cell 2003, 15, 2383–2398. [Google Scholar] [CrossRef] [Green Version]
- Wasternack, C. Jasmonates: An update on biosynthesis, signal transduction and action in plant stress response, growth and development. Ann. Bot. 2007, 100, 681–697. [Google Scholar] [CrossRef] [Green Version]
- Bessire, M.; Chassot, C.; Jacquat, A.C.; Humphry, M.; Borel, S.; Petétot, J.M.; Métraux, J.P.; Nawrath, C. A permeable cuticle in Arabidopsis leads to a strong resistance to Botrytis cinerea. EMBO J. 2007, 26, 2158–2168. [Google Scholar] [CrossRef]
- Xia, Y.; Yu, K.; Navarre, D.; Seebold, K.; Kachroo, A.; Kachroo, P. The glabra1 mutation affects cuticle formation and plant responses to microbes. Plant Physiol. 2010, 154, 833–846. [Google Scholar] [CrossRef] [Green Version]
- Xia, Y.; Yu, K.; Gao, Q.M.; Wilson, E.V.; Navarre, D.; Kachroo, P.; Kachroo, A. Acyl CoA binding proteins are required for cuticle formation and plant responses to microbes. Front. Plant Sci. 2012, 3, 224. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.; Lim, G.H.; Kachroo, P. Transport of chemical signals in systemic acquired resistance. J. Integr. Plant Biol. 2017, 59, 336–344. [Google Scholar] [CrossRef] [PubMed]
- Upchurch, R.G. Fatty acid unsaturation, mobilization, and regulation in the response of plants to stress. Biotechnol. Lett. 2008, 30, 967–977. [Google Scholar] [CrossRef]
- Walley, J.W.; Kliebenstein, D.J.; Bostock, R.M.; Dehesh, K. Fatty acids and early detection of pathogens. Curr. Opin. Plant Biol. 2013, 16, 520–526. [Google Scholar] [CrossRef] [Green Version]
- Lim, G.-H.; Singhal, R.; Kachroo, A.; Kachroo, P. Fatty acid- and lipid-mediated signaling in plant defense. Annu. Rev. Phytopatol. 2017, 55, 505–536. [Google Scholar] [CrossRef]
- Yaeno, T.; Matsuda, O.; Iba, K. Role of chloroplast trienoic fatty acids in plant disease defense responses. Plant J. 2004, 40, 931–941. [Google Scholar] [CrossRef]
- Xue, H.Q.; Upchurch, R.G.; Kwanyuen, P. Relationships between oleic and linoleic acid content and seed colonization by Cercospora kikuchii and Diaporthe phaseolorum. Plant Dis. 2008, 92, 1038–1042. [Google Scholar] [CrossRef]
- Xing, J.; Chin, C.-K. Modification of fatty acids in eggplant affects its resistance to Verticillium dahlia. Physiol. Mol. Plant Pathol. 2000, 56, 217–225. [Google Scholar] [CrossRef]
- Wang, C.L.; Chin, C.K.; Ho, C.T.; Hwang, C.F.; Polashock, J.J.; Martin, C.E. Changes of fatty acids and fatty acid-derived flavor compounds by expressing the yeast ∆-9 desaturase gene in tomato enhances its resistance to powdery mildew. Physiol. Mol. Plant Physiol. 1998, 52, 371–383. [Google Scholar] [CrossRef]
- Harwood, J.L. Recent advances in the biosynthesis of plant fatty acids. Biochim. Biophys. Acta 1996, 1301, 7–56. [Google Scholar] [CrossRef]
- Troncoso-Ponce, M.A.; Nikovics, K.; Marchive, C.; Lepiniec, L.; Baud, S. New insights on the organization and regulation of the fatty acid biosynthetic network in the model higher plant Arabidopsis thaliana. Biochimie 2016, 120, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Turnham, E.; Northcote, D.H. Changes in the activity of acetyl-CoA carboxylase during rape-seed formation. Biochem. J. 1983, 212, 223–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, A.P.; Affleck, V.; Fawcett, T.; Slabas, A.R. Tandem affinity purification tagging of fatty acid biosynthetic enzymes in Synechocystis sp PCC6803 and Arabidopsis thaliana. J. Exp. Bot. 2006, 57, 1563–1571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimakata, T.; Stumpf, P.K. Purification and characterization of β-ketoacyl-[acyl-carrier-protein] reductase, β-hydroxyacyl-[acyl-carrier-protein] dehydrase, and enoyl-[acyl-carrier-protein] reductase from Spinacia oleracea leaves. Arch. Biochem. Biophys. 1982, 218, 77–91. [Google Scholar] [CrossRef]
- Jaworski, J.G.; Clough, R.C.; Barnum, S.R. A cerulenin insensitive short chain 3-ketoacyl-acyl carrier protein synthase in Spinacia oleracea leaves. Plant Physiol. 1989, 90, 41–44. [Google Scholar] [CrossRef] [Green Version]
- Ohlrogge, J.; Browse, J. Lipid biosynthesis. Plant Cell 1995, 7, 957–970. [Google Scholar]
- Salas, J.J.; Ohlrogge, J.B. Characterization of substrate specificity of plant FatA and FatB acyl-ACP thioesterases. Arch. Biochem. Biophys. 2002, 403, 25–34. [Google Scholar] [CrossRef]
- He, M.; Qin, C.-X.; Wang, X.; Ding, N.-Z. Plant unsaturated fatty acids: Biosynthesis and regulation. Front. Plant Sci. 2020, 11, 390. [Google Scholar] [CrossRef]
