
Phosphorylation-Mediated Molecular Pathway Changes in Human Pituitary Neuroendocrine Tumors Identified by Quantitative Phosphoproteomics

Abstract
:1. Introduction
2. Materials and Methods
2.1. Tissue Specimen and Preparation of Protein Samples
2.2. Enzyme Hydrolysis and Peptide Quantification
2.3. TMT Labeling
2.4. TiO2 Enrichment of Phosphopeptides
2.5. LC-MS/MS Analysis of Enriched Phosphopeptides
2.6. Statistical Analysis and Bioinformatics
2.7. Immunoprecipitation and Western Blot Analyses of DPP Calnexin
3. Results
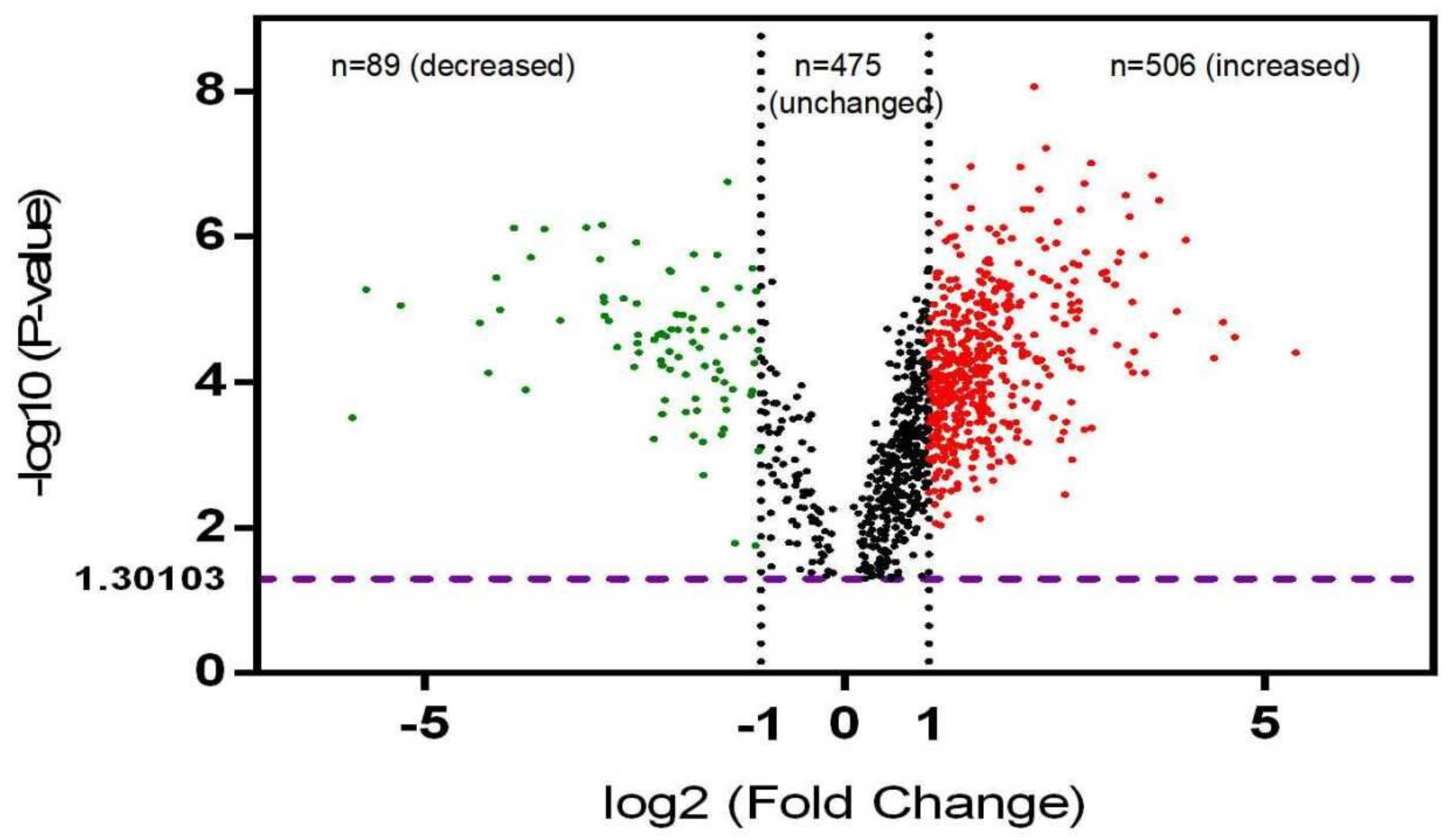
3.1. Differentially Phosphorylated Protein (DPP) Profiling in NF-PitNETs
3.2. Functional Characteristics of DPPs in NF-PitNETs
3.3. Phosphorylation-Involved Signaling Pathway Alterations in NF-PitNETs
3.4. Upstream Kinase Profiling Analysis of DPPs in NF-PitNETs
3.5. Verification of DPPs in NF-PitNETs Compared to Controls
4. Discussion
4.1. Phosphorylation-Mediated Biological Processes in NF-PitNETs
4.2. The Functions of Kinases and Their Corresponding Substrates Associated with Quantified Phosphoproteins
4.3. The Phosphorylation of Calnexin in NF-PitNETs
5. Strengths and Limitations
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lander, E.S.; Linton, L.M.; Birren, B.; Nusbaum, C.; Zody, M.C.; Baldwin, J.; Devon, K.; Dewar, K.; Doyle, M.; FitzHugh, W.; et al. Initial sequencing and analysis of the human genome. Nature 2001, 409, 860–921. [Google Scholar] [CrossRef] [Green Version]
- Zhan, X.; Desiderio, D.M. The use of variations in proteomes to predict, prevent, and personalize treatment for clinically nonfunctional pituitary adenomas. EPMA J. 2010, 1, 439–459. [Google Scholar] [CrossRef] [Green Version]
- Zhan, X.; Desiderio, D.M. Comparative proteomics analysis of human pituitary adenomas: Current status and future perspectives. Mass Spectrom. Rev. 2005, 24, 783–813. [Google Scholar] [CrossRef] [PubMed]
- Zhan, X.; Yang, H.; Peng, F.; Li, J.; Mu, Y.; Long, Y.; Cheng, T.; Huang, Y.; Li, Z.; Lu, M.; et al. How many proteins can be identified in a 2DE gel spot within an analysis of a complex human cancer tissue proteome? Electrophoresis 2018, 39, 965–980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Xu, M.L.; Li, B.Q.; Zhai, H.L.; Liu, J.J.; Li, S.Y. Prediction of phosphorylation sites based on Krawtchouk image moments. Proteins 2017, 85, 2231–2238. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, P.A.; Murray, B.W. Protein kinase biochemistry and drug discovery. Bioorg. Chem. 2011, 39, 192–210. [Google Scholar] [CrossRef] [PubMed]
- Nishi, H.; Shaytan, A.; Panchenko, A.R. Physicochemical mechanisms of protein regulation by phosphorylation. Front. Genet. 2014, 5, 270. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.J.; Hsieh, F.C.; Song, H.; Lin, J. Elevated phosphorylation and activation of PDK-1/AKT pathway in human breast cancer. Br. J. Cancer 2005, 93, 1372–1381. [Google Scholar] [CrossRef]
- Xie, W.; Liu, C.; Wu, D.; Li, Z.; Li, C.; Zhang, Y. Phosphorylation of kinase insert domain receptor by cyclin-dependent kinase 5 at serine 229 is associated with invasive behavior and poor prognosis in prolactin pituitary adenomas. Oncotarget 2016, 7, 50883–50894. [Google Scholar] [CrossRef] [Green Version]
- Zhan, X.; Desiderio, D.M. Editorial: Systems Biological Aspects of Pituitary Tumors. Front. Endocrinol. 2016, 7, 86. [Google Scholar] [CrossRef] [Green Version]
- Cheng, S.; Wu, J.; Li, C.; Li, Y.; Liu, C.; Li, G.; Li, W.; Hu, S.; Ying, X.; Zhang, Y. Predicting the regrowth of clinically non-functioning pituitary adenoma with a statistical model. J. Transl. Med. 2019, 17, 164. [Google Scholar] [CrossRef]
- Yamanaka, R.; Abe, E.; Sato, T.; Hayano, A.; Takashima, Y. Secondary Intracranial Tumors Following Radiotherapy for Pituitary Adenomas: A Systematic Review. Cancers 2017, 9, 103. [Google Scholar] [CrossRef] [Green Version]
- Song, Z.J.; Reitman, Z.J.; Ma, Z.Y.; Chen, J.H.; Zhang, Q.L.; Shou, X.F.; Huang, C.X.; Wang, Y.F.; Li, S.Q.; Mao, Y.; et al. The genome-wide mutational landscape of pituitary adenomas. Cell Res. 2016, 26, 1255–1259. [Google Scholar] [CrossRef] [Green Version]
