
Exosome-Mediated Immunosuppression in Tumor Microenvironments

 and
and
Abstract
:1. Introduction
2. Life Course of Exosomes
2.1. Formation of Exosomes
2.2. Release of Exosomes
2.3. Interaction between Exosomes and Target Cells
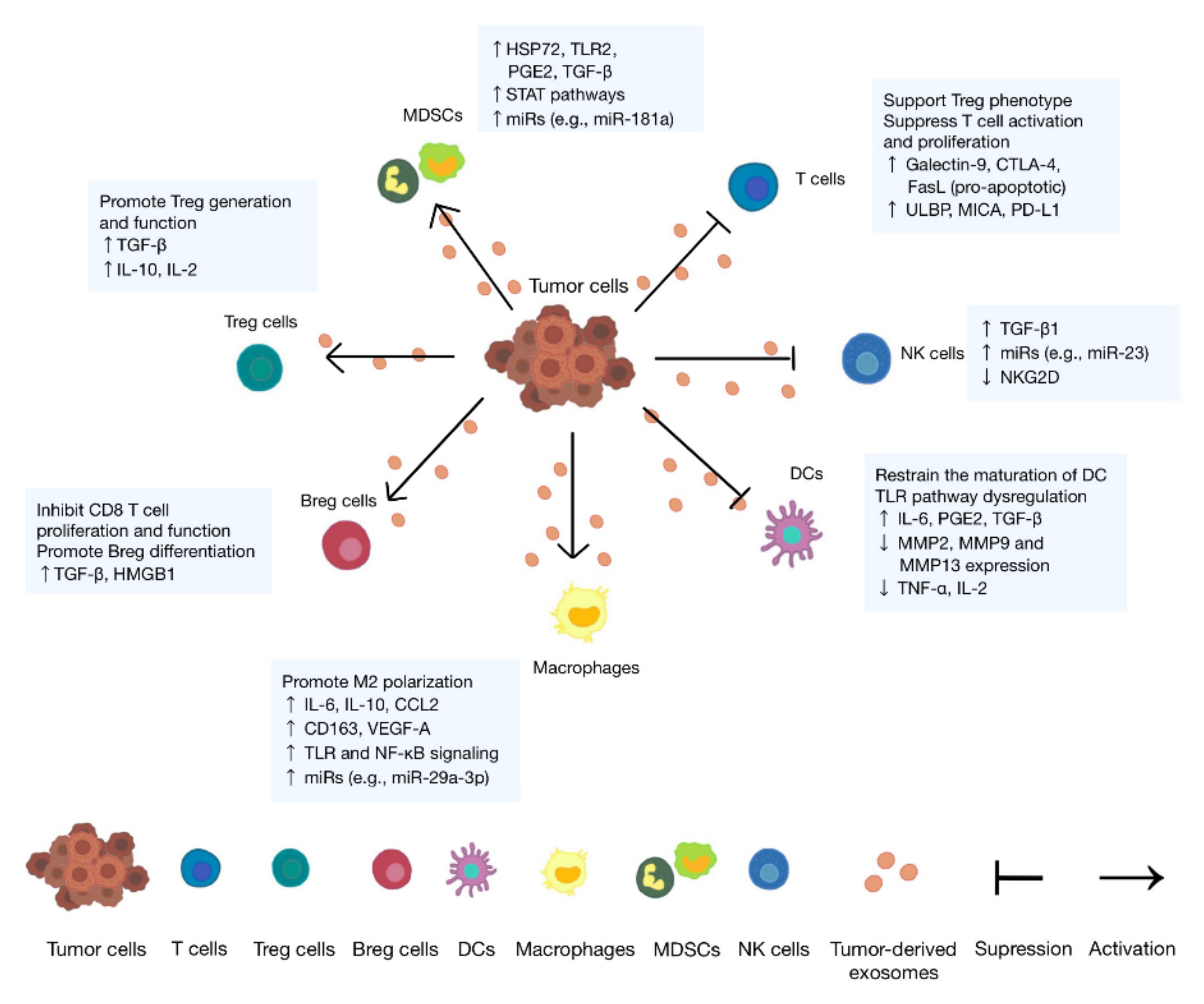
3. Immunosuppressive Role of Tumor Cell-Derived Exosomes
3.1. Lymphocytes
3.2. Macrophages
3.3. Dendritic Cells
3.4. Natural Killer Cells
3.5. Myeloid-Derived Suppressor Cells
4. Immunosuppressive Role of Exosomes Derived from Nontumor Cells
4.1. Cancer-Associated Fibroblast-Derived Exosomes
4.2. Treg-Derived Exosomes
4.3. Myeloid-Derived Suppressor Cell-Derived Exosomes
4.4. Tumor-Associated Macrophage-Derived Exosomes
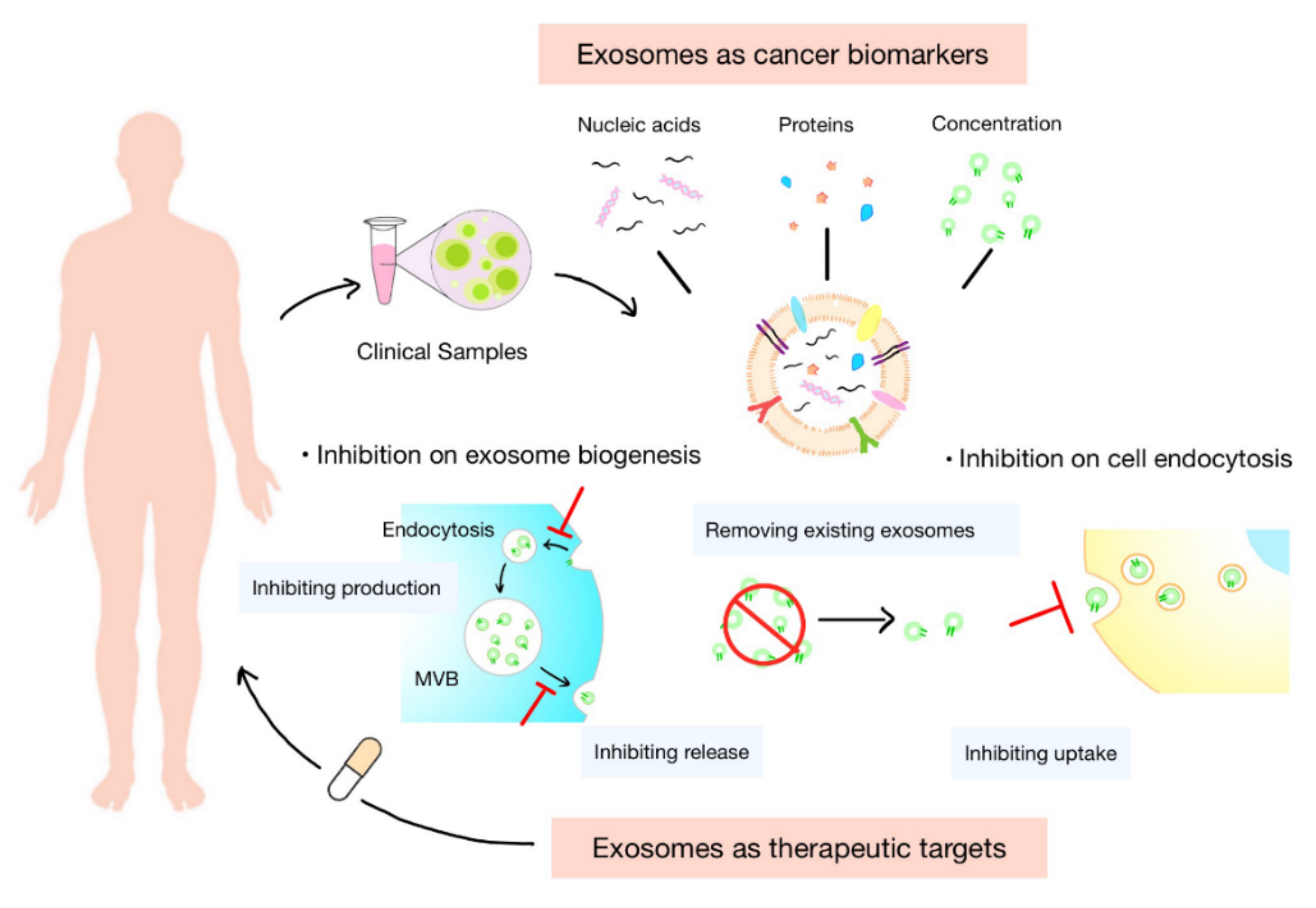
5. Exosomes as Cancer Biomarkers
5.1. Concentration of Exosomes
5.2. Nucleic Acids in Exosomes
5.3. Proteins in Exosomes
5.4. Isolation and Identification of Exosomes
6. Exosomes as Therapeutic Targets
6.1. Inhibition of Biogenesis of Exosomes
6.2. Inhibition of Endocytosis of Exosomes by Recipient Cells