- Kachroo, A.; Shanklin, J.; Whittle, E.; Lapchyk, L.; Hildebrand, D.; Kachroo, P. The Arabidopsis stearoyl-acyl carrier protein-desaturase family and the contribution of leaf isoforms to oleic acid synthesis. Plant Mol. Biol. 2007, 63, 257–271. [Google Scholar] [CrossRef]
- Cahoon, E.B.; Shah, S.; Shanklin, J.; Browse, J. A determinant of substrate specificity predicted from the acyl-acyl carrier protein desaturase of developing cat’s claw seed. Plant Physiol. 1998, 117, 593–598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bryant, F.M.; Munoz-Azcarate, O.; Kelly, A.A.; Beaudoin, F.; Kurup, S.; Eastmond, P.J. ACYL-ACYL CARRIER PROTEIN DESATURASE2 and 3 are responsible for making omega-7 fatty acids in the Arabidopsis aleurone. Plant Physiol. 2016, 172, 154–162. [Google Scholar] [CrossRef]
- Troncoso-Ponce, M.A.; Barthole, G.; Tremblais, G.; To, A.; Miquel, M.; Lepiniec, L.; Baud, S. Transcriptional activation of two delta-9 palmitoyl-ACP desaturase genes by MYB115 and MYB118 is critical for biosynthesis of omega-7 monounsaturated fatty acids in the endosperm of Arabidopsis seeds. Plant Cell 2016, 28, 2666–2682. [Google Scholar] [CrossRef] [Green Version]
- Cahoon, E.B.; Cranmer, A.M.; Shanklin, J.; Ohlrogge, J.B. Δ6 Hexadecenoic acid is synthesized by the activity of a soluble Δ6 palmitoyl-acyl carrier protein desaturase in Thunbergia alata endosperm. J. Biol. Chem. 1994, 269, 27519–27526. [Google Scholar] [CrossRef]
- Cahoon, E.B.; Ohlrogge, J.B. Metabolic evidence for the involvement of a Δ4-palmitoyl-acyl carrier protein desaturase in petroselinic acid synthesis in coriander endosperm and transgenic tobacco cells. Plant Physiol. 1994, 104, 827–837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, G.A.; Scherer, D.E.; Foxall-Van Aken, S.; Kenny, J.W.; Young, H.L.; Shintani, D.K.; Kridl, J.C.; Knauf, V.C. Primary structures of the precursor and mature forms of stearoyl-acyl carrier protein desaturase from safflower embryos and requirement of ferredoxin for enzyme activity. Proc. Natl. Acad. Sci. USA 1991, 88, 2578–2582. [Google Scholar] [CrossRef] [Green Version]
- Shanklin, J.; Somerville, C. Stearoyl-acyl-carrier-protein desaturase from higher plants is structurally unrelated to the animal and fungal homologs. Proc. Natl. Acad. Sci. USA 1991, 88, 2510–2514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shanklin, J.; Guy, J.E.; Mishra, G.; Lindqvist, Y. Desaturases: Emerging models for understanding functional diversification of diiron-containing enzymes. J. Biol. Chem. 2009, 284, 18559–18563. [Google Scholar] [CrossRef] [Green Version]
- Fox, B.G.; Shanklin, J.; Somerville, C.; Münck, E. Stearoyl-acyl carrier protein Δ9 desaturase from Ricinus communis is a diiron-oxo protein. Proc. Natl. Acad. Sci. USA 1993, 90, 2486–2490. [Google Scholar] [CrossRef] [Green Version]
- Fox, B.G.; Lyle, K.S.; Rogge, C.E. Reactions of the diiron enzyme stearoyl-acyl carrier protein desaturase. Acc. Chem. Res. 2004, 37, 421–429. [Google Scholar] [CrossRef]
- Lindqvist, Y.; Huang, W.; Schneider, G.; Shanklin, J. Crystal structure of delta9 stearoyl-acyl carrier protein desaturase from castor seed and its relationship to other di-iron proteins. EMBO J. 1996, 15, 4081–4092. [Google Scholar] [CrossRef] [PubMed]
- Guy, J.E.; Whittle, E.; Kumaran, D.; Lindqvist, Y.; Shanklin, J. The crystal structure of the ivy ∆4–16:0-ACP desaturase reveals structural details of the oxidized active site and potential determinants of regioselectivity. J. Biol. Chem. 2007, 282, 19863–19871. [Google Scholar] [CrossRef] [Green Version]
- Shanklin, J.; Cahoon, E.B. Desaturation and related modifications of fatty acids. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1998, 49, 611–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Behrouzian, B.; Savile, C.K.; Dawson, B.; Buist, P.H.; Shanklin, J. Exploring the hydroxylation-dehydrogenation connection: Novel catalytic activity of castor stearoyl-ACP Δ9 desaturase. J. Am. Chem. Soc. 2002, 124, 3277–3283. [Google Scholar] [CrossRef]
- Cahoon, E.B.; Coughlan, S.J.; Shanklin, J. Characterization of a structurally and functionally diverged acyl-acyl carrier protein desaturase from milkweed seed. Plant Mol. Biol. 1997, 33, 1106–1110. [Google Scholar]
- Cahoon, E.B.; Lindqvist, Y.; Schneider, G.; Shanklin, J. Redesign of soluble fatty acid desaturases from plants for altered substrate specificity and double bond position. Proc. Natl. Acad. Sci. USA 1997, 94, 4872–4877. [Google Scholar] [CrossRef] [Green Version]
- Guy, E.J.; Whittle, E.; Moche, M.; Lengqvist, J.; Lindqvist, Y.; Shanklin, J. Remote control of regioselectivity in acyl-acyl carrier protein-desaturase. Proc. Natl. Acad. Sci. USA 2011, 108, 16594–16599. [Google Scholar] [CrossRef] [Green Version]