- Välimäki, N.; Schalin-Jäntti, C.; Karppinen, A.; Paetau, A.; Kivipelto, L.; Aaltonen, L.A.; Karhu, A. Genetic and epigenetic characterization of growth hormone-secreting pituitary tumors. Mol. Cancer Res. 2019, 17, 2432–2443. [Google Scholar] [CrossRef] [Green Version]
- Theodoropoulou, M.; Reincke, M.; Fassnacht, M.; Komada, M. Decoding the genetic basis of Cushing’s disease: USP8 in the spotlight. Eur. J. Endocrinol. 2015, 173, M73–M83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Li, C.; Wang, J.; Song, G.; Zhao, Z.; Wang, H.; Wang, W.; Li, H.; Li, Z.; Miao, Y.; et al. Genome-wide analysis of differentially expressed lncRNAs and mRNAs in primary gonadotrophin adenomas by RNA-seq. Oncotarget 2017, 8, 4585–4606. [Google Scholar] [CrossRef] [Green Version]
- Ubersax, J.A.; Ferrell, J.E., Jr. Mechanisms of specificity in protein phosphorylation. Nat. Rev. Mol. Cell Biol. 2007, 8, 530–541. [Google Scholar] [CrossRef] [PubMed]
- Manning, G.; Whyte, D.B.; Martinez, R.; Hunter, T.; Sudarsanam, S. The protein kinase complement of the human genome. Science 2002, 298, 1912–1934. [Google Scholar] [CrossRef] [Green Version]
- Amano, M.; Nishioka, T.; Tsuboi, D.; Kuroda, K.; Funahashi, Y.; Yamahashi, Y.; Kaibuchi, K. Comprehensive analysis of kinase-oriented phospho-signalling pathways. J. Biochem. 2019, 165, 301–307. [Google Scholar] [CrossRef]
- McAlister, G.C.; Huttlin, E.L.; Haas, W.; Ting, L.; Jedrychowski, M.P.; Rogers, J.C.; Kuhn, K.; Pike, I.; Grothe, R.A.; Blethrow, J.D.; et al. Increasing the multiplexing capacity of TMTs using reporter ion isotopologues with isobaric masses. Anal. Chem. 2012, 84, 7469–7478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Brien, D.P.; Timms, J.F. Employing TMT Quantification in a Shotgun-MS Platform. In Shotgun Proteomics: Methods and Protocols; Methods in Molecular Biology; Martins, D., Ed.; Humana Press: New York, NY, USA, 2014; Volume 1156, pp. 187–199. [Google Scholar]
- Chahrour, O.; Cobice, D.; Malone, J. Stable isotope labelling methods in mass spectrometry-based quantitative proteomics. J. Pharm. Biomed. Anal. 2015, 113, 2–20. [Google Scholar] [CrossRef]
- Lee, S.C.; Abdel-Wahab, O. Therapeutic targeting of splicing in cancer. Nat. Med. 2016, 22, 976–986. [Google Scholar] [CrossRef]
- Duden, R. ER-to-Golgi transport: COP I and COP II function (Review). Mol. Membr. Biol. 2003, 20, 197–207. [Google Scholar] [CrossRef]
- Wang, J.; Davis, S.; Zhu, M.; Miller, E.A.; Ferro-Novick, S. Autophagosome formation: Where the secretory and autophagy pathways meet. Autophagy 2017, 13, 973–974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Errington, W.J.; Bruncsics, B.; Sarkar, C.A. Mechanisms of noncanonical binding dynamics in multivalent protein-protein interactions. Proc. Natl. Acad. Sci. USA 2019, 116, 25659–25667. [Google Scholar] [CrossRef] [PubMed]
- Friedl, P.; Alexander, S. Cancer invasion and the microenvironment: Plasticity and reciprocity. Cell 2011, 147, 992–1009. [Google Scholar] [CrossRef] [Green Version]
- Anastasiadou, E.; Jacob, L.S.; Slack, F.J. Non-coding RNA networks in cancer. Nat. Rev. Cancer 2018, 18, 5–18. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Zhang, C.; Zhao, Q.; Li, D. Spectrin: Structure, function and disease. Sci. China Life Sci. 2013, 56, 1076–1085. [Google Scholar] [CrossRef] [Green Version]
- Hodges, C.; Kirkland, J.G.; Crabtree, G.R. The Many Roles of BAF (mSWI/SNF) and PBAF Complexes in Cancer. Cold Spring Harb. Perspect. Med. 2016, 6, a026930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mashtalir, N.; D’Avino, A.R.; Michel, B.C.; Luo, J.; Pan, J.; Otto, J.E.; Zullow, H.J.; McKenzie, Z.M.; Kubiak, R.L.; St Pierre, R.; et al. Modular Organization and Assembly of SWI/SNF Family Chromatin Remodeling Complexes. Cell 2018, 175, 1272–1288.e20. [Google Scholar] [CrossRef] [Green Version]
- Panwalkar, P.; Pratt, D.; Chung, C.; Dang, D.; Le, P.; Martinez, D.; Bayliss, J.M.; Smith, K.S.; Adam, M.; Potter, S.; et al. SWI/SNF complex heterogeneity is related to polyphenotypic differentiation, prognosis, and immune response in rhabdoid tumors. Neuro Oncol. 2020, 22, 785–796. [Google Scholar] [CrossRef]