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Van Niel, G.; D’Angelo, G. Shedding light on the cell biology of extracellular vesicles. Nat. Rev. Mol. Cell Biol. 2018, 19, 213–228. [Google Scholar] [CrossRef] [PubMed]
- Théry, C.; Witwer, W.K. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sung, B.H.; Parent, C.A. Extracellular vesicles: Critical players during cell migration. Dev. Cell 2021, 56, 1861–1874. [Google Scholar] [CrossRef]
- Mashouri, L.; Yousefi, H. Exosomes: Composition, biogenesis, and mechanisms in cancer metastasis and drug resistance. Mol. Cancer 2019, 18, 75. [Google Scholar] [CrossRef] [PubMed]
- Yang, E.; Wang, X. Exosome-mediated metabolic reprogramming: The emerging role in tumor microenvironment remodeling and its influence on cancer progression. Signal Transduct. Target. Ther. 2020, 5, 242. [Google Scholar] [CrossRef] [PubMed]
- Kalluri, R. The biology and function of exosomes in cancer. J. Clin. Investig. 2016, 126, 1208–1215. [Google Scholar] [CrossRef]
- Zhou, X.; Xie, F. The function and clinical application of extracellular vesicles in innate immune regulation. Cell. Mol. Immunol. 2020, 17, 323–334. [Google Scholar] [CrossRef]