- Kachroo, P.; Shanklin, J.; Shah, J.; Whittle, E.J.; Klessig, D.F. A fatty acid desaturase modulates the activation of defense signaling pathways in plants. Proc. Natl. Acad. Sci. USA 2001, 98, 9448–9453. [Google Scholar] [CrossRef] [Green Version]
- Arabidopsis Genome Initiative. Analysis of the genome sequence of the flowering plant Arabidopsis thaliana. Nature 2000, 408, 796–815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, Y.; Xian, M.; Yang, J.; Xu, X.; Liu, W.; Li, L. Heterologous expression of stearoyl-acyl carrier protein desaturase (S-ACP-DES) from Arabidopsis thaliana in Escherichia coli. Protein Expr. Purif. 2010, 69, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Klinkenberg, J.; Faist, H.; Saupe, S.; Lambertz, S.; Krischke, M.; Stingl, N.; Fekete, A.; Mueller, M.J.; Feussner, I.; Hedrich, R.; et al. Two fatty acid desaturases, STEAROYL-ACYL CARRIER PROTEIN ∆9-DESATURASE6 and FATTY ACID DESATURASE3, are involved in drought and hypoxia stress signaling in Arabidopsis crown galls. Plant Physiol. 2014, 164, 570–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kazaz, S.; Barthole, G.; Domergue, F.; Ettaki, H.; To, A.; Vasselon, D.; De Vos, D.; Belcram, K.; Lepiniec, L.; Baud, S. Differential activation of partially redundant Δ9 stearoyl-ACP desaturase genes is critical for omega-9 monounsaturated fatty acid biosynthesis during seed development in Arabidopsis. Plant Cell 2020, 32, 3613–3637. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; He, F.; Zhang, Z. Large-scale transcriptome analysis reveals arabidopsis metabolic pathways are frequently influenced by different pathogens. Plant Mol. Biol. 2017, 94, 453–467. [Google Scholar] [CrossRef] [PubMed]
- Winter, D.; Vinegar, B.; Nahal, H.; Ammar, R.; Wilson, G.V.; Provart, N.J. An “Electronic Fluorescent Pictograph” browser for exploring and analyzing large-scale biological data sets. PLoS ONE 2007, 2, e718. [Google Scholar] [CrossRef] [PubMed]
- Barrett, T.; Wilhite, S.E.; Ledoux, P.; Evangelista, C.; Kim, I.F.; Tomashevsky, M.; Marshall, K.A.; Phillippy, K.H.; Sherman, P.M.; Holko, M.; et al. NCBI GEO: Archive for functional genomics data sets—Update. Nucleic Acids Res. 2013, 41, D991–D995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brazma, A.; Parkinson, H.; Sarkans, U.; Shojatalab, M.; Vilo, J.; Abeygunawardena, N.; Holloway, E.; Kapushesky, M.; Kemmeren, P.; Lara, G.G.; et al. ArrayExpress—A public repository for microarray gene expression data at the EBI. Nucleic Acids Res. 2003, 31, 68–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, M.K.; Hagedorn, P.H.; de Torres-Zabala, M.; Grant, M.R.; Rung, J.H.; Collinge, D.B.; Lyngkjaer, M.F. Transcriptional regulation by an NAC (NAM-ATAF1,2-CUC2) transcription factor attenuates ABA signalling for efficient basal defence towards Blumeria graminis f. sp. hordei in Arabidopsis. Plant J. 2008, 56, 867–880. [Google Scholar]
- Ascencio-Ibáñez, J.T.; Sozzani, R.; Lee, T.J.; Chu, T.M.; Wolfinger, R.D.; Cella, R.; Hanley-Bowdoin, L. Global analysis of Arabidopsis gene expression uncovers a complex array of changes impacting pathogen response and cell cycle during geminivirus infection. Plant Physiol. 2008, 148, 436–454. [Google Scholar] [CrossRef] [Green Version]
- Mine, A.; Seyfferth, C.; Kracher, B.; Berens, M.L.; Becker, D.; Tsuda, K. The defense phytohormone signaling network enables rapid, high-amplitude transcriptional reprogramming during effector-triggered immunity. Plant Cell 2018, 30, 1199–1219. [Google Scholar] [CrossRef] [Green Version]
- Arabidopsis eFP Browser. Available online: http://bar.utoronto.ca/efp/cgi-bin/efpWeb.cgi (accessed on 17 March 2021).
- Zhang, W.; Corwin, J.A.; Copeland, D.; Feusier, J.; Eshbaugh, R.; Chen, F.; Atwell, S.; Kliebenstein, D.J. Plastic transcriptomes stabilize immunity to pathogen diversity: The jasmonic acid and salicylic acid networks within the Arabidopsis/Botrytis Pathosystem. Plant Cell 2017, 29, 2727–2752. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Corwin, J.A.; Copeland, D.H.; Feusier, J.; Eshbaugh, R.; Cook, D.E.; Atwell, S.; Kliebenstein, D.J. Plant-necrotroph co-transcriptome networks illuminate a metabolic battlefield. eLife 2019, 8, e44279. [Google Scholar] [CrossRef]
- Soltis, N.E.; Caseys, C.; Zhang, W.; Corwin, J.A.; Atwell, S.; Kliebenstein, D.J. Pathogen genetic control of transcriptome variation in the Arabidopsis thaliana-Botrytis cinerea pathosystem. Genetics 2020, 215, 253–266. [Google Scholar] [CrossRef] [Green Version]