- Guo, G.; Sun, X.; Chen, C.; Wu, S.; Huang, P.; Li, Z.; Dean, M.; Huang, Y.; Jia, W.; Zhou, Q.; et al. Whole-genome and whole-exome sequencing of bladder cancer identifies frequent alterations in genes involved in sister chromatid cohesion and segregation. Nat. Genet. 2013, 45, 1459–1463. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Pluhackova, K.; Böckmann, R.A. The Multifaceted Role of SNARE Proteins in Membrane Fusion. Front. Physiol. 2017, 8, 5. [Google Scholar] [CrossRef] [Green Version]
- Hedglin, M.; Benkovic, S.J. Regulation of Rad6/Rad18 Activity during DNA Damage Tolerance. Annu. Rev. Biophys. 2015, 44, 207–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alcón, P.; Shakeel, S.; Chen, Z.A.; Rappsilber, J.; Patel, K.J.; Passmore, L.A. FANCD2-FANCI is a clamp stabilized on DNA by monoubiquitination of FANCD2 during DNA repair. Nat. Struct. Mol. Biol. 2020, 27, 240–248. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J. Cell Fate Control by Translation: mRNA Translation Initiation as a Therapeutic Target for Cancer Development and Stem Cell Fate Control. Biomolecules 2019, 9, 665. [Google Scholar] [CrossRef] [Green Version]
- Sriram, A.; Bohlen, J.; Teleman, A.A. Translation acrobatics: How cancer cells exploit alternate modes of translational initiation. EMBO Rep. 2018, 19, e45947. [Google Scholar] [CrossRef]
- Stephens, A.D.; Banigan, E.J.; Marko, J.F. Chromatin’s physical properties shape the nucleus and its functions. Curr. Opin. Cell Biol. 2019, 58, 76–84. [Google Scholar] [CrossRef]
- Viphakone, N.; Sudbery, I.; Griffith, L.; Heath, C.G.; Sims, D.; Wilson, S.A. Co-transcriptional Loading of RNA Export Factors Shapes the Human Transcriptome. Mol. Cell 2019, 75, 310–323.e8. [Google Scholar] [CrossRef] [Green Version]
- Theocharis, A.D.; Skandalis, S.S.; Tzanakakis, G.N.; Karamanos, N.K. Proteoglycans in health and disease: Novel roles for proteoglycans in malignancy and their pharmacological targeting. FEBS J. 2010, 277, 3904–3923. [Google Scholar] [CrossRef]
- Morath, I.; Hartmann, T.N.; Orian-Rousseau, V. CD44: More than a mere stem cell marker. Int. J. Biochem. Cell Biol. 2016, 81, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.A.; Scheller, R.H. SNARE-mediated membrane fusion. Nat. Rev. Mol. Cell Biol. 2001, 2, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, G.H. The secretion, components, and properties of saliva. Annu. Rev. Food Sci. Technol. 2013, 4, 267–276. [Google Scholar] [CrossRef]
- Li, Z.; Delaney, M.K.; O’Brien, K.A.; Du, X. Signaling during platelet adhesion and activation. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 2341–2349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho-Tin-Noé, B.; Boulaftali, Y.; Camerer, E. Platelets and vascular integrity: How platelets prevent bleeding in inflammation. Blood 2018, 131, 277–288. [Google Scholar] [CrossRef]
- Veiga, E.; Guttman, J.A.; Bonazzi, M.; Boucrot, E.; Toledo-Arana, A.; Lin, A.E.; Enninga, J.; Pizarro-Cerdá, J.; Finlay, B.B.; Kirchhausen, T.; et al. Invasive and adherent bacterial pathogens co-Opt host clathrin for infection. Cell Host Microbe 2007, 2, 340–351. [Google Scholar] [CrossRef] [Green Version]
- Cossart, P.; Veiga, E. Non-classical use of clathrin during bacterial infections. J. Microsc. 2008, 231, 524–528. [Google Scholar] [CrossRef]
- Takano, K.; Kojima, T.; Sawada, N.; Himi, T. Role of tight junctions in signal transduction: An update. EXCLI J. 2014, 13, 1145–1162. [Google Scholar]
- González-Mariscal, L.; Domínguez-Calderón, A.; Raya-Sandino, A.; Ortega-Olvera, J.M.; Vargas-Sierra, O.; Martínez-Revollar, G. Tight junctions and the regulation of gene expression. Semin. Cell Dev. Biol. 2014, 36, 213–223. [Google Scholar] [CrossRef]
- Wier, W.G.; Mauban, J.R.H. Imaging sympathetic neurogenic Ca2+ signaling in blood vessels. Auton. Neurosci. 2017, 207, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Csordás, G.; Weaver, D.; Hajnóczky, G. Endoplasmic Reticulum-Mitochondrial Contactology: Structure and Signaling Functions. Trends Cell Biol. 2018, 28, 523–540. [Google Scholar] [CrossRef]
