Retinoic Acid Receptors and the Control of Positional Information in the Regenerating Axolotl Limb

Abstract
:1. Introduction
2. Materials and Methods
2.1. Axolotl Care
2.1.1. Axolotl Procurement and Husbandry
2.1.2. Limb Amputations
2.1.3. Systemic RAR Agonist Treatment
2.2. Solution Preparation
2.2.1. Victoria Blue Stain
2.2.2. RAR Agonist Solutions
2.3. Limb Analysis
2.3.1. Limb Collection and Staining
2.3.2. Duplication Scoring
2.4. RNA Sequencing
2.4.1. RNA Extraction
2.4.2. Library Construction and RNA Sequencing
2.4.3. Read Processing and Quality Control
2.4.4. Data Processing and Visualization
2.5. RNA-Seq Analysis
2.5.1. Transcriptome Assembly
2.5.2. Differential Expression Analysis
2.5.3. Transcript Annotation
2.5.4. Pathway Analysis
2.6. Data
3. Results
3.1. Human and Axolotl RARs Are Homologous
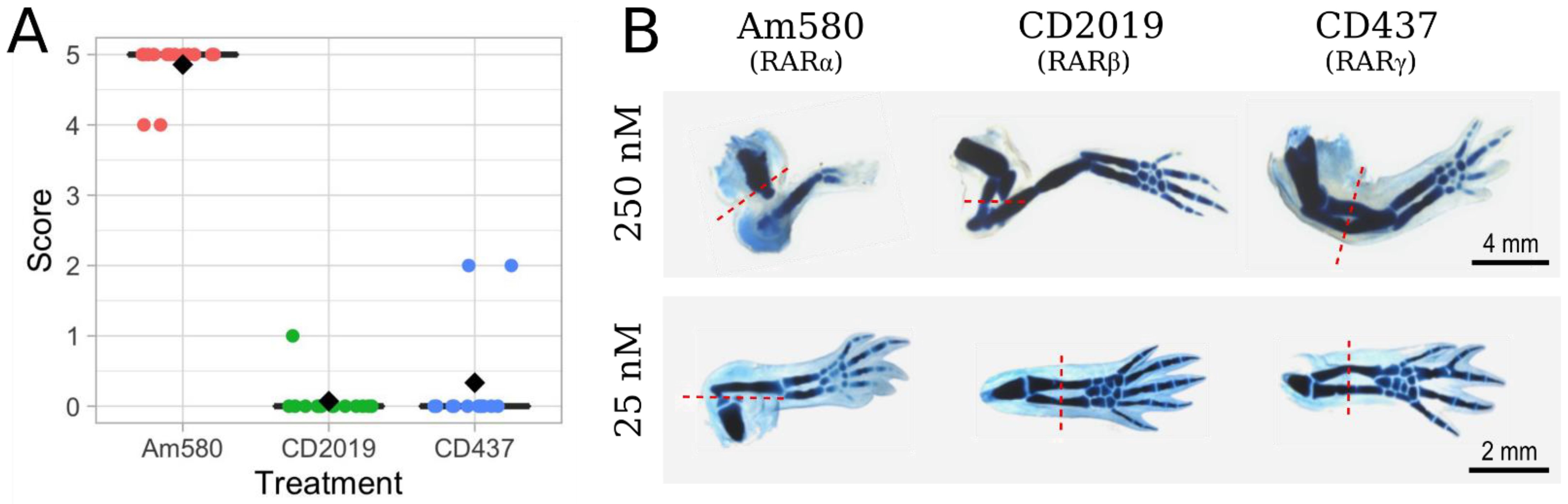
3.2. Effects of Selective RAR Agonists on Limb Regeneration
3.3. Systemic Treatment of 25 nM Am580 Consistently Respecifies Positional Information
3.4. RNA-Sequencing of Blastema Mesenchyme Tissue Following Am580 Treatment
3.4.1. Systemic Am580 Treatment
3.4.2. RNA-Sequencing Metrics
3.4.3. Transcriptome Assembly and Differential Expression Analysis
3.5. Differentially Expressed Genes
3.5.1. Integrin Signaling
3.5.2. Extracellular Matrix Genes
3.5.3. Cell Membrane and Secretory Protein Coding Genes
3.5.4. Unknown Genes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Crawford, K.; Stocum, D.L. Retinoic acid coordinately proximalizes regenerate pattern and blastema differential affinity in axolotl limbs. Development 1988, 102, 687–698. [Google Scholar] [CrossRef] [PubMed]
- Maden, M.; Turner, R.N. Supernumerary limbs in the axolotl. Nature 1978, 237, 232–235. [Google Scholar] [CrossRef] [PubMed]
- Nardi, J.B.; Stocum, D.L. Surface properties of regenerating limb cells: Evidence for gradation along the proximodistal axis. Differentiation 1983, 25, 27–31. [Google Scholar] [CrossRef]
- Maden, M. Vitamin A and pattern formation in the regenerating limb. Nature 1982, 295, 672–675. [Google Scholar] [CrossRef] [PubMed]
- Maden, M. The effect of vitamin A on the regenerating amphibian limb. J. Embryol. Exp. Morphol. 1983, 77, 273–295. [Google Scholar]
- da Silva, S.M.; Gates, P.B.; Brockes, J.P. The newt ortholog of CD59 is implicated in proximodistal identity during amphibian limb regeneration. Dev. Cell 2002, 3, 547–555. [Google Scholar] [CrossRef]
- Echeverri, K.; Tanaka, E.M. Proximodistal patterning during limb regeneration. Dev. Biol. 2005, 279, 391–401. [Google Scholar] [CrossRef]
- Blassberg, R.A.; Garza-Garcia, A.; Janmohamed, A.; Gates, P.B.; Brockes, J.P. Functional convergence of signalling by GPI-anchored and anchorless forms of a salamander protein implicated in limb regeneration. J. Cell Sci. 2011, 124, 47–56. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, M.; Singhal, P.; Piet, J.W.; Shefelbine, S.J.; Maden, M.; Voss, S.R.; Monaghan, J.R. Retinoic acid receptor regulation of epimorphic and homeostatic regeneration in the axolotl. Development 2017, 144, 601–611. [Google Scholar] [CrossRef] [Green Version]
