
Inference of Gene Regulatory Network Uncovers the Linkage between Circadian Clock and Crassulacean Acid Metabolism in Kalanchoë fedtschenkoi

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Time-Course Gene Expression Data
2.2. Identifying Candidate Core Clock Genes
2.3. Identify Core Clock-Regulators of Stomata-Related Genes
2.4. Gene Ontology Analysis
2.5. Comparative Analysis of Gene Expression
3. Results
3.1. Candidate Core Clock Transcription Factors in Kalanchoë fedtschenkoi
3.2. Morning Phased Candidate Core Clock Transcription Factors
3.3. Midday Phased Candidate Core Clock Transcription Factors
3.4. Evening Phased Candidate Core Clock Transcription Factors
3.5. Core Clock Regulation of Stomata-Related Genes in Kalanchoë fedtschenkoi
3.6. Regulation of Rescheduled Stomata-Related Genes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ehleringer, J.R.; Monson, R.K. Evolutionary and ecological aspects of photosynthetic pathway variation. Annu. Rev. Ecol. Syst. 1993, 24, 411–439. [Google Scholar] [CrossRef]
- Dai, A. Increasing drought under global warming in observations and models. Nat. Clim. Chang. 2013, 3, 52–58. [Google Scholar] [CrossRef]
- Cook, B.I.; Smerdon, J.E.; Seager, R.; Coats, S. Global warming and 21st century drying. Clim. Dyn. 2014, 43, 2607–2627. [Google Scholar] [CrossRef] [Green Version]
- Borland, A.M.; Hartwell, J.; Weston, D.J.; Schlauch, K.A.; Tschaplinski, T.J.; Tuskan, G.A.; Yang, X.; Cushman, J.C. Engineering crassulacean acid metabolism to improve water-use efficiency. Trends Plant Sci. 2014, 19, 327–338. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Cushman, J.C.; Borland, A.M.; Edwards, E.J.; Wullschleger, S.D.; Tuskan, G.A.; Owen, N.A.; Griffiths, H.; Smith, J.A.C.; De Paoli, H.C.; et al. A roadmap for research on crassulacean acid metabolism (CAM) to enhance sustainable food and bioenergy production in a hotter, drier world. New Phytol. 2015, 207, 491–504. [Google Scholar] [CrossRef]
- Hartwell, J. The co-ordination of central plant metabolism by the circadian clock. Biochem. Soc. Trans. 2005, 33, 945–948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartwell, J. Endogenous Plant Rhythms; Wiley-Blackwell Publishing Ltd: Hokoken, NJ, USA, 2007; The Circadian Clock in CAM Plants; pp. 211–236. [Google Scholar]
- Moseley, R.C.; Tuskan, G.A.; Yang, X. Comparative genomics analysis provides new insight into molecular basis of stomatal movement in Kalanchoë fedtschenkoi. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef]
- Harmer, S.L. The circadian system in higher plants. Annu. Rev. Plant Biol. 2009, 60, 357–377. [Google Scholar] [CrossRef] [Green Version]
- Hubbard, K.E.; Webb, A.A.R. Rhythms in Plants: Dynamic Responses in a Dynamic Environment; Springer-Verlag GmbH: Berlin/Heidelberg, Germany, 2015; Circadian rhythms in stomata: Physiological and molecular aspects; pp. 231–255. [Google Scholar]
- Kholodenko, B.N. Cell-signalling dynamics in time and space. Nat. Rev. Mol. Cell Biol. 2006, 7, 165–176. [Google Scholar] [CrossRef]
- Tiana, G.; Krishna, S.; Pigolotti, S.; Jensen, M.H.; Sneppen, K. Oscillations and temporal signalling in cells. Phys. Biol. 2007, 4. [Google Scholar] [CrossRef] [Green Version]
- Young, M.W.; Kay, S.A. Time zones: A comparative genetics of circadian clocks. Nat. Rev. Genet. 2001, 2, 702–715. [Google Scholar] [CrossRef]