- Yu, S.; Sun, L.; Jiao, Y.; Lee, L.T.O. The Role of G Protein-coupled Receptor Kinases in Cancer. Int. J. Biol. Sci. 2018, 14, 189–203. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.; Zhu, Y.; Xu, J.; Yang, M.; Chen, P.; Xu, W.; Zhao, J.; Geng, L.; Gong, S. PKN2 in colon cancer cells inhibits M2 phenotype polarization of tumor-associated macrophages via regulating DUSP6-Erk1/2 pathway. Mol. Cancer 2018, 17, 13. [Google Scholar] [CrossRef]
- Carling, D. AMPK signalling in health and disease. Curr. Opin. Cell Biol. 2017, 45, 31–37. [Google Scholar] [CrossRef]
- Herzig, S.; Shaw, R.J. AMPK: Guardian of metabolism and mitochondrial homeostasis. Nat. Rev. Mol. Cell Biol. 2018, 19, 121–135. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Yang, R.; Gu, J.; Yin, X.; Jin, N.; Xie, S.; Wang, Y.; Chang, H.; Qian, W.; Shi, J.; et al. Cross talk between PI3K-AKT-GSK-3beta and PP2A pathways determines tau hyperphosphorylation. Neurobiol. Aging 2015, 36, 188–200. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Lu, X.X.; Wang, J.R.; Yang, T.Y.; Li, X.M.; He, X.S.; Li, Y.; Ye, W.L.; Wu, Y.; Gan, W.J.; et al. TRAF6 inhibits colorectal cancer metastasis through regulating selective autophagic CTNNB1/β-catenin degradation and is targeted for GSK3B/GSK3β-mediated phosphorylation and degradation. Autophagy 2019, 15, 1506–1522. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, I.; Hoessli, D.C.; Walker-Nasir, E.; Choudhary, M.I.; Rafik, S.M.; Shakoori, A.R. Phosphorylation and glycosylation interplay: Protein modifications at hydroxy amino acids and prediction of signaling functions of the human β3 integrin family. J. Cell. Biochem. 2006, 99, 706–718. [Google Scholar] [CrossRef] [PubMed]
- Giorgianni, F.; Beranova-Giorgianni, S.; Desiderio, D.M. Identification and characterization of phosphorylated proteins in the human pituitary. Proteomics 2004, 4, 587–598. [Google Scholar] [CrossRef] [PubMed]
- Beranova-Giorgianni, S.; Zhao, Y.; Desiderio, D.M.; Giorgianni, F. Phosphoproteomic analysis of the human pituitary. Pituitary 2006, 9, 109–120. [Google Scholar] [CrossRef]
- Long, Y.; Lu, M.; Cheng, T.; Zhan, X.; Zhan, X. Multiomics-Based Signaling Pathway Network Alterations in Human Non-functional Pituitary Adenomas. Front. Endocrinol. 2019, 10, 835. [Google Scholar] [CrossRef]
- Peverelli, E.; Giardino, E.; Mangili, F.; Treppiedi, D.; Catalano, R.; Ferrante, E.; Sala, E.; Locatelli, M.; Lania, A.G.; Arosio, M.; et al. cAMP/PKA-induced filamin A (FLNA) phosphorylation inhibits SST2 signal transduction in GH-secreting pituitary tumor cells. Cancer Lett. 2018, 435, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y. Mechanistic insights into precursor messenger RNA splicing by the spliceosome. Nat. Rev. Mol. Cell Biol. 2017, 18, 655–670. [Google Scholar] [CrossRef] [PubMed]
- Sotillo, E.; Barrett, D.M.; Black, K.L.; Bagashev, A.; Oldridge, D.; Wu, G.; Sussman, R.; Lanauze, C.; Ruella, M.; Gazzara, M.R.; et al. Convergence of Acquired Mutations and Alternative Splicing of CD19 Enables Resistance to CART-19 Immunotherapy. Cancer Discov. 2015, 5, 1282–1295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vázquez-Borrego, M.C.; Fuentes-Fayos, A.C.; Venegas-Moreno, E.; Rivero-Cortés, E.; Dios, E.; Moreno-Moreno, P.; Madrazo-Atutxa, A.; Remón, P.; Solivera, J.; Wildemberg, L.E.; et al. Splicing machinery is dysregulated in pituitary neuroendocrine tumors and is associated with aggressiveness features. Cancers 2019, 11, 1439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, A.; Sipma, W.; Veenhoff, L.M.; Van der Giessen, E.; Onck, P.R. The Effect of FG-Nup Phosphorylation on NPC Selectivity: A One-Bead-Per-Amino-Acid Molecular Dynamics Study. Int. J. Mol. Sci. 2019, 20, 596. [Google Scholar] [CrossRef] [Green Version]
- Buffone, A.; Weaver, V.M. Don’t sugarcoat it: How glycocalyx composition influences cancer progression. J. Cell Biol. 2020, 219, e201910070. [Google Scholar] [CrossRef]
- Tarbell, J.M.; Cancel, L.M. The glycocalyx and its significance in human medicine. J. Intern. Med. 2016, 280, 97–113. [Google Scholar] [CrossRef] [Green Version]
- Kang, H.; Wu, Q.; Sun, A.; Liu, X.; Fan, Y.; Deng, X. Cancer Cell Glycocalyx and Its Significance in Cancer Progression. Int. J. Mol. Sci. 2018, 19, 2484. [Google Scholar] [CrossRef] [Green Version]
