
Reticulated Platelets—Which Functions Have Been Established by In Vivo and In Vitro Data?

 ,
,  ,
,
Abstract
:1. Introduction
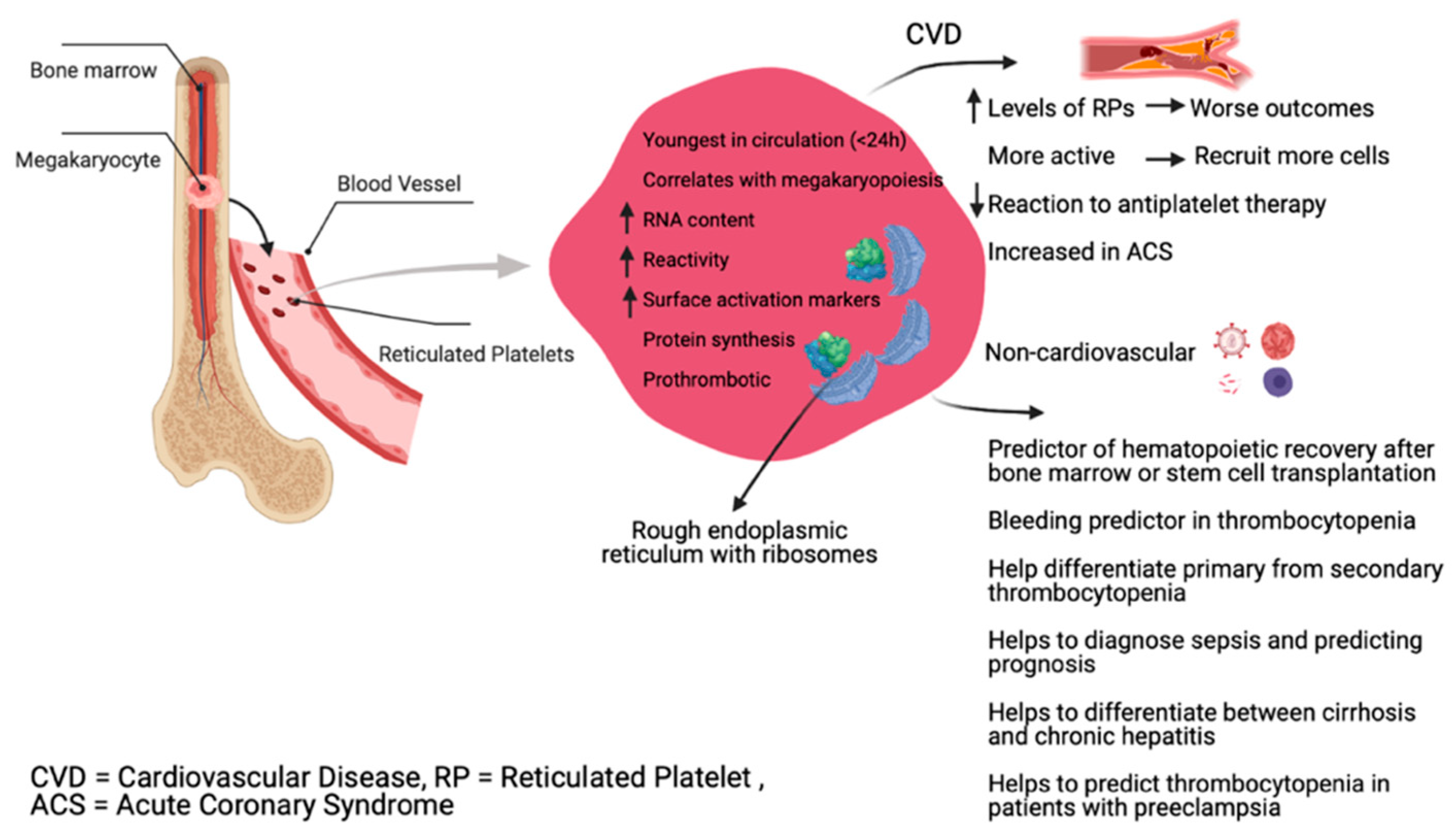
2. Physiology of Reticulated Platelets
3. Pre-Analytical Aspects
Pathogens
4. Methods for Reticulated Platelets Determination
4.1. Flow Cytometry
4.2. Fully Automated Analyzers
4.2.1. Sysmex Analyzers
4.2.2. Abbott Analyzer
4.2.3. Mindray Analyzer
5. In Vivo Data
5.1. Animal Studies
RP in Diabetes and Increased CVD Risk
5.2. Clinical Utility of RP
5.2.1. Thrombocytopenia
5.2.2. Myelodysplastic Syndromes
5.2.3. Cardiovascular Disease and Antiplatelet Therapy
5.2.4. Infection Diagnosis and Control
5.2.5. Pregnancy Complications
5.2.6. Liver Diseases
6. In Vitro Data
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Clark, V.L.; Kruse, J.A. Clinical Methods: The History, Physical, and Laboratory Examinations, 3rd ed.; Walker, H.K., Hall, W.D., Hurst, J.W., Eds.; Butterworths: Boston, MA, USA, 1990; ISBN 978-0-409-90077-4. [Google Scholar]
- Muronoi, T.; Koyama, K.; Nunomiya, S.; Lefor, A.K.; Wada, M.; Koinuma, T.; Shima, J.; Suzukawa, M. Immature platelet fraction predicts coagulopathy-related platelet consumption and mortality in patients with sepsis. Thromb. Res. 2016, 144, 169–175. [Google Scholar] [CrossRef]
- Wong, C.H.Y.; Jenne, C.N.; Petri, B.; Chrobok, N.L.; Kubes, P. Nucleation of platelets with blood-borne pathogens on Kupffer cells precedes other innate immunity and contributes to bacterial clearance. Nat. Immunol. 2013, 14, 785–792. [Google Scholar] [CrossRef] [Green Version]
- Barsam, S.J.; Psaila, B.; Forestier, M.; Page, L.K.; Sloane, P.A.; Geyer, J.T.; Villarica, G.O.; Ruisi, M.M.; Gernsheimer, T.B.; Beer, J.H.; et al. Platelet production and platelet destruction: Assessing mechanisms of treatment effect in immune thrombocytopenia. Blood 2011, 117, 5723–5732. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, J.J.M.L. Reticulated platelets: Analytical aspects and clinical utility. Clin. Chem. Lab. Med. 2014, 52, 1107–1117. [Google Scholar] [CrossRef] [Green Version]
- Ts’ao, C.H. Rough endoplasmic reticulum and ribosomes in blood platelets. Scand. J. Haematol. 1971, 8, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Angénieux, C.; Maître, B.; Eckly, A.; Lanza, F.; Gachet, C.; de la Salle, H. Time-Dependent Decay of mRNA and Ribosomal RNA during Platelet Aging and Its Correlation with Translation Activity. PLoS ONE 2016, 11, e0148064. [Google Scholar] [CrossRef]
- Schmitt, A.; Guichard, J.; Massé, J.M.; Debili, N.; Cramer, E.M. Of mice and men: Comparison of the ultrastructure of megakaryocytes and platelets. Exp. Hematol. 2001, 29, 1295–1302. [Google Scholar] [CrossRef]