- Bilgin, D.D.; Zavala, J.A.; Zhu, J.; Clough, S.J.; Ort, D.R.; DeLucia, E.H. Biotic stress globally downregulates photosynthesis genes. Plant Cell Environ. 2010, 33, 1597–1613. [Google Scholar] [CrossRef] [Green Version]
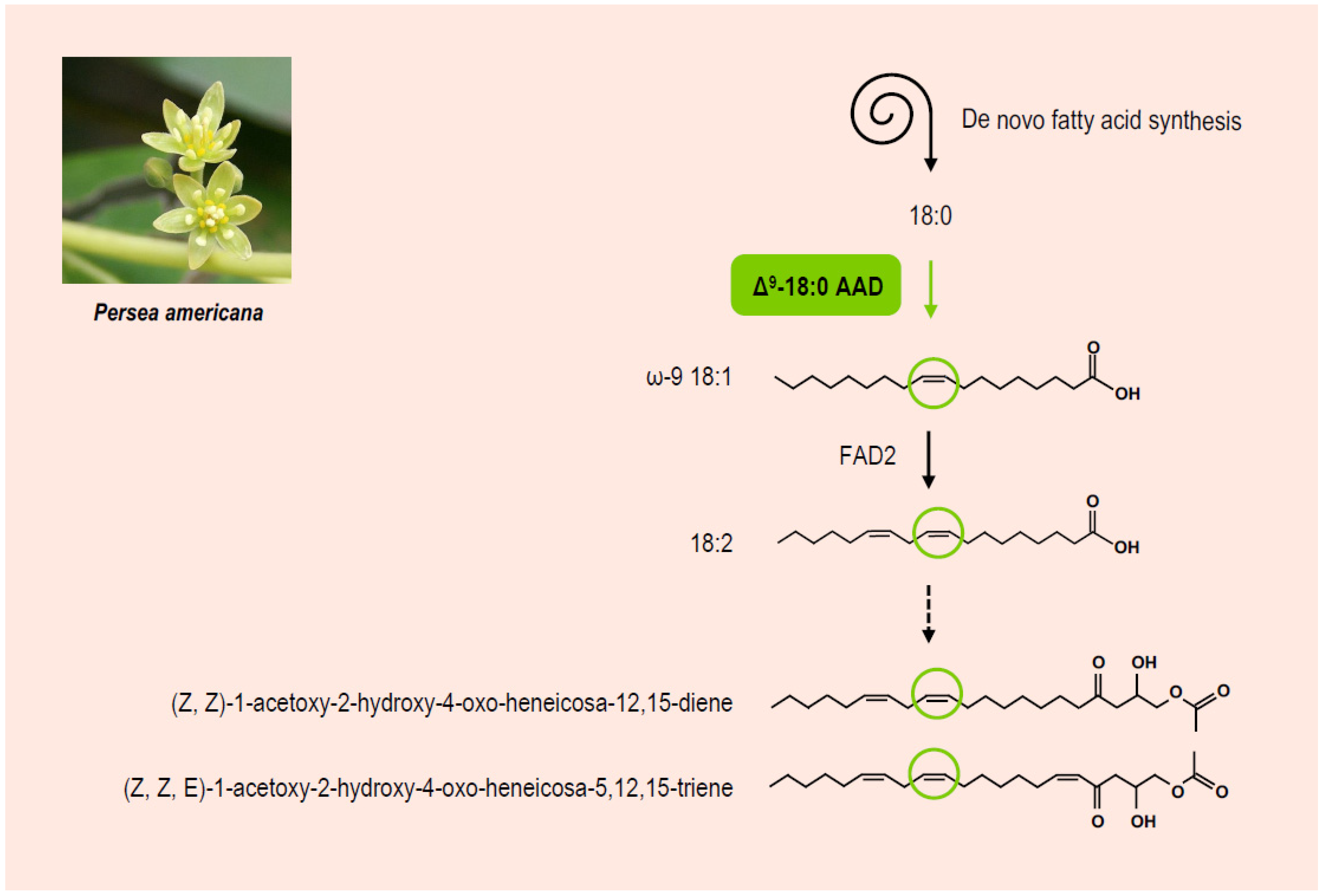
- Rodríguez-Sánchez, D.G.; Pacheco, A.; García-Cruz, M.I.; Gutiérrez-Uribe, J.A.; Benavides-Lozano, J.A.; Hernández-Brenes, C. Isolation and structure elucidation of avocado seed (Persea americana) lipid derivatives that inhibit Clostridium sporogenes endospore germination. J. Agric. Food Chem. 2013, 61, 7403–7411. [Google Scholar] [CrossRef]
- Yakoby, N.; Kobiler, I.; Dinoor, A.; Prusky, D. pH regulation of pectate lyase secretion modulates the attack of Collectotrichum gloeosporioides on avocado fruits. Appl. Environ. Microbiol. 2000, 66, 1026–1030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domergue, F.; Gregory, L.H.; Prusky, D.; Browse, J. Antifungal compounds from idioblast cells isolated from avocado fruits. Phytochemistry 2000, 54, 183–189. [Google Scholar] [CrossRef]
- Madi, L.; Wang, X.; Kobiler, I.; Lichter, A.; Prusky, D. Stress on avocado regulates ∆9-stearoyl ACP desaturase expression, fatty acid composition, antifungal diene level and resistance to Collectrichum gloeosporioides attack. Physiol. Mol. Plant Pathol. 2003, 62, 277–283. [Google Scholar] [CrossRef]
- Navez, B. Wikimedia Commons. File Licensed under the Creative Commons Attribution-Share 3.0 Unported License. Available online: https://commons.wikimedia.org/wiki/File:Persea_americana_flowers.jpg (accessed on 17 March 2021).
- Deeken, R.; Engelmann, J.C.; Efetova, M.; Czirjak, T.; Müller, T.; Kaiser, W.M.; Tietz, O.; Krischke, M.; Mueller, M.J.; Palme, K. An integrated view of gene expression and solute profiles of Arabidopsis tumors: A genome-wide approach. Plant Cell 2006, 18, 3617–3634. [Google Scholar] [CrossRef] [Green Version]
- Gigon, A.; Matos, A.-R.; Laffray, D.; Zuily-Fodil, Y.; Pham-Thi, A.-T. Effect of drought stress on lipid metabolism in the leaves of Arabidopsis thaliana (ecotype Columbia). Ann. Bot. 2004, 94, 345–351. [Google Scholar] [CrossRef]
- Shah, J.; Kachroo, P.; Nandi, A.; Klessig, D.F. A recessive mutation in the Arabidopsis SSI2 gene confers SA- and NPR1-independent expression of PR genes and resistance against bacterial and oomycete pathogens. Plant J. 2001, 25, 563–574. [Google Scholar] [CrossRef] [PubMed]
- Sekine, K.-T.; Nandi, A.; Ishihara, T.; Hase, S.; Ikegami, M.; Shah, J.; Takakashi, H. Enhanced resistance to Cucumber mosaic virus in the Arabidopsis thaliana ssi2 mutant is mediated via an SA-independent mechanism. Mol. Plant Microbe Interact. 2004, 17, 623–632. [Google Scholar] [CrossRef]
- Chandra-Shekara, A.C.; Venugopal, S.C.; Barman, S.R.; Kachroo, A.; Kachroo, P. Plastidial fatty acid levels regulate resistance gene-dependent defense signaling in Arabidopsis. Proc. Natl. Acad. Sci. USA 2007, 104, 7277–7282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, N.; Hu, Z.; Li, Y.; Li, C.; Peng, F.; Yao, Y.; Peng, H.; Ni, Z.; Xie, C.; Sun, Q. Overexpression of a wheat stearoyl-ACP desaturase (SACPD) gene TaSSI2 in Arabidopsis ssi2 mutant compromise its resistance to powdery mildew. Gene 2013, 524, 220–227. [Google Scholar] [CrossRef]
- Gao, Q.-M.; Venugopal, S.; Navarre, D.; Kachroo, A. Low oleic acid-derived repression of jasmonic acid-inducible defense responses requires the WRKY50 and WRKY51 proteins. Plant Physiol. 2011, 155, 464–476. [Google Scholar] [CrossRef] [Green Version]
- Kachroo, A.; Fu, D.-Q.; Havens, W.; Navarre, D.; Kachroo, P.; Ghabrial, S.A. An oleic acid-mediated pathway induces constitutive defense signaling and enhanced resistance to multiple pathogens in soybean. Mol. Plant Microbe Interact. 2008, 21, 564–575. [Google Scholar] [CrossRef] [Green Version]
- Jiang, C.-J.; Shimono, M.; Maeda, S.; Inoue, H.; Mori, M.; Hasegawa, M.; Sugano, S.; Takatsuji, H. Suppression of the rice fatty-acid desaturase gene OsSSI2 enhances resistance to blast and leaf blight diseases in rice. Mol. Plant Microbe Interact. 2009, 22, 820–829. [Google Scholar] [CrossRef] [Green Version]
- Kunst, L.; Browse, J.; Somerville, C. Altered regulation of lipid biosynthesis in a mutant of Arabidopsis deficient in chloroplast glycerol-3-phosphate acyltransferase activity. Proc. Natl. Acad. Sci. USA 1988, 85, 4113–4147. [Google Scholar] [CrossRef] [Green Version]