- Kavalali, E.T. SNARE interactions in membrane trafficking: A perspective from mammalian central synapses. Bioessays 2002, 24, 926–936. [Google Scholar] [CrossRef] [PubMed]
- Risinger, C.; Bennett, M.K. Differential phosphorylation of syntaxin and synaptosome-associated protein of 25 kDa (SNAP-25) isoforms. J. Neurochem. 1999, 72, 614–624. [Google Scholar] [CrossRef] [PubMed]
- Otani, T.; Furuse, M. Tight Junction Structure and Function Revisited. Trends Cell Biol. 2020, 30, 805–817. [Google Scholar] [CrossRef] [PubMed]
- Anbalagan, S.; Gordon, L.; Blechman, J.; Matsuoka, R.L.; Rajamannar, P.; Wircer, E.; Biran, J.; Reuveny, A.; Leshkowitz, D.; Stainier, D.Y.R.; et al. Pituicyte Cues Regulate the Development of Permeable Neuro-Vascular Interfaces. Dev. Cell 2018, 47, 711–726.e5. [Google Scholar] [CrossRef] [Green Version]
- Hashimoto, Y.; Tachibana, K.; Krug, S.M.; Kunisawa, J.; Fromm, M.; Kondoh, M. Potential for Tight Junction Protein-Directed Drug Development Using Claudin Binders and Angubindin-1. Int. J. Mol. Sci. 2019, 20, 4016. [Google Scholar] [CrossRef] [Green Version]
- Spadaro, D.; Le, S.; Laroche, T.; Mean, I.; Jond, L.; Yan, J.; Citi, S. Tension-Dependent Stretching Activates ZO-1 to Control the Junctional Localization of Its Interactors. Curr. Biol. 2017, 27, 3783–3795.e8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolinger, M.T.; Ramshekar, A.; Waldschmidt, H.V.; Larsen, S.D.; Bewley, M.C.; Flanagan, J.M.; Antonetti, D.A. Occludin S471 Phosphorylation Contributes to Epithelial Monolayer Maturation. Mol. Cell. Biol. 2016, 36, 2051–2066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhullar, K.S.; Lagarón, N.O.; McGowan, E.M.; Parmar, I.; Jha, A.; Hubbard, B.P.; Rupasinghe, H.P.V. Kinase-targeted cancer therapies: Progress, challenges and future directions. Mol. Cancer 2018, 17, 48. [Google Scholar] [CrossRef]
- Yan, Y.; Zhou, X.E.; Xu, H.E.; Melcher, K. Structure and Physiological Regulation of AMPK. Int. J. Mol. Sci. 2018, 19, 3534. [Google Scholar] [CrossRef] [Green Version]
- Singh, V.; Ram, M.; Kumar, R.; Prasad, R.; Roy, B.K.; Singh, K.K. Phosphorylation: Implications in Cancer. Protein J. 2017, 36, 1–6. [Google Scholar] [CrossRef]
- Cicenas, J.; Cicenas, E. Multi-kinase inhibitors, AURKs and cancer. Med. Oncol. 2016, 33, 43. [Google Scholar] [CrossRef]
- Xiao, A.; Li, H.; Shechter, D.; Ahn, S.H.; Fabrizio, L.A.; Erdjument-Bromage, H.; Ishibe-Murakami, S.; Wang, B.; Tempst, P.; Hofmann, K.; et al. WSTF regulates the H2A.X DNA damage response via a novel tyrosine kinase activity. Nature 2009, 457, 57–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turinetto, V.; Giachino, C. Multiple facets of histone variant H2AX: A DNA double-strand-break marker with several biological functions. Nucleic Acids Res. 2015, 43, 2489–2498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okazaki, Y.; Ohno, H.; Takase, K.; Ochiai, T.; Saito, T. Cell surface expression of calnexin, a molecular chaperone in the endoplasmic reticulum. J. Biol. Chem. 2000, 275, 35751–35758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Ma, D.; Wang, X.; Fang, J.; Liu, X.; Song, J.; Li, X.; Ren, X.; Li, Q.; Li, Q.; et al. Calnexin Impairs the Antitumor Immunity of CD4+ and CD8+ T Cells. Cancer Immunol. Res. 2019, 7, 123–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishioka, H.; Inoshita, N. New WHO classification of pituitary adenomas (4th edition): Assessment of pituitary transcription factors and the prognostic histological factors. Brain Tumor Pathol. 2018, 35, 57–61. [Google Scholar] [CrossRef] [PubMed]
- Mete, O.; Lopes, M.B. Overview of the 2017 WHO classification of pituitary tumors. Endocr. Pathol. 2017, 28, 228–243. [Google Scholar] [CrossRef] [PubMed]
- Saeger, W.; Wilczak, P.; Luüdecke, D.K.; Buchfelder, M.; Fahlbusch, R. Hormone markers in pituitary adenomas: Changes within last decade resulting from improved method. Endocr. Pathol. 2003, 14, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Saeger, W.; Lüdecke, D.K.; Buchfelder, M.; Fahlbusch, R.; Quabbe, H.J.; Petersenn, S. Pathohistological classification of pituitary tumors: 10 years of experience with the German Pituitary Tumor Registry. Eur. J. Endocrinol. 2007, 156, 203–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitrofanova, L.B.; Konovalov, P.V.; Krylova, J.S.; Polyakova, V.O.; Kvetnoy, I.M. Plurihormonal cells of normal anterior pituitary: Facts and conclusions. Oncotarget 2017, 8, 29282–29299. [Google Scholar] [CrossRef] [Green Version]