- Delescluse, C.; Cavey, M.; Martin, B.; Bernard, B.; Reichert, U.; Maignan, J.; Darmon, M.; Shroot, B. Selective high affinity retinoic acid receptor alpha or beta-gamma ligands. Mol. Pharm. 1991, 40, 556–562. [Google Scholar]
- Martin, B.; Bernardon, J.; Cavey, M.; Bernard, B.; Carlavan, I.; Charpentier, B.; Pilgrim, W.; Shroot, B.; Reichert, U. Selective synthetic ligands for human nuclear retinoic acid receptors. Ski. Pharm. 1992, 5, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 1 January 2010).
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing. Available online: https://www.R-project.org/ (accessed on 1 January 2019).
- RStudio Team. RStudio: Integrated Development for R. Available online: http://www.rstudio.com/ (accessed on 1 January 2020).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Nowoshilow, S.; Schloissnig, S.; Fei, J.; Dahl, A.; Andy, W.; Pang, C.; Pippel, M.; Falcon, F.; Knapp, D.; Powell, S.; et al. The axolotl genome and the evolution of key tissue formation regulators. Nat. Publ. Gr. 2018, 554, 50–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R.; Data, G.P.; et al. The sequence alignment/map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [Green Version]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.-C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef] [Green Version]
- Pertea, M.; Kim, D.; Pertea, G.M.; Leek, J.T.; Salzberg, S.L. Transcript-level expression analysis of RNA- seq experiments with HISAT, StringTie and Transcript-level expression analysis of RNA-seq experiments with HISAT, StringTie and Ballgown. Nat. Protoc. 2016, 11, 1650–1667. [Google Scholar] [CrossRef]
- Pertea, G.; Pertea, M. GFF Utilities: GffRead and GffCompare. F1000Research 2020, 9, 1–20. [Google Scholar] [CrossRef]
- Liao, Y.; Smyth, G.K.; Shi, W. FeatureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [Green Version]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2009, 26, 139–140. [Google Scholar] [CrossRef] [Green Version]
- Ragsdale Jr, C.W.; Petkovich, M.; Gates, P.B.; Chambon, P.; Brockes, J.P. Identification of a novel retinoic acid receptor in regenerative tissues of the newt. Nature 1989, 341, 654–657. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.J.; Putta, S.; Walker, J.A.; Kump, D.K.; Samuels, A.K.; Monaghan, J.R.; Weisrock, D.W.; Staben, C.; Voss, S.R. Sal-Site: Integrating new and existing ambystomatid salamander research and informational resources. BMC Genom. 2005, 6, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Schilthuis, J.G.; Gann, A.; Brockes, J.P. Chimeric retinoic acid/thyroid hormone receptors implicate RAR-alpha 1 as mediating growth inhibition by retinoic acid. Embo J. 1993, 12, 3459–3466. [Google Scholar] [CrossRef] [PubMed]