- Kachroo, A.; Lapchyk, L.; Fukushige, H.; Hildebrand, D.; Klessig, D.; Kachroo, P. Plastidial fatty acid signaling modulates salicylic acid- and jasmonic acid-mediated defense pathways in the Arabidopsis ssi2 mutant. Plant Cell 2003, 15, 2952–2965. [Google Scholar] [CrossRef] [Green Version]
- Xia, Y.; Gao, Q.-M.; Yu, K.; Lapchyk, L.; Navarre, D.; Hildebrand, D.; Kachroo, A.; Kachroo, P. An intact cuticle in distal tissues is essential for the induction of systemic acquired resistance in plants. Cell Host Microbe 2009, 5, 151–165. [Google Scholar] [CrossRef] [Green Version]
- Lorenc-Kukula, K.; Chaturvedi, R.; Roth, M.; Welti, R.; Shah, J. Biochemical and molecular-genetic characterization of SFD1’s involvement in lipid metabolism and defense signaling. Front. Plant Sci. 2012, 3, 26. [Google Scholar] [CrossRef] [Green Version]
- Kachroo, A.; Venugopal, S.C.; Lepchyk, L.; Falcone, D.; Hildebrand, D.; Kachroo, P. Oleci acid levels regulated by glycerolipid metabolism modulate defense gene expression in Arabidopsis. Proc. Natl. Acad. Sci. USA 2004, 101, 5152–5157. [Google Scholar] [CrossRef] [Green Version]
- Marie-Lan Nguyen. Wikimedia Commons. File Licensed under the Creative Commons Attribution 2.5 Generic License. Available online: https://commons.wikimedia.org/wiki/File:Arabidopsis_thaliana_JdP_2013-04-28.jpg (accessed on 17 March 2021).
- Mandal, M.K.; Chandra-Shekara, A.C.; Jeong, R.-D.; Yu, K.; Zhu, S.; Chanda, B.; Navarre, D.; Kachroo, A.; Kachroo, P. Oleic acid-dependent modulation of NITRIC OXIDE ASSOCIATED1 protein levels regulates nitric oxide-mediated defense signaling in Arabidopsis. Plant Cell 2012, 24, 1654–1674. [Google Scholar] [CrossRef] [Green Version]
- Davda, R.K.; Stepniakowski, K.T.; Lu, G.; Ullian, M.E.; Goodfriend, T.L.; Egan, B.M. Oleic acid inhibits endothelial nitric oxide synthase by a protein kinase C-independent mechanism. Hypertension 1995, 26, 764–770. [Google Scholar] [CrossRef]
- Venugopal, S.C.; Jeong, R.D.; Mandal, M.K.; Zhu, S.; Chandra-Shekara, A.C.; Xia, Y.; Hersh, M.; Stromberg, A.J.; Navarre, D.; Kachroo, A.; et al. Enhanced disease susceptibility 1 and salicylic acid act redundantly to regulate resistance gene-mediated signaling. PLoS Genet. 2009, 5, e1000545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kachroo, P.; Venugopal, S.C.; Navarre, D.A.; Lapchyk, L.; Kachroo, A. Role of salicylic acid and fatty acid desaturation pathways in ssi2-mediated signaling. Plant Physiol. 2005, 139, 1717–1735. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; El-Shetehy, M.; Shine, M.B.; Yu, K.; Navarre, D.; Wendehenne, D.; Kachroo, A.; Kachroo, P. Free radicals mediate systemic acquired resistance. Cell Rep. 2014, 7, 348–355. [Google Scholar] [CrossRef]
- Lim, G.H.; Shine, M.B.; de Lorenzo, L.; Yu, K.; Cui, W.; Navarre, D.; Hunt, A.G.; Lee, J.Y.; Kachroo, A.; Kachroo, P. Plasmodesmata localizing proteins regulate transport and signaling during systemic acquired immunity in plants. Cell Host Microbe 2016, 19, 541–549. [Google Scholar] [CrossRef] [Green Version]
- Osman, S.M.; Abdel-Megied, A.M.; Zain Eldain, M.H.; Haleema, S.; Gopinath, C.; Amma Sumalekshmy, S.; Aboul-Enein, H.Y. A highly sensitive GC-MS method for simultaneous determination of anacardic acids in cashew (Anacardium occidentale) nut shell oil in the presence of other phenolic lipid derivatives. Biomed. Chromatogr. 2019, 33, e4659. [Google Scholar] [CrossRef] [PubMed]
- Yalpani, M.; Tyman, J.H. The phenolics acids of Pistachia vera. Phytochemistry 1983, 22, 2263–2266. [Google Scholar] [CrossRef]
- Li, R.; Shen, Y.; Zhang, X.; Ma, M.; Chen, B.; van Beek, T.A. Efficient purification of ginkgolic acids from Ginkgo biloba leaves by selective adsorption on Fe3O4 magnetic nanoparticles. J. Nat. Prod. 2014, 77, 571–575. [Google Scholar] [CrossRef] [PubMed]
- Gerhold, D.L.; Craig, R.; Mumma, R.O. Analysis of trichome exudate from mite-resistant geraniums. J. Chem. Ecol. 1984, 10, 713–722. [Google Scholar] [CrossRef] [PubMed]
- Reffstrup, T.; Hammershoy, O.; Boll, P.M.; Schmidt, H. Philodendron scandens Koch Et Sello subsp. oxycardium (Schott) Bunting, a new source of allergenic alkyl resorcinols. Acta Chem. Scand. Ser. B 1982, 36, 291–294. [Google Scholar] [CrossRef]
- Chen, J.; Zhang, Y.H.; Wang, L.K.; Sucheck, S.J.; Snow, A.M.; Hecht, S.M. Inhibitors of DNA polymerase β from Schoepfia californica. Chem. Commun. 1998, 24, 2769–2770. [Google Scholar] [CrossRef]
- Lee, S.J.; Park, W.H.; Moon, H.-I. Bioassay-guided isolation of antiplasmodial anacardic acids derivatives from the whole plants of Viola websteri Hemsl. Parasitol. Res. 2009, 104, 463–466. [Google Scholar] [CrossRef]