- Jackson, C.W.; Steward, S.A.; Chenaille, P.J.; Ashmun, R.A.; McDonald, T.P. An analysis of megakaryocytopoiesis in the C3H mouse: An animal model whose megakaryocytes have 32N as the modal DNA class. Blood 1990, 76, 690–696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levin, J.; Ebbe, S. Why are recently published platelet counts in normal mice so low? Blood 1994, 83, 3829–3831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukuda, T.; Asou, E.; Nogi, K.; Goto, K. Evaluation of mouse red blood cell and platelet counting with an automated hematology analyzer. J. Vet. Med. Sci. 2017, 79, 1707–1711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corash, L.; Chen, H.Y.; Levin, J.; Baker, G.; Lu, H.; Mok, Y. Regulation of thrombopoiesis: Effects of the degree of thrombocytopenia on megakaryocyte ploidy and platelet volume. Blood 1987, 70, 177–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasmussen, T.V.; Ahlen, M.T. Murine models for studying treatment, prevention and pathogenesis of FNAIT. Transfus. Apher. Sci. 2020, 59, 102706. [Google Scholar] [CrossRef]
- Ault, K.A.; Knowles, C. In Vivo biotinylation demonstrates that reticulated platelets are the youngest platelets in circulation. Exp. Hematol. 1995, 23, 996–1001. [Google Scholar] [PubMed]
- Ault, K.A.; Rinder, H.M.; Mitchell, J.; Carmody, M.B.; Vary, C.P.; Hillman, R.S. The significance of platelets with increased RNA content (reticulated platelets). A measure of the rate of thrombopoiesis. Am. J. Clin. Pathol. 1992, 98, 637–646. [Google Scholar] [CrossRef] [PubMed]
- Stohlawetz, P.; Schulenburg, A.; Stiegler, G.; Panzer, S.; Höcker, P.; Kalhs, P.; Schwarzinger, I.; Müllauer, L.; Simonitsch, I.; Geissler, K.; et al. The proportion of reticulated platelets is higher in bone marrow than in peripheral blood in haematological patients. Eur. J. Haematol. 1999, 63, 239–244. [Google Scholar] [CrossRef]
- Pankraz, A.; Ledieu, D.; Pralet, D.; Provencher-Bolliger, A. Detection of reticulated platelets in whole blood of rats using flow cytometry. Exp. Toxicol. Pathol. Off. J. Ges. Toxikol. Pathol. 2008, 60, 443–448. [Google Scholar] [CrossRef] [PubMed]
- Kono, M.; Matsuhiroya, S.; Nakazawa, F.; Kaido, M.; Wada, A.; Tomiyama, Y. Morphological and optical properties of human immature platelet-enriched population produced in immunodeficient mice. Platelets 2019, 30, 652–657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kern, B.; Molineux, G.; Briddell, R. A method for the determination of the number of reticulated platelets from whole blood. Exp. Hematol. 2000, 28, 92. [Google Scholar] [CrossRef]
- Detwiler, T.C.; Odell, T.T.; MacDONALD, T.P. Platelet size, ATP content, and clot retraction in relation to platelet age. Am. J. Physiol. 1962, 203, 107–110. [Google Scholar] [CrossRef] [PubMed]
- Martin, J.F.; Trowbridge, E.A.; Salmon, G.; Plumb, J. The biological significance of platelet volume: Its relationship to bleeding time, platelet thromboxane B2 production and megakaryocyte nuclear DNA concentration. Thromb. Res. 1983, 32, 443–460. [Google Scholar] [CrossRef]
- Balduini, C.L.; Noris, P.; Spedini, P.; Belletti, S.; Zambelli, A.; Da Prada, G.A. Relationship between size and thiazole orange fluorescence of platelets in patients undergoing high-dose chemotherapy. Br. J. Haematol. 1999, 106, 202–207. [Google Scholar] [CrossRef] [PubMed]