- Zhan, X.; Desiderio, D.M. Heterogeneity analysis of the human pituitaryproteome. Clin. Chem. 2003, 49, 1740–1751. [Google Scholar] [CrossRef] [Green Version]
- Moreno, C.S.; Evans, C.O.; Zhan, X.; Okor, M.; Desiderio, D.M.; Oyesiku, N.M. Novel molecular signaling and classification of human clinically nonfunctional pituitary adenomas identified by gene expression profiling and proteomic analyses. Cancer Res. 2005, 65, 10214–10222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhan, X.; Wang, X.; Long, Y.; Desiderio, D.M. Heterogeneity analysis of the proteomes in clinically nonfunctional pituitary adenomas. BMC Med. Genet. 2014, 7, 69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhan, X.; Long, Y.; Zhan, X.; Mu, Y. Consideration of statistical vs. biological significances for omics data-based pathway network analysis. Med. One 2017, 1, e170002. [Google Scholar] [CrossRef]
- Christ-Crain, M. Vasopressin and Copeptin in health and disease. Rev. Endocr. Metab. Disord. 2019, 20, 283–294. [Google Scholar] [CrossRef] [PubMed]
- Klussmann, E.; Maric, K.; Rosenthal, W. The mechanisms of aquaporin control in the renal collecting duct. Rev. Physiol. Biochem. Pharmacol. 2000, 141, 33–95. [Google Scholar] [CrossRef]
- Proctor, G.B. The physiology of salivary secretion. Periodontol. 2000 2016, 70, 11–25. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | ID | Count | % | p-Value | Genes | |
|---|---|---|---|---|---|---|
| Annotation Cluster 1 Enrichment Score: 12.9 | ||||||
| GOTERM_CC_DIRECT | GO:0005913 | cell–cell adherens junction | 42 | 7.4 | 5.37 × 10−15 | Q9UHB6, Q9UPN3, P18206, Q9H0B6, A0A087WUZ3, A0A0U4BW16, Q9UDY2, P35221, A0A024R1S8, Q9C0C2, Q15762, P55196, Q6PKG0, Q09666, O60716, C9J6P4, P42166, A0A024R4E5, P07948, E9PRY8, Q9H4G0, Q9ULH1, P35611, Q9H2G2, P21333, Q07960, O00567, Q92522, P35579, Q13813, Q15149, O60763, P08238, O95292, Q9UQN3, Q9BY44, A0A024RAN2, Q16513, O76021, E7EX44, Q92597, P26232, Q14247 |
| GOTERM_MF_DIRECT | GO:0098641 | cadherin binding involved in cell–cell adhesion | 40 | 7.0 | 2.04 × 10−14 | Q9UHB6, Q9UPN3, P18206, Q9H0B6, A0A087WUZ3, Q9UDY2, P35221, A0A0U4BW16, A0A024R1S8, Q9C0C2, P55196, Q6PKG0, Q09666, O60716, C9J6P4, P42166, A0A024R4E5, E9PRY8, Q9H4G0, Q9ULH1, P35611, Q9H2G2, P21333, Q07960, O00567, Q92522, P35579, Q13813, Q15149, O60763, P08238, O95292, Q9UQN3, Q9BY44, A0A024RAN2, Q16513, O76021, E7EX44, Q92597, P26232, Q14247 |
| GOTERM_BP_DIRECT | GO:0098609 | cell–cell adhesion | 34 | 6.0 | 2.25 × 10−11 | Q9UHB6, Q9UPN3, Q9H0B6, A0A087WUZ3, Q9UDY2, A0A024R1S8, Q9C0C2, P55196, Q6PKG0, Q09666, C9J6P4, P42166, A0A024R4E5, E9PRY8, Q9H4G0, Q9ULH1, P35611, Q9H2G2, Q07960, O00567, Q92522, Q13813, Q15149, O60763, P08238, O95292, Q9BY44, Q9UQN3, A0A024RAN2, Q16513, O76021, E7EX44, Q92597, Q14247 |
| Annotation Cluster 2 Enrichment Score: 7.9 | ||||||
| GOTERM_BP_DIRECT | GO:0006405 | RNA export from nucleus | 15 | 2.6 | 9.39 × 10−10 | Q15287, Q13247, O95391, Q96FV9, P52948, A0A0S2Z4Z6, J3KTL2, Q08170, Q16629, O75694, Q05519, Q13242, P35658, P09651, Q01130 |
| GOTERM_BP_DIRECT | GO:0006406 | mRNA export from nucleus | 19 | 3.3 | 1.69 × 10−9 | Q15287, Q9BRD0, Q13247, O75494, O95391, L0R530, Q96FV9, P52948, A0A0S2Z4Z6, J3KTL2, Q08170, P49792, Q16629, O75694, Q05519, Q13242, Q9P2I0, P35658, Q01130 |
| GOTERM_BP_DIRECT | GO:0031124 | mRNA 3’-end processing | 13 | 2.3 | 3.10 × 10−8 | Q15287, Q08170, Q13247, Q16629, Q12996, O95391, Q05519, Q13242, Q96FV9, Q9P2I0, A0A0S2Z4Z6, Q01130, J3KTL2 |
| GOTERM_BP_DIRECT | GO:0006369 | termination of RNA polymerase II transcription | 13 | 2.3 | 5.63 × 10−7 | Q15287, Q08170, Q13247, Q16629, Q12996, O95391, Q05519, Q13242, Q96FV9, Q9P2I0, A0A0S2Z4Z6, Q01130, J3KTL2 |
| Annotation Cluster 3 Enrichment Score: 4.0 | ||||||