- Spencer, G.F.; Tjarks, L.W.; Kleiman, R. Alkyl and phenylalkyl anacardic acids from Knema elegans seed oil. J. Nat. Prod. 1980, 43, 724–730. [Google Scholar] [CrossRef]
- Gény, C.; Rivière, G.; Bignon, J.; Birlirakis, N.; Guitter, E.; Awang, K.; Litaudon, M.; Roussi, F.; Dumontet, V. Anacradic acids from Knema kookeriana as modlulators of Bcl-xl/Bak and Mcl-1/bid interactions. J. Nat. Prod. 2016, 79, 838–844. [Google Scholar] [CrossRef]
- Lemes, L.F.N.; de Andrade Ramos, G.; de Oliveira, A.S.; da Silva, F.M.R.; de Castro Couto, G.; da Silva Boni, M.; Guimaräes, M.J.R.; Souza, I.N.P.; Bartolini, M.; Andrisano, V.; et al. Cardanol-derived AChE inhibitors: Towards the development of dual binding derivatives for Alzheimer’s disease. Eur. J. Med. Chem. 2016, 108, 687–700. [Google Scholar] [CrossRef]
- Huang, H.; Hua, X.; Liu, N.; Liu, S.; Chen, X.; Zhao, C.; Lan, X.; Yang, C.; Dou, Q.P.; Liu, J. Anacardic acid induces cell apoptosis associated with induction of ATF4-dependent endoplasmic reticulum stress. Toxicol. Lett. 2014, 228, 170–178. [Google Scholar] [CrossRef]
- Hemshekhar, M.; Santhosh, M.S.; Kemparaju, K.; Girish, K.S. Emerging roles of anacardic acid and its derivatives: A pharmacological overview. Basic Clin. Pharmacol. Toxicol. 2012, 110, 122–132. [Google Scholar] [CrossRef] [PubMed]
- Hollands, A.; Corriden, R.; Gysler, G.; Dahesh, S.; Olson, J.; Raza Ali, S.; Kunkel, M.T.; Lin, A.E.; Forli, S.; Newton, A.C.; et al. Natural product anacardic acid from cashew nut shells stimulates neutrophil extracellular trap production and bactericidal activity. J. Biol. Chem. 2016, 291, 13964–13973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prithiviraj, B.; Manickam, M.; Singh, U.P.; Ray, A.B. Antifungal activity of anacardic acid, a naturally occurring derivative of salicylic acid. Can. J. Bot. 1997, 75, 207–211. [Google Scholar] [CrossRef]
- Muzaffar, S.; Bose, S.; Banerji, A.; Nair, B.G.; Chattoo, B.B. Anacardic acid induces apoptosis-like cell death in the rice blast fungus Magnaporthe oryzae. Appl. Microbiol. Biotechnol. 2016, 100, 323–335. [Google Scholar] [CrossRef] [PubMed]
- Schultz, D.J.; Olsen, C.; Cobbs, G.A.; Stolowich, N.J.; Parrott, M.M. Bioactivity of anacardic acid against Colorado potato beetle (Leptinotarsa decemlineata) larvae. J. Agric. Food Chem. 2006, 54, 7522–7529. [Google Scholar] [CrossRef]
- Ferreira de Carvalho, G.H.; Lucília Dos Santos, M.; Monnerat, R.; Aparecida Andrade, M.; Gonçalves de Andrade, M.; Barbosa Dos Santos, A.; Marques Dourado Bastos, I.; de Santana, J.M. Ovicidal and deleterious effects of cashew (Anacardium occidentale) nut shell oil and its fractions on Musca domestica, Chrysoma megacephala, Anticarsia gemmatalis and Spodoptera frigiperda. Chem. Biodivers. 2019, 16, e1800468. [Google Scholar] [CrossRef]
- Matutino Bastos, T.; Mannochio Russo, H.; Silvio Moretti, N.; Schenkman, S.; Marcourt, L.; Gupta, M.P.; Wolfender, J.L.; Ferreira Queiroz, E.; Bothelho Pereira Soares, M. Chemical constituents of Anacardium occidentale as inhibitors of Trypanosoma cruzi sirtuins. Molecules 2019, 24, 1299. [Google Scholar] [CrossRef] [Green Version]
- Yuan, M.; Song, X.; Lv, W.; Xin, Q.; Wang, L.; Gao, Q.; Zhang, G.; Lioa, W.; Lian, S.; Jing, T. Effect of anacardic acid against echinococcosis through inhibition of VEGF-induced angiogenesis. Vet. Res. 2019, 50, 3. [Google Scholar] [CrossRef] [Green Version]
- Walters, D.S.; Craig, R.; Mumma, R.O. Fatty acid incorporation in the biosynthesis of anacardic acids of geraniums. Phytochemistry 1990, 29, 1815–1822. [Google Scholar] [CrossRef]
- Narnoliya, L.K.; Kaushal, G.; Singh, S.P.; Sangwan, R.S. De novo transcriptome analysis of rose-scented geranium provides insights into the metabolic specificity of terpene and tartaric acid biosynthesis. BMC Genom. 2017, 18, 74. [Google Scholar] [CrossRef] [Green Version]
- Schultz, D.J.; Wickramasinghe, N.S.; Klinge, C.M. Anacardic acid biosynthesis and bioactivity. Recent Adv. Phytochem. 2006, 40, 131–156. [Google Scholar]
- Oliveira, M.S.; Morais, S.M.; Magalhães, D.V.; Batista, W.P.; Vieira, I.G.; Craveiro, A.A.; de Manezes, J.E.; Carvalho, A.F.; de Lima, G.P. Antioxidant, larvicidal and antiacetylcholinesterase activities of cashew nut shell liquid constituents. Acta Trop. 2011, 117, 165–170. [Google Scholar] [CrossRef]
- Schultz, D.J.; Medford, J.I.; Cox-Foster, D.; Grazzini, R.A.; Craig, R.; Mumma, R.O. Anacardic acids in trichomes of Pelargonium: Biosynthesis, molecular biology and ecological effects. Adv. Bot. Res. 2000, 31, 175–192. [Google Scholar]