- Mazzi, S.; Lordier, L.; Debili, N.; Raslova, H.; Vainchenker, W. Megakaryocyte and polyploidization. Exp. Hematol. 2018, 57, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Bessman, J.D. The relation of megakaryocyte ploidy to platelet volume. Am. J. Hematol. 1984, 16, 161–170. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, J.J.M.L.; van den Broek, N.M.A.; Curvers, J. Reference intervals of reticulated platelets and other platelet parameters and their associations. Arch. Pathol. Lab. Med. 2013, 137, 1635–1640. [Google Scholar] [CrossRef] [PubMed]
- Handtke, S.; Thiele, T. Large and small platelets-(When) do they differ? J. Thromb. Haemost. JTH 2020, 18, 1256–1267. [Google Scholar] [CrossRef] [PubMed]
- Golde, W.T.; Gollobin, P.; Rodriguez, L.L. A rapid, simple, and humane method for submandibular bleeding of mice using a lancet. Lab Anim. 2005, 34, 39–43. [Google Scholar] [CrossRef] [PubMed]
- Parasuraman, S.; Raveendran, R.; Kesavan, R. Blood sample collection in small laboratory animals. J. Pharmacol. Pharmacother. 2010, 1, 87–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishiyama, M.; Hayashi, S.; Kabutomori, O.; Yamanishi, H.; Suehisa, E.; Kurata, Y.; Kashiwagi, H.; Tomiyama, Y. Effects of anticoagulants and storage temperature on immature platelet fraction % (IPF%) values in stored samples measured by the automated hematology analyzer, XE-5000--utility of CTAD-anticoagulation and room temperature storage. Rinsho Byori. Jpn. J. Clin. Pathol. 2011, 59, 452–458. [Google Scholar]
- Meintker, L.; Haimerl, M.; Ringwald, J.; Krause, S.W. Measurement of immature platelets with Abbott CD-Sapphire and Sysmex XE-5000 in haematology and oncology patients. Clin. Chem. Lab. Med. 2013, 51, 2125–2131. [Google Scholar] [CrossRef]
- Buttarello, M.; Mezzapelle, G.; Freguglia, F.; Plebani, M. Reticulated platelets and immature platelet fraction: Clinical applications and method limitations. Int. J. Lab. Hematol. 2020, 42, 363–370. [Google Scholar] [CrossRef] [Green Version]
- Etulain, J.; Mena, H.A.; Meiss, R.P.; Frechtel, G.; Gutt, S.; Negrotto, S.; Schattner, M. An optimised protocol for platelet-rich plasma preparation to improve its angiogenic and regenerative properties. Sci. Rep. 2018, 8, 1513. [Google Scholar] [CrossRef]
- Matos-Rodrigues, G.E.; Masseron, C.C.; Silva, F.J.M.D.; Frajblat, M.; Moreira, L.O.; Martins, R.A.P. PCR-based detection of Helicobacter spp. in animal facilities of a University in Rio de Janeiro, Brazil. An. Acad. Bras. Cienc. 2020, 92, e20191517. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.; MacHin, S.; Mackie, I.; Harrison, P. In vivo biotinylation studies: Specificity of labelling of reticulated platelets by thiazole orange and mepacrine. Br. J. Haematol. 2000, 108, 859–864. [Google Scholar] [CrossRef] [PubMed]