- McClung, C.R. Plant circadian rhythms. Plant Cell 2006, 18, 792–803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michael, T.P.; Mockler, T.C.; Breton, G.; McEntee, C.; Byer, A.; Trout, J.D.; Hazen, S.P.; Shen, R.; Priest, H.D.; Sullivan, C.M.; et al. Network discovery pipeline elucidates conserved time-of-day-specific cis-regulatory modules. PLoS Genet. 2008, 4, e14. [Google Scholar] [CrossRef] [Green Version]
- Filichkin, S.A.; Breton, G.; Priest, H.D.; Dharmawardhana, P.; Jaiswal, P.; Fox, S.E.; Michael, T.P.; Chory, J.; Kay, S.A.; Mockler, T.C. Global profiling of rice and poplar transcriptomes highlights key conserved Circadian-controlled pathways and cis-regulatory modules. PLoS ONE 2011, 6, e16907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ming, R.; VanBuren, R.; Wai, C.M.; Tang, H.; Schatz, M.C.; Bowers, J.E.; Lyons, E.; Wang, M.L.; Chen, J.; Biggers, E.; et al. The pineapple genome and the evolution of CAM photosynthesis. Nat. Genet. 2015, 47, 1435–1442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abraham, P.E.; Yin, H.; Borland, A.M.; Weighill, D.; Lim, S.D.; De Paoli, H.C.; Engle, N.; Jones, P.C.; Agh, R.; Weston, D.J.; et al. Transcript, protein and metabolite temporal dynamics in the CAM plant Agave. Nat. Plants 2016, 2, 16178. [Google Scholar] [CrossRef] [Green Version]
- Higashi, T.; Tanigaki, Y.; Takayama, K.; Nagano, A.J.; Honjo, M.N.; Fukuda, H. Detection of diurnal variation of tomato transcriptome through the molecular timetable method in a sunlight-type plant factory. Front. Plant Sci. 2016, 7, 87. [Google Scholar] [CrossRef] [Green Version]
- Koda, S.; Onda, Y.; Matsui, H.; Takahagi, K.; Yamaguchi-Uehara, Y.; Shimizu, M.; Inoue, K.; Yoshida, T.; Sakurai, T.; Honda, H.; et al. Diurnal transcriptome and gene network represented through sparse modeling in brachypodium distachyon. Front. Plant Sci. 2017, 8, 2055. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Hu, R.; Yin, H.; Jenkins, J.; Shu, S.; Tang, H.; Liu, D.; Weighill, D.A.; Cheol Yim, W.; Ha, J.; et al. The Kalanchoë genome provides insights into convergent evolution and building blocks of crassulacean acid metabolism. Nat. Commun. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- McGoff, K.A.; Guo, X.; Deckard, A.; Kelliher, C.M.; Leman, A.R.; Francey, L.J.; Hogenesch, J.B.; Haase, S.B.; Harer, J.L. The Local Edge Machine: Inference of dynamic models of gene regulation. Genome Biol. 2016, 17, 214. [Google Scholar] [CrossRef] [Green Version]
- Cummins, B.; Gedeon, T.; Harker, S.; Mischaikow, K. Database of Dynamic Signatures Generated by Regulatory Networks (DSGRN). Lect. Notes Comput. Sci. (including Subser. Lect. Notes Artif. Intell. Lect. Notes Bioinformatics) 2017, 10545 LNBI, 300–308. [Google Scholar] [CrossRef]
- Grundy, J.; Stoker, C.; Carré, I.A. Circadian regulation of abiotic stress tolerance in plants. Front. Plant Sci. 2015, 6, 648. [Google Scholar] [CrossRef] [PubMed]
- Mockler, T.C.; Michael, T.P.; Priest, H.D.; Shen, R.; Sullivan, C.M.; Givan, S.A.; Mcentee, C.; Kay, S.A.; Chory, J. The diurnal project: Diurnal and circadian expression profiling, model-based pattern matching, and promoter analysis. In Proceedings of the Cold Spring Harbor Symposia on Quantitative Biology. Cold Spring Harb. Symp. Quant. Biol. 2007, 72, 353–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, B.; Zhang, P.; Chen, X.; Liu, L.; Wang, Y.; He, S.; Chen, R. Predicting housekeeping genes based on fourier analysis. PLoS One 2011, 6, e21012. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.; Azencott, R.; Zhao, Y. Modeling miRNA-mRNA interactions: Fitting chemical kinetics equations to microarray data. BMC Syst. Biol. 2014, 8, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moseley, R.C.; Mewalal, R.; Motta, F.; Tuskan, G.A.; Haase, S.; Yang, X. Conservation and diversification of circadian rhythmicity between a model crassulacean acid metabolism plant kalanchoë fedtschenkoi and a model C3 photosynthesis plant Arabidopsis thaliana. Front. Plant Sci. 2018, 9, 1757. [Google Scholar] [CrossRef]