- Ha, T.J.; Kubo, I. Lipoxygenase inhibitory activity of anacardic acids. J. Agric. Food Chem. 2005, 53, 4350–4354. [Google Scholar] [CrossRef]
- Kubo, I.; Kinsthori, I.; Yokokawa, Y. Tyrosinase inhibitors from Anacardium occidentale fruits. J. Nat. Prod. 1994, 57, 545–551. [Google Scholar] [CrossRef]
- Sun, Y.; Jiang, X.; Chen, S.; Price, B.D. Inhibition of histone acetyltransferase activity by anacardic acid sensitizes tumor cells to ionizing radiation. FEBS Lett. 2006, 580, 4353–4356. [Google Scholar] [CrossRef] [Green Version]
- Forest and Kim Starr. Photo Gallery. File Licensed under the Creative Commons Attribution 3.0 Unported License. Available online: http://luirig.altervista.org/pics/display.php?pos=121940 (accessed on 17 March 2021).
- Schultz, D.J.; Cahoon, E.B.; Shanklin, J.; Craig, R.; Cox-Foster, D.L.; Mumma, R.O.; Meford, J.I. Expression of a Δ9 14:0-acyl carrier protein fatty acid desaturase gene is necessary for the production of ω5 anacardic acids found in pest-resisant geranium (Pelargonium xhortorum). Proc. Natl. Acad. Sci. USA 1996, 93, 8771–8775. [Google Scholar] [CrossRef] [Green Version]
- Grazzini, R.; Walters, D.; Harmon, J.; Hesk, D.J.; Cox-Foster, D.; Medford, J.; Craig, R.; Mumma, R.O. Inheritance of biochemical and morphological characters associated with two spotted spider mite resistance in Pelargonium x horturum. J. Am. Soc. Hortic. Sci. 1997, 122, 373–379. [Google Scholar] [CrossRef] [Green Version]
- Piñeiro Fernández, L.; Byers, K.J.R.P.; Cai, J.; Sedeek, K.E.M.; Kellenberger, R.T.; Russo, A.; Qi, W.; Aquino Fournier, C.; Schlüter, P.M. A phylogenomic analysis of the floral transcriptomes of sexually deceptive and rewarding European orchids, Ophrys and Gymnadenia. Front. Plant Sci. 2019, 29, 1553. [Google Scholar] [CrossRef] [PubMed]
- Breitkopf, H.; Onstein, R.E.; Cafasso, D.; Schülter, P.M.; Cozzolino, S. Multiple shifts to different pollinators fuelled rapid diversification in sexually deceptive Ophrys orchids. New Phytol. 2015, 207, 377–389. [Google Scholar] [CrossRef] [PubMed]
- Schlüter, P.M.; Schiestl, F.P. Molecular mechanisms of floral mimicry in orchids. Trends Plant Sci. 2008, 13, 228–235. [Google Scholar] [CrossRef] [PubMed]
- Bohman, B.; Weinstein, A.M.; Mozuraitis, R.; Flematti, G.; Borg-Karlson, A.-K. Identification of (Z)-8-heptadecene and n-pentadecane as electrophysiologically active compounds in Ophrys insectifera and its Argorytes pollinator. Int. J. Mol. Sci. 2020, 17, 21. [Google Scholar]
- Galizia, C.G.; Kunze, J.; Gumbert, A.; Borg-Karlson, A.-K.; Sachse, S.; Markl, C.; Menzel, R. Relationship of visual and olfactory signal parameters in food-deceptive flower mimicry system. Behav. Ecol. 2005, 16, 159–168. [Google Scholar] [CrossRef]
- Gasket, A.C. Orchid pollination by sexual deception: Pollinator perspectives. Biol. Rev. 2011, 86, 33–75. [Google Scholar] [CrossRef] [PubMed]
- Bohman, B.; Flematti, G.R.; Barrow, R.A.; Pichersky, E.; Peakall, R. Pollination by sexual deception-it takes chemistry to work. Curr. Opin. Plant Biol. 2016, 32, 37–46. [Google Scholar] [CrossRef]
- Xu, S.; Schlüter, P.M. Modeling the two-locus architecture of divergent pollinator adaptation: How variation in SAD paralogs affects fitness and evolutionary divergence in sexually deceptive orchids. Ecol. Evol. 2015, 5, 493–502. [Google Scholar] [CrossRef]
- Xu, S.; Schlüter, P.M.; Grossniklaus, U.; Schiestl, F.P. The genetic basis of pollinator adaptation in a sexually deceptive orchid. PLoS Genet. 2012, 8, e1002889. [Google Scholar] [CrossRef]
- Bernard, A.; Domergue, F.; Pascal, S.; Jetter, R.; Renne, C.; Faure, J.-D.; Haslam, R.P.; Napier, J.A.; Lessire, R.; Joubès, J. Reconstitution of plant alkane biosynthesis in yeast demonstrates the Arabidopsis ECERIFERUM1 and ECERIFERUM3 are core components of a very-long-chain alkane synthesis complex. Plant Cell 2012, 24, 3106–3118. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Suh, M. Advances in the understanding of cuticular waxes in Arabidopsis thaliana and crop species. Plant Cell Rep. 2015, 34, 557–572. [Google Scholar] [CrossRef]
- Schlüter, P.M.; Xu, S.; Gagliardini, V.; Whitlle, E.; Shanklin, J.; Grossniklaus, U.; Schiestl, F.P. Stearoyl-acyl carrier protein desaturases are associated with floral isolation in sexually deceptive orchids. Proc. Natl. Acad. Sci. USA 2011, 108, 5696–5701. [Google Scholar] [CrossRef] [Green Version]
- Espirat. Wikimedia Commons. File Licensed under the Creative Commons Attribution-Share 4.0 International License. Available online: https://commons.wikimedia.org/wiki/File:OPHRYS_SPHEGODES.jpg (accessed on 17 March 2021).