- Hille, L.; Cederqvist, M.; Hromek, J.; Stratz, C.; Trenk, D.; Nührenberg, T.G. Evaluation of an Alternative Staining Method Using SYTO 13 to Determine Reticulated Platelets. Thromb. Haemost. 2019, 119, 779–785. [Google Scholar] [CrossRef] [Green Version]
- Wada, A.; Takagi, Y.; Kono, M.; Morikawa, T. Accuracy of a New Platelet Count System (PLT-F) Depends on the Staining Property of Its Reagents. PLoS ONE 2015, 10, e0141311. [Google Scholar] [CrossRef] [PubMed]
- Jung, H.; Jeon, H.-K.; Kim, H.-J.; Kim, S.-H. Immature platelet fraction: Establishment of a reference interval and diagnostic measure for thrombocytopenia. Korean J. Lab. Med. 2010, 30, 451–459. [Google Scholar] [CrossRef]
- Ali, U.; Knight, G.; Gibbs, R.; Tsitsikas, D.A. Reference intervals for absolute and percentage immature platelet fraction using the Sysmex XN-10 automated haematology analyser in a UK population. Scand. J. Clin. Lab. Investig. 2017, 77, 658–664. [Google Scholar] [CrossRef] [Green Version]
- Morkis, I.V.C.; Farias, M.G.; Scotti, L. Determination of reference ranges for immature platelet and reticulocyte fractions and reticulocyte hemoglobin equivalent. Rev. Bras. Hematol. E Hemoter. 2016, 38, 310–313. [Google Scholar] [CrossRef] [Green Version]
- Haffner, S.M.; Lehto, S.; Rönnemaa, T.; Pyörälä, K.; Laakso, M. Mortality from coronary heart disease in subjects with type 2 diabetes and in nondiabetic subjects with and without prior myocardial infarction. N. Engl. J. Med. 1998, 339, 229–234. [Google Scholar] [CrossRef]
- Lüscher, T.F.; Creager, M.A.; Beckman, J.A.; Cosentino, F. Diabetes and vascular disease: Pathophysiology, clinical consequences, and medical therapy: Part II. Circulation 2003, 108, 1655–1661. [Google Scholar] [CrossRef] [PubMed]
- Pasterkamp, G. Methods of accelerated atherosclerosis in diabetic patients. Heart Br. Card. Soc. 2013, 99, 743–749. [Google Scholar] [CrossRef]
- Kraakman, M.J.; Lee, M.K.S.; Al-Sharea, A.; Dragoljevic, D.; Barrett, T.J.; Montenont, E.; Basu, D.; Heywood, S.; Kammoun, H.L.; Flynn, M.; et al. Neutrophil-derived S100 calcium-binding proteins A8/A9 promote reticulated thrombocytosis and atherogenesis in diabetes. J. Clin. Investig. 2017, 127, 2133–2147. [Google Scholar] [CrossRef] [Green Version]
- Sakuragi, M.; Hayashi, S.; Maruyama, M.; Kabutomori, O.; Kiyokawa, T.; Nagamine, K.; Kato, H.; Kashiwagi, H.; Kanakura, Y.; Tomiyama, Y. Clinical significance of IPF% or RP% measurement in distinguishing primary immune thrombocytopenia from aplastic thrombocytopenic disorders. Int. J. Hematol. 2015, 101, 369–375. [Google Scholar] [CrossRef] [PubMed]
- Pons, I.; Monteagudo, M.; Lucchetti, G.; Muñoz, L.; Perea, G.; Colomina, I.; Guiu, J.; Obiols, J. Correlation between immature platelet fraction and reticulated platelets. Usefulness in the etiology diagnosis of thrombocytopenia. Eur. J. Haematol. 2010, 85, 158–163. [Google Scholar] [PubMed]