- Anafi, R.C.; Lee, Y.; Sato, T.K.; Venkataraman, A.; Ramanathan, C.; Kavakli, I.H.; Hughes, M.E.; Baggs, J.E.; Growe, J.; Liu, A.C.; et al. Machine Learning Helps Identify CHRONO as a Circadian Clock Component. PLoS Biol. 2014, 12, e1001840. [Google Scholar] [CrossRef]
- Sharma, A.; Wai, C.M.; Ming, R.; Yu, Q. Diurnal cycling transcription factors of pineapple revealed by genome-wide annotation and global transcriptomic analysis. Genome Biol. Evol. 2017, 9, 2170–2190. [Google Scholar] [CrossRef] [Green Version]
- de Lichtenberg, U.; Jensen, L.J.; Fausbøll, A.; Jensen, T.S.; Bork, P.; Brunak, S. Comparison of computational methods for the identification of cell cycle-regulated genes. Bioinformatics 2005, 21, 1164–1171. [Google Scholar] [CrossRef]
- pyDL. Available online: https://gitlab.com/biochron/pydl (accessed on 12 July 2017).
- ME, H.; JB, H.; K, K. JTK_CYCLE: An efficient nonparametric algorithm for detecting rhythmic components in genome-scale data sets. J. Biol. Rhythms 2010, 25, 372–380. [Google Scholar] [CrossRef]
- PyJTK. Available online: https://gitlab.com/biochron/pyjtk (accessed on 12 July 2017).
- Motta, F.C.; Moseley, R.C.; Cummins, B.; Deckard, A.; Haase, S.B. Conservation of dynamic characteristics of transcriptional regulatory elements in periodic biological processes. bioRxiv 2020. [Google Scholar] [CrossRef]
- LEMpy. Available online: https://gitlab.com/biochron/lempy (accessed on 25 July 2017).
- Nohales, M.A.; Kay, S.A. Molecular mechanisms at the core of the plant circadian oscillator. Nat. Struct. Mol. Biol. 2016, 23, 1061–1069. [Google Scholar] [CrossRef]
- Nusinow, D.A.; Helfer, A.; Hamilton, E.E.; King, J.J.; Imaizumi, T.; Schultz, T.F.; Farré, E.M.; Kay, S.A. The ELF4-ELF3-"LUX complex links the circadian clock to diurnal control of hypocotyl growth. Nature 2011, 475, 398–404. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, T.; Nomoto, Y.; Oka, H.; Kitayama, M.; Takeuchi, A.; Tsubouchi, M.; Yamashino, T. Ambient temperature signal feeds into the circadian clock transcriptional circuitry through the EC night-time repressor in arabidopsis thaliana. Plant Cell Physiol. 2014, 55, 958–976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helfer, A.; Nusinow, D.A.; Chow, B.Y.; Gehrke, A.R.; Bulyk, M.L.; Kay, S.A. LUX ARRHYTHMO encodes a nighttime repressor of circadian gene expression in the Arabidopsis core clock. Curr. Biol. 2011, 21, 126–133. [Google Scholar] [CrossRef] [Green Version]
- Goodstein, D.M.; Shu, S.; Howson, R.; Neupane, R.; Hayes, R.D.; Fazo, J.; Mitros, T.; Dirks, W.; Hellsten, U.; Putnam, N.; et al. Phytozome: A comparative platform for green plant genomics. Nucleic Acids Res. 2012, 40. [Google Scholar] [CrossRef]
- Bindea, G.; Mlecnik, B.; Hackl, H.; Charoentong, P.; Tosolini, M.; Kirilovsky, A.; Fridman, W.H.; Pagès, F.; Trajanoski, Z.; Galon, J. ClueGO: A Cytoscape plug-in to decipher functionally grouped gene ontology and pathway annotation networks. Bioinformatics 2009, 25, 1091–1093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sture Holm A Simple Sequentially Rejective Multiple Test Procedure. Scand. J. Stat. 1979, 6, 65–70.