| GOTERM_CC_DIRECT | GO:0014731 | spectrin-associated cytoskeleton | 6 | 1.1 | 1.24 × 10−6 | Q12955, Q08495, P16157, A0A087WUZ3, P11171, P11277 |
| GOTERM_CC_DIRECT | GO:0008091 | spectrin | 5 | 0.9 | 8.97 × 10−5 | A0A087WUZ3, P11171, P11277, O43491, Q13813 |
| GOTERM_BP_DIRECT | GO:0051693 | actin filament capping | 4 | 0.7 | 6.84 × 10−3 | Q08495, A0A087WUZ3, P11277, Q13813 |
| Annotation Cluster 4 Enrichment Score: 3.0 | ||||||
| GOTERM_BP_DIRECT | GO:0043044 | ATP-dependent chromatin remodeling | 7 | 1.2 | 5.76 × 10−5 | Q13547, P07910, Q92769, Q14839, B4DY08, Q92922, O96019, F8VXC8 |
| GOTERM_MF_DIRECT | GO:0031492 | nucleosomal DNA binding | 9 | 1.6 | 8.71 × 10−5 | Q13547, P05114, P07910, Q92769, Q14839, B4DY08, Q92922, P49450, O96019, F8VXC8 |
| GOTERM_CC_DIRECT | GO:0000790 | nuclear chromatin | 13 | 2.3 | 1.39 × 10−2 | P51531, Q9H1E3, Q9Y618, P52701, Q92769, O75376, Q14839, O96019, F8VXC8, Q13547, P07910, P16402, B4DY08, Q92922 |
| GOTERM_MF_DIRECT | GO:0000980 | RNA polymerase II distal enhancer sequence-specific DNA binding | 7 | 1.2 | 1.68 × 10−2 | Q13547, P07910, Q92769, Q14839, B4DY08, Q92922, O96019, F8VXC8 |
| Annotation Cluster 5 Enrichment Score: 2.9 | ||||||
| GOTERM_CC_DIRECT | GO:0071564 | npBAF complex | 5 | 0.9 | 3.28 × 10−4 | P51531, Q8WUB8, Q92922, O96019, F8VXC8 |
| GOTERM_CC_DIRECT | GO:0016514 | SWI/SNF complex | 5 | 0.9 | 8.42 × 10−4 | P51531, Q92922, Q8NFD5, O96019, F8VXC8 |
| GOTERM_CC_DIRECT | GO:0071565 | nBAF complex | 4 | 0.7 | 7.65 × 10−3 | P51531, Q92922, Q8NFD5, F8VXC8 |
| Annotation Cluster 6 Enrichment Score: 2.4 | ||||||
| GOTERM_BP_DIRECT | GO:0007064 | mitotic sister chromatid cohesion | 5 | 0.9 | 7.31 × 10−4 | Q9NTI5, Q7Z5K2, Q29RF7, Q6KC79, Q9UQE7 |
| GOTERM_CC_DIRECT | GO:0000775 | chromosome, centromeric region | 7 | 1.2 | 7.06 × 10−3 | Q9NTI5, P83916, Q13185, Q7Z5K2, Q29RF7, P49450, Q9UQE7 |
| GOTERM_BP_DIRECT | GO:0007062 | sister chromatid cohesion | 9 | 1.6 | 1.53 × 10−2 | Q9NTI5, P49792, O75122, Q7Z5K2, Q29RF7, P49450, Q9UQE7, Q8WYP5, P52948 |
| Annotation Cluster 7 Enrichment Score: 2.3 | ||||||
| GOTERM_BP_DIRECT | GO:0061025 | membrane fusion | 9 | 1.6 | 5.61 × 10−5 | O00161, D3DUW5, P63027, Q05193, Q16623, O60763, Q9UNZ2, Q9UQ16, P61266 |
| KEGG_PATHWAY | hsa04130:S | NARE interactions in vesicular transport | 6 | 1.1 | 4.25 × 10−3 | O00161, P63027, Q16623, O75396, P61266, O75379 |
| GOTERM_MF_DIRECT | GO:0005484 | SNAP receptor activity | 6 | 1.1 | 7.42 × 10−3 | O00161, P63027, Q16623, O75396, P61266, O75379 |
| GOTERM_BP_DIRECT | GO:0016192 | vesicle-mediated transport | 12 | 2.1 | 8.19 × 10−3 | P63027, Q16623, O75396, Q13439, O00203, O75131, P61266, P35606, O75379, Q13367, Q9UPT6, Q9UN37 |
| GOTERM_CC_DIRECT | GO:0031201 | SNARE complex | 6 | 1.1 | 2.10 × 10−2 | O00161, P63027, Q16623, O75396, P61266, O75379 |
| GOTERM_BP_DIRECT | GO:0017157 | regulation of exocytosis | 4 | 0.7 | 4.62 × 10−2 | P63027, Q16623, P61266, Q9Y6V0 |
| Annotation Cluster 8 Enrichment Score: 2.3 | ||||||
| GOTERM_BP_DIRECT | GO:0016925 | protein sumoylation | 14 | 2.5 | 7.34 × 10−5 | Q02880, Q12888, Q99502, A0A024R2M8, Q14676, L0R530, Q8NDX5, P52948, P07910, P49792, O75694, B4DY08, Q9UQE7, P35658, P29590 |
| GOTERM_BP_DIRECT | GO:1900034 | regulation of cellular response to heat | 9 | 1.6 | 2.30 × 10−3 | Q96B36, P07900, P08238, P49792, O75694, B3KUY2, L0R530, P35658, P52948 |
| GOTERM_BP_DIRECT | GO:0007077 | mitotic nuclear envelope disassembly | 7 | 1.2 | 2.31 × 10−3 | P02545, P49792, O75694, P17252, L0R530, P35658, P52948 |
| GOTERM_BP_DIRECT | GO:0006409 | tRNA export from nucleus | 5 | 0.9 | 1.69 × 10−2 | P49792, O75694, L0R530, P35658, P52948 |
| GOTERM_BP_DIRECT | GO:0010827 | regulation of glucose transport | 5 | 0.9 | 1.87 × 10−2 | P49792, O75694, L0R530, P35658, P52948 |
| GOTERM_BP_DIRECT | GO:0075733 | intracellular transport of virus | 6 | 1.1 | 2.10 × 10−2 | P49792, O75694, L0R530, P35658, O00505, P52948 |
| GOTERM_CC_DIRECT | GO:0044615 | nuclear pore nuclear basket | 3 | 0.5 | 4.86 × 10−2 | P49792, P35658, P52948 |
| Annotation Cluster 9 Enrichment Score: 2.2 | ||||||
| GOTERM_BP_DIRECT | GO:0031032 | actomyosin structure organization | 7 | 1.2 | 1.52 × 10−4 | P35580, Q9H4G0, Q9Y2J2, Q92614, A0A0U4BW16, P11171, P35579, O43491 |