- Kienast, J.; Schmitz, G. Flow cytometric analysis of thiazole orange uptake by platelets: A diagnostic aid in the evaluation of thrombocytopenic disorders. Blood 1990, 75, 116–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Briggs, C.; Kunka, S.; Hart, D.; Oguni, S.; Machin, S.J. Assessment of an immature platelet fraction (IPF) in peripheral thrombocytopenia. Br. J. Haematol. 2004, 126, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Cybulska, A.; Meintker, L.; Ringwald, J.; Krause, S.W. Measurements of immature platelets with haematology analysers are of limited value to separate immune thrombocytopenia from bone marrow failure. Br. J. Haematol. 2017, 177, 612–619. [Google Scholar] [CrossRef] [Green Version]
- Larruzea Ibarra, A.; Muñoz Marín, L.; Perea Durán, G.; Torra Puig, M. Evaluation of immature platelet fraction in patients with myelodysplastic syndromes. Association with poor prognosis factors. Clin. Chem. Lab. Med. 2019, 57, e128–e130. [Google Scholar] [CrossRef]
- Sugimori, N.; Kondo, Y.; Shibayama, M.; Omote, M.; Takami, A.; Sugimori, C.; Ishiyama, K.; Yamazaki, H.; Nakao, S. Aberrant increase in the immature platelet fraction in patients with myelodysplastic syndrome: A marker of karyotypic abnormalities associated with poor prognosis. Eur. J. Haematol. 2009, 82, 54–60. [Google Scholar] [CrossRef]
- Cesari, F.; Marcucci, R.; Gori, A.M.; Caporale, R.; Fanelli, A.; Casola, G.; Balzi, D.; Barchielli, A.; Valente, S.; Giglioli, C.; et al. Reticulated platelets predict cardiovascular death in acute coronary syndrome patients. Insights from the AMI-Florence 2 Study. Thromb. Haemost. 2013, 109, 846–853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stratz, C.; Nührenberg, T.; Valina, C.; Löffelhardt, N.; Mashayekhi, K.; Ferenc, M.; Trenk, D.; Neumann, F.-J.; Hochholzer, W. Impact of Reticulated Platelets on the Antiplatelet Effect of the Intravenous P2Y12-Receptor Inhibitor Cangrelor. Thromb. Haemost. 2018, 118, 362–368. [Google Scholar] [CrossRef]
- McBane, R.D.; Gonzalez, C.; Hodge, D.O.; Wysokinski, W.E. Propensity for young reticulated platelet recruitment into arterial thrombi. J. Thromb. Thrombolysis 2014, 37, 148–154. [Google Scholar] [CrossRef]
- Armstrong, P.C.; Hoefer, T.; Knowles, R.B.; Tucker, A.T.; Hayman, M.A.; Ferreira, P.M.; Chan, M.V.; Warner, T.D. Newly Formed Reticulated Platelets Undermine Pharmacokinetically Short-Lived Antiplatelet Therapies. Arterioscler. Thromb. Vasc. Biol. 2017, 37, 949–956. [Google Scholar] [CrossRef]
- Liu, Q.-H.; Song, M.-Y.; Yang, B.-X.; Xia, R.-X. Clinical significance of measuring reticulated platelets in infectious diseases. Medicine 2017, 96, e9424. [Google Scholar] [CrossRef]
- De Blasi, R.A.; Cardelli, P.; Costante, A.; Sandri, M.; Mercieri, M.; Arcioni, R. Immature platelet fraction in predicting sepsis in critically ill patients. Intensive Care Med. 2013, 39, 636–643. [Google Scholar] [CrossRef]