- SciPy. Available online: https://scipy.org/ (accessed on 15 November 2017).
- Zhang, R.; Lahens, N.F.; Ballance, H.I.; Hughes, M.E.; Hogenesch, J.B. A circadian gene expression atlas in mammals: Implications for biology and medicine. Proc. Natl. Acad. Sci. USA 2014, 111, 16219–16224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jakoby, M.; Weisshaar, B.; Dröge-Laser, W.; Vicente-Carbajosa, J.; Tiedemann, J.; Kroj, T.; Parcy, F. bZIP transcription factors in Arabidopsis. Trends Plant Sci. 2002, 7, 106–111. [Google Scholar] [CrossRef]
- Gibalová, A.; Reňák, D.; Matczuk, K.; Dupl’Áková, N.; Cháb, D.; Twell, D.; Honys, D. AtbZIP34 is required for Arabidopsis pollen wall patterning and the control of several metabolic pathways in developing pollen. Plant Mol. Biol. 2009, 70, 581–601. [Google Scholar] [CrossRef]
- Fornara, F.; Panigrahi, K.C.S.; Gissot, L.; Sauerbrunn, N.; Rühl, M.; Jarillo, J.A.; Coupland, G. Arabidopsis DOF Transcription Factors Act Redundantly to Reduce CONSTANS Expression and Are Essential for a Photoperiodic Flowering Response. Dev. Cell 2009, 17, 75–86. [Google Scholar] [CrossRef] [Green Version]
- Nakamichi, N.; Kita, M.; Niinuma, K.; Ito, S.; Yamashino, T.; Mizoguchi, T.; Mizuno, T. Arabidopsis clock-associated pseudo-response regulators PRR9, PRR7 and PRR5 coordinately and positively regulate flowering time through the canonical CONSTANS-dependent photoperiodic pathway. Plant Cell Physiol. 2007, 48, 822–832. [Google Scholar] [CrossRef] [Green Version]
- Niwa, Y.; Ito, S.; Nakamichi, N.; Mizoguchi, T.; Niinuma, K.; Yamashino, T.; Mizuno, T. Genetic linkages of the circadian clock-associated genes, TOC1, CCA1 and LHY, in the photoperiodic control of flowering time in Arabidopsis thaliana. Plant Cell Physiol. 2007, 48, 925–937. [Google Scholar] [CrossRef]
- Ito, S.; Niwa, Y.; Nakamichi, N.; Kawamura, H.; Yamashino, T.; Mizuno, T. Insight into missing genetic links between two evening-expressed pseudo-response regulator genes TOC1 and PRR5 in the circadian clock-controlled circuitry in Arabidopsis thaliana. Plant Cell Physiol. 2008, 49, 201–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simmons Kovacs, L.A.; Mayhew, M.B.; Orlando, D.A.; Jin, Y.; Li, Q.; Huang, C.; Reed, S.I.; Mukherjee, S.; Haase, S.B. Cyclin-dependent kinases are regulators and effectors of oscillations driven by a transcription factor network. Mol. Cell 2012, 45, 669–679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamichi, N.; Kiba, T.; Kamioka, M.; Suzukie, T.; Yamashino, T.; Higashiyama, T.; Sakakibara, H.; Mizuno, T. Transcriptional repressor PRR5 directly regulates clock-output pathways. Proc. Natl. Acad. Sci. USA 2012, 109, 17123–17128. [Google Scholar] [CrossRef] [Green Version]