| GOTERM_CC_DIRECT | GO:0019898 | extrinsic component of membrane | 7 | 1.2 | 3.67 × 10−2 | Q9UEW8, Q9H4G0, Q9Y2J2, Q96C24, P11171, Q9Y4F1, O43491 |
| GOTERM_BP_DIRECT | GO:0030866 | cortical actin cytoskeleton organization | 4 | 0.7 | 3.76 × 10−2 | Q9H4G0, Q9Y2J2, P11171, O43491 |
| Annotation Cluster 10 Enrichment Score: 2.1 | ||||||
| GOTERM_BP_DIRECT | GO:0033523 | histone H2B ubiquitination | 4 | 0.7 | 1.50 × 10−3 | Q5VTR2, Q6PD62, Q8WVC0, Q8N7H5 |
| GOTERM_BP_DIRECT | GO:0010390 | histone monoubiquitination | 4 | 0.7 | 4.13 × 10−3 | Q5VTR2, Q6PD62, Q8WVC0, Q8N7H5 |
| GOTERM_BP_DIRECT | GO:0001711 | endodermal cell fate commitment | 3 | 0.5 | 1.34 × 10−2 | Q6PD62, Q8WVC0, Q8N7H5 |
| GOTERM_CC_DIRECT | GO:0016593 | Cdc73/Paf1 complex | 3 | 0.5 | 1.71 × 10−2 | Q6PD62, Q8WVC0, Q8N7H5 |
| GOTERM_BP_DIRECT | GO:0045638 | negative regulation of myeloid cell differentiation | 4 | 0.7 | 2.02 × 10−2 | Q96T37, Q6PD62, Q8WVC0, Q8N7H5 |
| Annotation Cluster 11 Enrichment Score: 1.5 | ||||||
| GOTERM_BP_DIRECT | GO:0006446 | regulation of translational initiation | 5 | 0.9 | 2.51 × 10−2 | B5ME19, O60841, E7EX17, Q59GJ0, P04792, P23588 |
| GOTERM_BP_DIRECT | GO:0006413 | translational initiation | 10 | 1.8 | 2.79 × 10−2 | Q8NE71, P05387, Q13144, B5ME19, Q6PKG0, Q9BY44, O60841, P05388, E7EX17, Q59GJ0, P23588 |
| GOTERM_MF_DIRECT | GO:0003743 | translation initiation factor activity | 6 | 1.1 | 4.37 × 10−2 | Q13144, B5ME19, Q9BY44, O60841, E7EX17, Q59GJ0, P23588 |
| Annotation Cluster 12 Enrichment Score: 1.4 | ||||||
| GOTERM_BP_DIRECT | GO:1904903 | ESCRT III complex disassembly | 3 | 0.5 | 3.70 × 10−2 | A0A024R2C5, Q9UQN3, Q9UN37 |
| GOTERM_BP_DIRECT | GO:1902188 | positive regulation of viral release from host cell | 3 | 0.5 | 4.44 × 10−2 | A0A024R2C5, Q9UQN3, Q9UN37 |
| GOTERM_BP_DIRECT | GO:0006997 | nucleus organization | 4 | 0.7 | 4.62 × 10−2 | A0A024R2C5, Q9UQN3, Q14980, Q9UN37 |
| Accession | KINASE | GENE | SUB | Description | Coverage | Proteins | Unique Peptides |
|---|---|---|---|---|---|---|---|
| P11021 | GRP78 | HSPA5 | GRP78 | 78 kDa glucose-regulated protein OS = Homo sapiens GN = HSPA5 PE = 1 SV = 2 [GRP78_HUMAN] | 6.57 | 12 | 2 |
| Q9UIG0 | WSTF | BAZ1B | H2AX | Tyrosine-protein kinase BAZ1B OS = Homo sapiens GN = BAZ1B PE = 1 SV = 2 [BAZ1B_HUMAN] | 1.15 | 1 | 1 |
| Q16513 | PKN2 | PKN2 | pyrin | Serine/threonine-protein kinase N2 OS = Homo sapiens GN = PKN2 PE = 1 SV = 1 [PKN2_HUMAN] | 1.42 | 1 | 1 |
| Q13523 | PRP4 | PRPF4B | ELK1 | Serine/threonine-protein kinase PRP4 homolog OS = Homo sapiens GN = PRPF4B PE = 1 SV = 3 [PRP4B_HUMAN] | 5.46 | 2 | 1 |
| O94804 | LOK | STK10 | Radixin, Ezrin, PLK1, Moesin | Serine/threonine-protein kinase 10 OS = Homo sapiens GN = STK10 PE = 1 SV = 1 [STK10_HUMAN] | 1.55 | 1 | 1 |
| Q96PY6 | NEK1 | NEK1 | TAZ, VDAC1, VHL, RAD54L | Serine/threonine-protein kinase Nek1 OS = Homo sapiens GN = NEK1 PE = 1 SV = 2 [NEK1_HUMAN] | 1.27 | 1 | 1 |
| Q13131 | AMPKA1 | PRKAA1 | 5’-AMP-activated protein kinase catalytic subunit alpha-1 OS = Homo sapiens GN = PRKAA1 PE = 1 SV = 4 [AAPK1_HUMAN] | 1.79 | 1 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Wen, S.; Li, B.; Li, N.; Zhan, X. Phosphorylation-Mediated Molecular Pathway Changes in Human Pituitary Neuroendocrine Tumors Identified by Quantitative Phosphoproteomics. Cells 2021, 10, 2225. https://doi.org/10.3390/cells10092225
Li J, Wen S, Li B, Li N, Zhan X. Phosphorylation-Mediated Molecular Pathway Changes in Human Pituitary Neuroendocrine Tumors Identified by Quantitative Phosphoproteomics. Cells. 2021; 10(9):2225. https://doi.org/10.3390/cells10092225
Chicago/Turabian StyleLi, Jiajia, Siqi Wen, Biao Li, Na Li, and Xianquan Zhan. 2021. "Phosphorylation-Mediated Molecular Pathway Changes in Human Pituitary Neuroendocrine Tumors Identified by Quantitative Phosphoproteomics" Cells 10, no. 9: 2225. https://doi.org/10.3390/cells10092225
APA StyleLi, J., Wen, S., Li, B., Li, N., & Zhan, X. (2021). Phosphorylation-Mediated Molecular Pathway Changes in Human Pituitary Neuroendocrine Tumors Identified by Quantitative Phosphoproteomics. Cells, 10(9), 2225. https://doi.org/10.3390/cells10092225