- Koyama, K.; Katayama, S.; Muronoi, T.; Tonai, K.; Goto, Y.; Koinuma, T.; Shima, J.; Nunomiya, S. Time course of immature platelet count and its relation to thrombocytopenia and mortality in patients with sepsis. PLoS ONE 2018, 13, e0192064. [Google Scholar] [CrossRef] [Green Version]
- Everett, T.R.; Garner, S.F.; Lees, C.C.; Goodall, A.H. Immature platelet fraction analysis demonstrates a difference in thrombopoiesis between normotensive and preeclamptic pregnancies. Thromb. Haemost. 2014, 111, 1177–1179. [Google Scholar] [CrossRef] [PubMed]
- Nomura, T.; Kubota, Y.; Kitanaka, A.; Kurokouchi, K.; Inage, T.; Saigo, K.; Iseki, K.; Baba, N.; Yamaoka, G.; Arai, T.; et al. Immature platelet fraction measurement in patients with chronic liver disease: A convenient marker for evaluating cirrhotic change. Int. J. Lab. Hematol. 2010, 32, 299–306. [Google Scholar] [CrossRef]
- Kim, H.R.; Park, B.R.G.; Lee, M.K.; Park, A.J.; Ahn, J.Y. Comparison of an immature platelet fraction and reticulated platelet in liver cirrhosis. Korean J. Lab. Med. 2007, 27, 7–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rauber, P.; Lammert, F.; Grotemeyer, K.; Appenrodt, B. Immature platelet fraction and thrombopoietin in patients with liver cirrhosis: A cohort study. PLoS ONE 2018, 13, e0192271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonçalo, A.P.; Barbosa, I.L.; Campilho, F.; Campos, A.; Mendes, C. Predictive value of immature reticulocyte and platelet fractions in hematopoietic recovery of allograft patients. Transplant. Proc. 2011, 43, 241–243. [Google Scholar] [CrossRef]
- Takami, A.; Shibayama, M.; Orito, M.; Omote, M.; Okumura, H.; Yamashita, T.; Shimadoi, S.; Yoshida, T.; Nakao, S.; Asakura, H. Immature platelet fraction for prediction of platelet engraftment after allogeneic stem cell transplantation. Bone Marrow Transplant. 2007, 39, 501–507. [Google Scholar] [CrossRef]
- Saigo, K.; Sakota, Y.; Masuda, Y.; Matsunaga, K.; Takenokuchi, M.; Nishimura, K.; Sugimoto, T.; Sakurai, K.; Hashimoto, M.; Yanai, T.; et al. Automatic detection of immature platelets for decision making regarding platelet transfusion indications for pediatric patients. Transfus. Apher. Sci. Off. J. World Apher. Assoc. Off. J. Eur. Soc. Haemapheresis 2008, 38, 127–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Have, L.W.J.; Hasle, H.; Vestergaard, E.M.; Kjaersgaard, M. Absolute immature platelet count may predict imminent platelet recovery in thrombocytopenic children following chemotherapy. Pediatr. Blood Cancer 2013, 60, 1198–1203. [Google Scholar] [CrossRef] [PubMed]
- Morkis, I.V.C.; Farias, M.G.; Rigoni, L.D.C.; Scotti, L.; Gregianin, L.J.; Daudt, L.E.; Silla, L.M.d.R.; Paz, A.A. Assessment of immature platelet fraction and immature reticulocyte fraction as predictors of engraftment after hematopoietic stem cell transplantation. Int. J. Lab. Hematol. 2015, 37, 259–264. [Google Scholar] [CrossRef]
- Ibrahim, H.; Nadipalli, S.; Usmani, S.; DeLao, T.; Green, L.; Kleiman, N.S. Detection and quantification of circulating immature platelets: Agreement between flow cytometric and automated detection. J. Thromb. Thrombolysis 2016, 42, 77–83. [Google Scholar] [CrossRef]