- Seaton, D.D.; Smith, R.W.; Song, Y.H.; MacGregor, D.R.; Stewart, K.; Steel, G.; Foreman, J.; Penfield, S.; Imaizumi, T.; Millar, A.J.; et al. Linked circadian outputs control elongation growth and flowering in response to photoperiod and temperature. Mol. Syst. Biol. 2015, 11, 776. [Google Scholar] [CrossRef]
- Imaizumi, T.; Schultz, T.F.; Harmon, F.G.; Ho, L.A.; Kay, S.A. Plant science: FKF1 F-box protein mediates cyclic degradation of a repressor of CONSTANS in Arabidopsis. Science (80-) 2005, 309, 293–297. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.H.; Smith, R.W.; To, B.J.; Millar, A.J.; Imaizumi, T. FKF1 conveys timing information for CONSTANS stabilization in photoperiodic flowering. Science (80-) 2012, 336, 1045–1049. [Google Scholar] [CrossRef] [Green Version]
- Keurentjes, J.J.B.; Fu, J.; Terpstra, I.R.; Garcia, J.M.; van den Ackerveken, G.; Snoek, L.B.; Peeters, A.J.M.; Vreugdenhil, D.; Koornneef, M.; Jansen, R.C. Regulatory network construction in Arabidopsis by using genome-wide gene expression quantitative trait loci. Proc. Natl. Acad. Sci. USA 2007, 104, 1708–1713. [Google Scholar] [CrossRef] [Green Version]
- Mandadi, K.K.; Misra, A.; Ren, S.; McKnight, T.D. BT2, a BTB Protein, Mediates Multiple Responses to Nutrients, Stresses, and Hormones in Arabidopsis. Plant Physiol. 2009, 150, 1930–1939. [Google Scholar] [CrossRef] [Green Version]
- Araus, V.; Vidal, E.A.; Puelma, T.; Alamos, S.; Mieulet, D.; Guiderdoni, E.; Gutiérrez, R.A. Members of BTB gene family of scaffold proteins suppress nitrate uptake and nitrogen use efficiency. Plant Physiol. 2016, 171, 1523–1532. [Google Scholar] [CrossRef] [Green Version]
- Bencivenga, S.; Simonini, S.; Benková, E.; Colombo, L. The transcription factors BEL1 and SPL are required for cytokinin and auxin signaling during ovule development in Arabidopsis. Plant Cell 2012, 24, 2886–2897. [Google Scholar] [CrossRef] [Green Version]
- Fonseca, S.; Fernández-Calvo, P.; Fernández, G.M.; Díez-Díaz, M.; Gimenez-Ibanez, S.; López-Vidriero, I.; Godoy, M.; Fernández-Barbero, G.; Van Leene, J.; De Jaeger, G.; et al. bHLH003, bHLH013 and bHLH017 are new targets of JAZ repressors negatively regulating JA responses. PLoS ONE 2014, 9, e86182. [Google Scholar] [CrossRef] [Green Version]