- Esculapio. Wikimedia Commons. File Licensed under the Creative Commons Attribution 2.5 Generic License. Available online: https://commons.wikimedia.org/wiki/File:Ophrys_exaltata_zingaro_112.jpg (accessed on 17 March 2021).
- Sedeek, K.E.M.; Whittle, E.; Guthörl, D.; Grossniklaus, U.; Shanklin, J.; Schlüter, P.M. Amino acid change in an orchid desaturase enables mimicry of the pollinator’s sex pheromone. Curr. Biol. 2016, 26, 1505–1511. [Google Scholar] [CrossRef] [Green Version]
- Sedeek, K.E.M.; Qi, W.; Schauer, M.A.; Gupta, A.K.; Poveda, L.; Xu, S.; Liu, Z.-J.; Grossniklaus, U.; Schiestl, F.P.; Schlüter, P.M. Transcriptome and proteome data reveal candidate genes for pollinator attraction in sexually deceptive orchids. PLoS ONE 2013, 8, e64621. [Google Scholar] [CrossRef]
- Monteiro, F.; Sebastiana, M.; Figueiredo, A.; Sousa, L.; Cotrim, H.C.; Pais, M.S. Labellum transcriptome reveals alkene biosynthetic genes involved in orchid sexual deception and pollination-induced senescence. Funct. Integr. Genom. 2012, 12, 693–703. [Google Scholar] [CrossRef]
- Vereecken, N.J.; Wilson, C.A.; Hötling, S.; Schulz, S.; Banketov, S.A.; Mardulyn, P. Pre-adaptations and the evolution of pollination by sexual deception: Cope’s rule of specialization revisited. Proc. Biol. Sci. 2012, 279, 4786–4794. [Google Scholar] [CrossRef] [Green Version]
- Ellis, A.G.; Johnson, S.D. Floral mimicry enhances pollen export: The evolution of pollination by sexual deceit outside the Orchidaceae. Am. Nat. 2010, 176, E143–E151. [Google Scholar] [CrossRef]
- Fuji, T.; Yamamoto, M.; Nakano, R.; Nirazawa, T.; Rong, Y.; Dong, S.L.; Ishikawa, Y. Alkenyl sex pheromone analogs in the hemolymph of an arctiid Eilema japonica and several non-arctiid moths. J. Insect Physiol. 2015, 82, 109–113. [Google Scholar] [CrossRef]
- Silva, W.D.; Bento, J.M.S.; Hanks, L.M.; Millar, J.G. (Z)-7-Hexadecene is an aggregation-sex pheromone produced by males of the South American cerambycid beetle Susuacanga octoguttata. J. Chem. Ecol. 2018, 44, 1115–1119. [Google Scholar] [CrossRef] [PubMed]
- Van Vang, L.; Yan, Q.; Nghia, N.T.N.; Khank, C.N.Q.; Ando, T. Unsaturated cuticular hydrocarbon components of the sex pheromone of eggplant fruit borer, Leucinodes orbonalis Guenée (Lepidoptera: Crambidae). J. Chem. Ecol. 2018, 44, 631–636. [Google Scholar] [CrossRef] [PubMed]
- Jeon, J.E.; Kim, J.G.; Fischer, C.R.; Mehta, N.; Dufour-Schroif, C.; Wemmer, K.; Mudgett, M.B.; Sattely, E. A pathogen-responsive gene cluster for highly modified fatty acids in tomato. Cell 2020, 180, 176–187. [Google Scholar] [CrossRef]
- Zidorn, C.; Jöhrer, K.; Ganzera, M.; Schubert, B.; Sigmund, E.M.; Mader, J.; Greil, R.; Ellmerer, E.P.; Stuppner, H. Polyacetylenes from the Apiaceae vegetables carrot, celery, fennel, parsley, and parsnip and their cytotoxic activities. J. Agric. Food Chem. 2005, 53, 2518–2523. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kazaz, S.; Miray, R.; Baud, S. Acyl–Acyl Carrier Protein Desaturases and Plant Biotic Interactions. Cells 2021, 10, 674. https://doi.org/10.3390/cells10030674
Kazaz S, Miray R, Baud S. Acyl–Acyl Carrier Protein Desaturases and Plant Biotic Interactions. Cells. 2021; 10(3):674. https://doi.org/10.3390/cells10030674
Chicago/Turabian StyleKazaz, Sami, Romane Miray, and Sébastien Baud. 2021. "Acyl–Acyl Carrier Protein Desaturases and Plant Biotic Interactions" Cells 10, no. 3: 674. https://doi.org/10.3390/cells10030674
APA StyleKazaz, S., Miray, R., & Baud, S. (2021). Acyl–Acyl Carrier Protein Desaturases and Plant Biotic Interactions. Cells, 10(3), 674. https://doi.org/10.3390/cells10030674