- Gonzalez-Porras, J.R.; Martin-Herrero, F.; Gonzalez-Lopez, T.J.; Olazabal, J.; Diez-Campelo, M.; Pabon, P.; Alberca, I.; San Miguel, J.F. The role of immature platelet fraction in acute coronary syndrome. Thromb. Haemost. 2010, 103, 247–249. [Google Scholar] [CrossRef] [PubMed]
- López-Jiménez, R.A.; Martín-Herrero, F.; González-Porras, J.R.; Sánchez-Barba, M.; Martín-Luengo, C.; Pabón-Osuna, P. Immature Platelet Fraction: A New Prognostic Marker in Acute Coronary Syndrome. Rev. Esp. Cardiol. Engl. Ed. 2013, 66, 147–148. [Google Scholar] [CrossRef] [PubMed]
- Grove, E.L.; Hvas, A.-M.; Kristensen, S.D. Immature platelets in patients with acute coronary syndromes. Thromb. Haemost. 2009, 101, 151–156. [Google Scholar]
- Tscharre, M.; Farhan, S.; Bruno, V.; Rohla, M.; Egger, F.; Weiss, T.W.; Hübl, W.; Willheim, M.; Wojta, J.; Geppert, A.; et al. Impact of platelet turnover on long-term adverse cardiovascular outcomes in patients undergoing percutaneous coronary intervention. Eur. J. Clin. Investig. 2019, 49, e13157. [Google Scholar] [CrossRef]
- Ibrahim, H.; Schutt, R.C.; Hannawi, B.; DeLao, T.; Barker, C.M.; Kleiman, N.S. Association of immature platelets with adverse cardiovascular outcomes. J. Am. Coll. Cardiol. 2014, 64, 2122–2129. [Google Scholar] [CrossRef] [Green Version]
- Lakkis, N.; Dokainish, H.; Abuzahra, M.; Tsyboulev, V.; Jorgensen, J.; De Leon, A.P.; Saleem, A. Reticulated platelets in acute coronary syndrome: A marker of platelet activity. J. Am. Coll. Cardiol. 2004, 44, 2091–2093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stratz, C.; Bömicke, T.; Younas, I.; Kittel, A.; Amann, M.; Valina, C.M.; Nührenberg, T.; Trenk, D.; Neumann, F.-J.; Hochholzer, W. Comparison of Immature Platelet Count to Established Predictors of Platelet Reactivity During Thienopyridine Therapy. J. Am. Coll. Cardiol. 2016, 68, 286–293. [Google Scholar] [CrossRef] [PubMed]
- Vaughan, J.; Wiggill, T.; Munster, M. Immature platelet fraction levels in a variety of bone marrow pathologies in South African HIV-positive patients with thrombocytopenia. Hematology 2014, 19, 417–423. [Google Scholar] [CrossRef] [PubMed]
- Dadu, T.; Sehgal, K.; Joshi, M.; Khodaiji, S. Evaluation of the immature platelet fraction as an indicator of platelet recovery in dengue patients. Int. J. Lab. Hematol. 2014, 36, 499–504. [Google Scholar] [CrossRef]
- Ratsch, U.; Kaiser, T.; Stepan, H.; Jank, A. Evaluation of bone marrow function with immature platelet fraction in normal pregnancy. Pregnancy Hypertens. 2017, 10, 70–73. [Google Scholar] [CrossRef]
- Bernstein, U.; Kaiser, T.; Stepan, H.; Jank, A. The immature platelet fraction in hypertensive disease during pregnancy. Arch. Gynecol. Obs. 2019, 299, 1537–1543. [Google Scholar] [CrossRef]