- Nakata, M.; Mitsuda, N.; Herde, M.; Koo, A.J.K.; Moreno, J.E.; Suzuki, K.; Howe, G.A.; Ohme-Takagi, M. A bHLH-type transcription factor, ABA-INDUCIBLE BHLH-TYPE TRANSCRIPTION FACTOR/JA-ASSOCIATED MYC2-LIKE1, acts as a repressor to negatively regulate jasmonate signaling in Arabidopsis. Plant Cell 2013, 25, 1641–1656. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Sun, J.; Xu, Y.; Jiang, H.; Wu, X.; Li, C. The bHLH-type transcription factor AtAIB positively regulates ABA response in Arabidopsis. Plant Mol. Biol. 2007, 65, 655–665. [Google Scholar] [CrossRef]
- Hansen, L.L.; van Ooijen, G. Rapid analysis of circadian phenotypes in arabidopsis protoplasts transfected with a luminescent clock reporter. J. Vis. Exp. 2016, 17, e54586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.S.; Pandey, S.; Li, S.; Gookin, T.E.; Zhao, Z.; Albert, R.; Assmann, S.M. Common and unique elements of the ABA-regulated transcriptome of Arabidopsis guard cells. BMC Genom. 2011, 12, 1–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mustilli, A.C.; Merlot, S.; Vavasseur, A.; Fenzi, F.; Giraudat, J. Arabidopsis OST1 protein kinase mediates the regulation of stomatal aperture by abscisic acid and acts upstream of reactive oxygen species production. Plant Cell 2002, 14, 3089–3099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vahisalu, T.; Kollist, H.; Wang, Y.F.; Nishimura, N.; Chan, W.Y.; Valerio, G.; Lamminmäki, A.; Brosché, M.; Moldau, H.; Desikan, R.; et al. SLAC1 is required for plant guard cell S-type anion channel function in stomatal signalling. Nature 2008, 452, 487–491. [Google Scholar] [CrossRef] [PubMed]
- Sirichandra, C.; Gu, D.; Hu, H.; Davanture, M.; Lee, S.; Djaoui, M.; Valot, B.; Zivy, M.; Jeffrey Leung, S.M.; Kwak, J.M.; et al. Phosphorylation of the Arabidopsis AtrbohF NADPH oxidase by OST1 protein kinase. FEBS Lett. 2009, 583, 2982–2986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grondin, A.; Rodrigues, O.; Verdoucq, L.; Merlot, S.; Leonhardt, N.; Maurel, C. Aquaporins contribute to ABA-triggered stomatal closure through OST1-mediated phosphorylation. Plant Cell 2015, 27, 1945–1954. [Google Scholar] [CrossRef] [Green Version]