- Pecorino, L.T.; Lo, D.C.; Brockes, J.P. Isoform-specific induction of a retinoid-responsive antigen after biolistic transfection of chimaeric retinoic acid / thyroid hormone receptors into a regenerating limb. Development 1994, 120, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Kastner, P.; Mark, M.; Chambon, P. Nonsteroid nuclear receptors: What are genetic studies telling us about their role in real life? Cell 1995, 83, 859–869. [Google Scholar] [CrossRef] [Green Version]
- Phan, A.Q.; Lee, J.; Oei, M.; Flath, C.; Hwe, C.; Mariano, R.; Vu, T.; Shu, C.; Dinh, A.; Simkin, J.; et al. Positional information in axolotl and mouse limb extracellular matrix is mediated via heparan sulfate and fibroblast growth factor during limb regeneration in the axolotl (Ambystoma mexicanum). Regeneration 2015, 2, 182–201. [Google Scholar] [CrossRef]
- Maden, M.; Chambers, D.; Monaghan, J. Retinoic acid and the genetics of positional information. In Regenerative Engineering and Developmental Biology: Principles and Applications; Gardiner, D.M., Ed.; CRC Press: Boca Raton, FL, USA, 2017; Chapter 7; pp. 163–180. [Google Scholar]
- Maden, M. Does vitamin A act on pattern formation via the epidermis or the mesenchyme? J. Exp. Zool. 1984, 230, 387–392. [Google Scholar] [CrossRef]
- Mercader, N.; Leonardo, E.; Piedra, M.E.; Martínez, A.C.; Ros, M.Á.; Torres, M. Opposing RA and FGF signals control proximodistal vertebrate limb development through regulation of Meis genes. Development 2000, 127, 3961–3970. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.C.; Eichele, G.; Thaller, C. Ligand-bound RXR can mediate retinoid signal transduction during embryogenesis. Development 1997, 124, 195–203. [Google Scholar] [CrossRef]
- Taniguchi, Y.; Komatsu, N.; Moriuchi, T. Overexpression of the HOX4A (HOXD3) homeobox gene in human erythroleukemia HEL cells results in altered adhesive properties. Blood 1995, 85, 2786–2794. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Godwin, J.W.; Gates, P.B.; Acely Garza-Garcia, A.; Brockes, J.P. Molecular Basis for the Nerve Dependence of Limb Regeneration in an Adult Vertebrate. Science 2007, 318, 772–777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliviera, C.R.; Knapp, D.; Elewa, A.; Gonzalez Malagon, S.G.; Gates, P.B.; Petzhold, A.; Arce, H.; Cordoba, R.C.; Chara, C.; Tanaka, E.M.; et al. Tig1 regulates proximo-distal identity during salamander limb regeneration. bioRxiv 2021. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Transcript Name | UniProt ID | Gene Name | log2FC |
|---|---|---|---|
| AMEXTC_0340000164913_cytochrome | CP26A_CHICK | CP26A | 15.382878 |
| AMEXTC_0340000004000_LOC108712603 | MMP21_CYNPY | MMP21 | 8.68081722 |
| AMEXTC_0340000018651_retinoic | RARB_HUMAN | RARB | 8.34534428 |