- Zucker, M.L.; Hagedorn, C.H.; Murphy, C.A.; Stanley, S.; Reid, K.J.; Skikne, B.S. Mechanism of thrombocytopenia in chronic hepatitis C as evaluated by the immature platelet fraction. Int. J. Lab. Hematol. 2012, 34, 525–532. [Google Scholar] [CrossRef]
- Dou, J.; Lou, Y.; Wu, J.; Lu, Y.; Jin, Y. Thrombocytopenia in patients with hepatitis B virus-related chronic hepatitis: Evaluation of the immature platelet fraction. Platelets 2014, 25, 399–404. [Google Scholar] [CrossRef]


{kind=link}
| Route | Blood Volume | Collection Tool | Endpoint | Notes |
|---|---|---|---|---|
| Retrobulbar venous | ~0.5 mL | ~1.5 cm long glass capillary. | Sacrifice the animal. | Blood contact with glass can activate platelets. |
| Cardiac puncture | ~1 mL | 21 G needle and a syringe. | Sacrifice the animal. | Tearing of the heart muscle can lead to thrombin generation and platelet activation. |
| Inferior vena cava | ~1.2 mL | 22–27 G needle and a syringe. | Sacrifice the animal. | This method probably results in the least platelet activation. |
| Tail veins | Up to ~50 µL | Horizontal incision in the tail vein. | Suggested for multiple blood collection. | ‘Milking’ the tail should be avoided to avoid higher erythrocyte and leukocyte count in the sample. |
| Proposed Clinical Applications | Intended Goal | Reference |
|---|---|---|
| Thrombocytopenia | Differentiating platelets hypoproduction from accelerated destruction | [44,45,46,47,48] |
| Bone marrow/stem cells transplantation or chemotherapy | Predicting platelet recovery | [49,50,51,52,53] |
| Myelodysplastic syndromes | Clinical evaluation and assessing prognosis | [49,50] |
| Cardiovascular diseases | Assessing the role of platelet activation in prognosis | [51,52,53] |
| Antiplatelet therapy | Predicting treatment response | [52,54] |
| Infectious diseases | Early diagnosis | [55] |
| Sepsis | Predicting sepsis in critically ill patients | [56,57] |
| Pregnancy complications | Monitoring preeclampsia | [58] |
| Liver diseases | Differential diagnosis between cirrhosis and chronic hepatitis | [59,60,61] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hamad, M.A.; Schanze, N.; Schommer, N.; Nührenberg, T.; Duerschmied, D. Reticulated Platelets—Which Functions Have Been Established by In Vivo and In Vitro Data? Cells 2021, 10, 1172. https://doi.org/10.3390/cells10051172
Hamad MA, Schanze N, Schommer N, Nührenberg T, Duerschmied D. Reticulated Platelets—Which Functions Have Been Established by In Vivo and In Vitro Data? Cells. 2021; 10(5):1172. https://doi.org/10.3390/cells10051172
Chicago/Turabian StyleHamad, Muataz Ali, Nancy Schanze, Nicolas Schommer, Thomas Nührenberg, and Daniel Duerschmied. 2021. "Reticulated Platelets—Which Functions Have Been Established by In Vivo and In Vitro Data?" Cells 10, no. 5: 1172. https://doi.org/10.3390/cells10051172
APA StyleHamad, M. A., Schanze, N., Schommer, N., Nührenberg, T., & Duerschmied, D. (2021). Reticulated Platelets—Which Functions Have Been Established by In Vivo and In Vitro Data? Cells, 10(5), 1172. https://doi.org/10.3390/cells10051172