- Kinoshita, T.; Shimazaki, K.I. Blue light activates the plasma membrane H+-ATPase by phosphorylation of the C-terminus in stomatal guard cells. EMBO J. 1999, 18, 5548–5558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inoue, S.I.; Kinoshita, T.; Matsumoto, M.; Nakayama, K.I.; Doi, M.; Shimazaki, K.I. Blue light-induced autophosphorylation of phototropin is a primary step for signaling. Proc. Natl. Acad. Sci. USA 2008, 105, 5626–5631. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Noguchi, K.; Ono, N.; Inoue, S.I.; Terashima, I.; Kinoshita, T. Overexpression of plasma membrane H+-ATPase in guard cells promotes light-induced stomatal opening and enhances plant growth. Proc. Natl. Acad. Sci. USA 2014, 111, 533–538. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
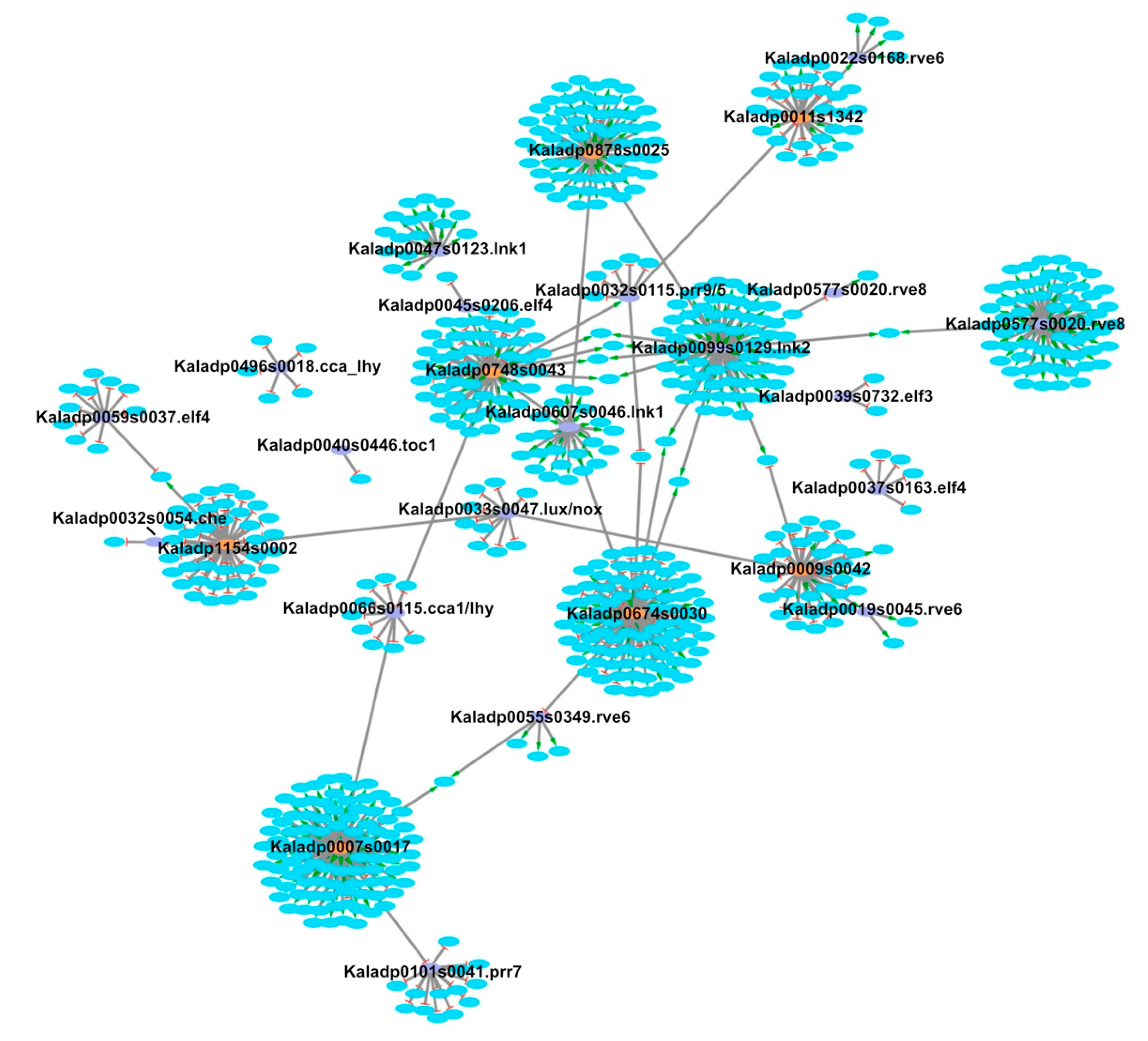{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Kafe Gene ID | Kafe Phase Call | Arth Ortholog ID | Arth Gene Symbol | Arth Gene Desc. | Arth Phase Call | Kafe Shift 1 (hrs) | Spear Shift 2 |
|---|---|---|---|---|---|---|---|
| Kaladp0748s0043 (CTF1) | 6 | AT5G41410 | BEL1 | POX (plant homeobox) family protein | 12 | −6 | 0.97 |
| Kaladp0007s0017 (CTF2) | 14 | AT2G46510 | JAM1 | ABA-inducible BHLH-type transcription factor | 12 | +2 | 0.91 |
| Kaladp0007s0017 (CTF2) | 14 | AT1G01260 | JAM2 | basic helix-loop-helix (bHLH) DNA-binding superfamily protein | 8 | +6 | 0.83 |
| Kaladp0011s1342 (CTF3) | 22 | AT2G42380 | BZIP34 | Basic-leucine zipper (bZIP) transcription factor family protein | 4 | −8 | 0.96 |
| Kaladp0011s1342 (CTF3) | 22 | AT3G58120 | BZIP61 | Basic-leucine zipper (bZIP) transcription factor family protein | 8 | −8 | 0.97 |
| Kaladp1154s0002 (CTF4) | 22 | AT3G47500 | CDF3 | cycling DOF factor 3 | 2 | −4 | 0.97 |
| Kaladp1154s0002 (CTF4) | 22 | AT5G62430 | CDF1 | cycling DOF factor 1 | 24 | −2 | 0.97 |
| Kaladp1154s0002 (CTF4) | 22 | AT5G39660 | CDF2 | cycling DOF factor 2 | 24 | −2 | 0.87 |
| Kaladp0674s0030 (CTF5) | 6 | AT5G63160 | BT1 | BTB and TAZ domain protein 1 | |||
| Kaladp0674s0030 (CTF5) | 6 | AT3G48360 | BT2 | BTB and TAZ domain protein 2 | 20 | +10 | 0.94 |
| Kaladp0878s0025 (CTF6) | 8 | AT1G07050 | CCT motif family protein | 12 | −2 | 0.92 | |
| Kaladp0009s0042 (CTF7) | 22 |
| Clock TF | Relationship 1 | Study | Target | Stomata-related 2 | Gene Description |
|---|---|---|---|---|---|
| Kaladp0748s0043 (CTF1) | Represses | [21] | Kaladp0059s0048 | New | aquaporin pip1-2 |
| Kaladp0748s0043 (CTF1) | Represses | [8] | Kaladp0062s0167 | New | receptor-like protein kinase haiku2 |
| Kaladp0007s0017 (CTF2) | Activates | [21] | Kaladp0011s0363 | New | trehalose-phosphate synthase |
| Kaladp0007s0017 (CTF2) | Activates | [8] | Kaladp0092s0084 | Known/GO | calcium-dependent protein kinase 26 |
| Kaladp0011s1342 (CTF3) | Represses | [21] | Kaladp0040s0264 | New | btb poz domain-containing protein npy2-like |