| MSTRG.18535 | RS25_SHEEP | RPS25 | 6.96185493 |
| AMEXTC_0340000220729_hypothetical | APMAP_RAT | APMAP | 6.31133061 |
| MSTRG.12045 | ATTY_MOUSE | TAT | 6.13262763 |
| MSTRG.12046 | ATTY_MOUSE | TAT | 5.72243555 |
| MSTRG.2544 | - | - | 5.61165445 |
| AMEXTC_0340000053364_LOXe | LX15B_HUMAN | ALOX15B | 5.26551299 |
| AMEXTC_0340000039558_TAT | ATTY_MOUSE | TAT | 5.26351744 |
| mRNA01059 | - | - | 5.21247926 |
| AMEXTC_0340000032012_LOC108645441 | - | - | 5.15846088 |
| AMEXTC_0340000247464_ripply3.L | DSCR6_XENLA | RIPPLY3 | 5.05107637 |
| MSTRG.17898 | NDNF_HUMAN | NDNF | 5.04336304 |
| AMEXTC_0340000179221_LOC102944588 | GIMA4_HUMAN | GIMAP4 | 5.02378211 |
| AMEXTC_0340000012549_fibroblast | FGF8_CHICK | FGF8 | −4.8925275 |
| MSTRG.4054 | - | - | −4.9088476 |
| MSTRG.23812 | - | - | −4.9178374 |
| MSTRG.16115 | - | - | −4.966291 |
| MSTRG.23482 | RTJK_DROME | POL | −5.0176166 |
| MSTRG.4440 | TPH_XENLA | TPH | −5.2716375 |
| AMEXTC_0340000028898_LOC108413561 | RTJK_DROME | POL | −5.2869995 |
| AMEXTC_0340000081353_OCSTAMP | OCSTP_HUMAN | OCSTAMP | −5.3164526 |
| MSTRG.1117 | - | - | −5.5152443 |
| MSTRG.16116 | LIN1_NYCCO | LIN1 | −5.6941371 |
| MSTRG.13846 | - | - | −5.702535 |
| AMEXTC_0340000024275_LOC101872464 | B3A2_PONAB | SLC4A2 | −5.8790351 |
| AMEXTC_0340000007120_HTR6 | 5HT6R_PANTR | HTR6 | −6.3188017 |
| MSTRG.5087 | - | - | −6.5656262 |
| MSTRG.25599 | LORF2_HUMAN | LIN1 | −6.8496992 |
| Gene Symbol | Entrez Gene Name | log2 Ratio |
|---|---|---|
| CAV1 | caveolin 1 | 0.642 |
| ITGA2 | integrin subunit alpha 2 | 1.139 |
| ITGA4 | integrin subunit alpha 4 | 1.139 |
| ITGA5 | integrin subunit alpha 5 | 0.492 |
| ITGA6 | integrin subunit alpha 6 | 0.608 |
| ITGA2B | integrin subunit alpha 2b | 1.001 |
| ITGAV | integrin subunit alpha V | −1.224 |
| ITGAX | integrin subunit alpha X | 1.723 |
| ITGB2 | integrin subunit beta 2 | −0.904 |
| ITGB3 | integrin subunit beta 3 | −1.307 |
| Transcript | UniProt ID | Gene Name | log2 FC |
|---|---|---|---|
| AMEXTC_0340000038644_LOC108803933 | P2RY1_MOUSE | P2RY1 | 4.48 |
| AMEXTC_0340000062820_GEM | GEM_HUMAN | GEM | 4.32 |
| MSTRG.21040 | HRH2_MOUSE | HRH2 | 3.32 |
| MSTRG.22927 | TLR4_PIG | TLR4 | 3.09 |
| AMEXTC_0340000034284_LOC102448090 | M4A4A_HUMAN | MS4A4A | 3.05 |
| AMEXTC_0340000013374_htr2b | 5HT2B_HUMAN | HTR2B | 2.93 |
| AMEXTC_0340000062083_FGD2 | FGD2_MOUSE | FGD2 | 2.83 |
| MSTRG.8675 | FGD2_HUMAN | FGD2 | 2.71 |
| MSTRG.3167 | FSHR_CAIMO | FSHR | 2.47 |
| AMEXTC_0340000014713_EPHA5 | EPHA5_HUMAN | EPHA5 | 2.4 |
| AMEXTC_0340000042498_ADRA2A | ADA2A_PIG | ADRA2A | 2.3 |
| AMEXTC_0340000030234_LOC109141183 | UNC5A_HUMAN | UNC5A | 2.24 |
| AMEXTC_0340000057843_LOC108718813 | MRC1_HUMAN | MRC1 | 2.2 |
| AMEXTC_0340000156303_BLNK | BLNK_CHICK | BLNK | 2.12 |
| AMEXTC_0340000062611_RGR | RGR_BOVIN | RGR | 2.06 |
| AMEXTC_0340000150384_cadherin | CAD17_MOUSE | CDH17 | 3.28 |