| Kaladp0011s1342 (CTF3) | Represses | [21] | Kaladp0008s0539 | New | mitogen-activated protein kinase |
| Kaladp0011s1342 (CTF3) | Represses | [21] | Kaladp0033s0113 | Known/GO | phototropin-2 |
| Kaladp0011s1342 (CTF3) | Represses | [8] | Kaladp0001s0016 | New | catalase isozyme 1 |
| Kaladp0011s1342 (CTF3) | Represses | [8] | Kaladp0093s0030 | New | homeobox-leucine zipper protein anthocyaninless 2 isoform x1 |
| Kaladp0059s0037 (ELF4) | Represses | [8] | Kaladp0062s0076 | New | 3-ketoacyl- synthase 19-like |
| Kaladp0047s0123 (LNK1) | Activates | [8] | Kaladp0008s0414 | New | cyclic nucleotide-gated ion channel 15 |
| Kaladp0060s0264 (LNK2) | Activates | [8] | Kaladp0024s0371 | New | pectin lyase-like superfamily protein isoform1 |
| Kaladp0099s0129 (LNK2) | Activates | [8] | Kaladp0042s0353 | Known | abscisic acid receptor pyl8-like |
| Kaladp0060s0264 (LNK2) | Activates | [8] | Kaladp0095s0634 | GO | mitogen-activated protein kinase homolog mmk2 |
| Kaladp0101s0041 (PRR7) | Represses | [8] | Kaladp0092s0115 | New | pleiotropic drug resistance protein 1-like |
| Kaladp0055s0349 (RVE6) | Activates | [8] | Kaladp0043s0103 | New | phospholipid-transporting atpase 3 |
| Kaladp0055s0349 (RVE6) | Activates | [8] | Kaladp0090s0003 | New | receptor-like protein kinase |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moseley, R.C.; Motta, F.; Tuskan, G.A.; Haase, S.B.; Yang, X. Inference of Gene Regulatory Network Uncovers the Linkage between Circadian Clock and Crassulacean Acid Metabolism in Kalanchoë fedtschenkoi. Cells 2021, 10, 2217. https://doi.org/10.3390/cells10092217
Moseley RC, Motta F, Tuskan GA, Haase SB, Yang X. Inference of Gene Regulatory Network Uncovers the Linkage between Circadian Clock and Crassulacean Acid Metabolism in Kalanchoë fedtschenkoi. Cells. 2021; 10(9):2217. https://doi.org/10.3390/cells10092217
Chicago/Turabian StyleMoseley, Robert C., Francis Motta, Gerald A. Tuskan, Steven B. Haase, and Xiaohan Yang. 2021. "Inference of Gene Regulatory Network Uncovers the Linkage between Circadian Clock and Crassulacean Acid Metabolism in Kalanchoë fedtschenkoi" Cells 10, no. 9: 2217. https://doi.org/10.3390/cells10092217
APA StyleMoseley, R. C., Motta, F., Tuskan, G. A., Haase, S. B., & Yang, X. (2021). Inference of Gene Regulatory Network Uncovers the Linkage between Circadian Clock and Crassulacean Acid Metabolism in Kalanchoë fedtschenkoi. Cells, 10(9), 2217. https://doi.org/10.3390/cells10092217