| AMEXTC_0340000048099_CDH4 | CADH4_CHICK | CDH4 | 3.11 |
| AMEXTC_0340000004184_LOC102348750 | CEAM5_MOUSE | CEACAM5 | 2.66 |
| AMEXTC_0340000028915_ICAM5 | ICAM5_MOUSE | ICAM5 | 2.44 |
| AMEXTC_0340000049914_Sell | LYAM2_HUMAN | SELE | 2.39 |
| AMEXTC_0340000220626_PLLP | PLLP_HUMAN | PLLP | 2.3 |
| AMEXTC_0340000220764_LOC101952950 | MYO10_BOVIN | MYO10 | 2.06 |
| Transcript | UniProt ID | Gene Name | log2FC |
|---|---|---|---|
| AMEXTC_0340000004000_LOC108712603 | MMP21_CYNPY | MMP21 | 8.68 |
| MSTRG.17898 | NDNF_HUMAN | NDNF | 5.04 |
| MSTRG.6144 | FCNV4_CERRY | - | 4.36 |
| AMEXTC_0340000030622_PROZ | PROZ_BOVIN | PROZ | 4.22 |
| AMEXTC_0340000250114_c4a.L | CO4_BOVIN | C4 | 3.87 |
| AMEXTC_0340000209130_LOC105403673 | CO5A1_MOUSE | COL5A1 | 3.63 |
| AMEXTC_0340000025257_HAPLN4 | HPLN4_MOUSE | HAPLN4 | 3.55 |
| MSTRG.15067 | VCO3_NAJKA | - | 3.42 |
| AMEXTC_0340000036400_LOC100552635 | FCNV1_VARKO | FCNV1_VARKO | 3.41 |
| AMEXTC_0340000044633_PKDCC | PKDCC_HUMAN | PKDCC | 3.29 |
| AMEXTC_0340000062173_Angiogenin | ANGI_MOUSE | ANG | 3.22 |
| MSTRG.6402 | MSMB_PIG | MSMB | 3.17 |
| AMEXTC_0340000056401_ostn | OSTN_HUMAN | OSTN | 3.09 |
| MSTRG.829 | IL12B_FELCA | IL12B | 2.85 |
| AMEXTC_0340000250115_complement | CO4_RAT | C4 | 2.6 |
| AMEXTC_0340000233392_CETP | CETP_CHICK | CETP | 2.6 |
| AMEXTC_0340000044847_Coiled-coil | WNT6_MOUSE | WNT6 | 2.48 |
| AMEXTC_0340000048200_TSPEAR | TSEAR_HUMAN | TSPEAR | 2.47 |
| MSTRG.13661 | IL18_CHICK | IL18 | 2.36 |
| MSTRG.10022 | IL8_CHICK | IL8 | 2.32 |
| AMEXTC_0340000000382_fibroblast | FGF2_BOVIN | FGF2 | 2.14 |
| MSTRG.1555 | GDF6A_DANRE | GDF6A | 2.14 |
| AMEXTC_0340000053380_COL27A1 | CO5A3_HUMAN | COL5A3 | 2.09 |
| AMEXTC_0340000232220_LOC107293967 | CHIT1_HUMAN | CHIT1 | 2.07 |
| AMEXTC_0340000170356_LOC102365657 | SFTPD_RAT | SFTPD | 2.05 |
| AMEXTC_0340000064262_ndnf | NDNF_XENTR | NDNF | 2.04 |
| MSTRG.5658 | SDF1_XENLA | SDF1 | 2.03 |
| AMEXTC_0340000010931_ADAMTS3 | ATS3_HUMAN | ADAMTS3 | 2.03 |
| MSTRG.22754 | HMCN1_HUMAN | HMCN1 | 2.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Polvadore, T.; Maden, M. Retinoic Acid Receptors and the Control of Positional Information in the Regenerating Axolotl Limb. Cells 2021, 10, 2174. https://doi.org/10.3390/cells10092174
Polvadore T, Maden M. Retinoic Acid Receptors and the Control of Positional Information in the Regenerating Axolotl Limb. Cells. 2021; 10(9):2174. https://doi.org/10.3390/cells10092174
Chicago/Turabian StylePolvadore, Trey, and Malcolm Maden. 2021. "Retinoic Acid Receptors and the Control of Positional Information in the Regenerating Axolotl Limb" Cells 10, no. 9: 2174. https://doi.org/10.3390/cells10092174
APA StylePolvadore, T., & Maden, M. (2021). Retinoic Acid Receptors and the Control of Positional Information in the Regenerating Axolotl Limb. Cells, 10(9), 2174. https://doi.org/10.3390/cells10092174