- Alonso, R.; Rodríguez, M.C. Diacylglycerol kinase alpha regulates the secretion of lethal exosomes bearing Fas ligand during activation-induced cell death of T lymphocytes. J. Biol. Chem. 2005, 280, 28439–28450. [Google Scholar] [CrossRef] [Green Version]
- Laulagnier, K.; Grand, D. PLD2 is enriched on exosomes and its activity is correlated to the release of exosomes. FEBS Lett. 2004, 572, 11–14. [Google Scholar] [CrossRef] [Green Version]
- D’Souza-Schorey, C.; Schorey, J.S. Regulation and mechanisms of extracellular vesicle biogenesis and secretion. Essays Biochem. 2018, 62, 125–133. [Google Scholar]
- Mathieu, M.; Martin-Jaular, L. Specificities of secretion and uptake of exosomes and other extracellular vesicles for cell-to-cell communication. Nat. Cell Biol. 2019, 21, 9–17. [Google Scholar] [CrossRef] [PubMed]
- De Souza, A.P.; Bonorino, C. Tumor immunosuppressive environment: Effects on tumor-specific and nontumor antigen immune responses. Expert Rev. Anticancer Ther. 2009, 9, 1317–1332. [Google Scholar] [CrossRef] [PubMed]
- Wan, M.; Ning, B. Tumor-derived exosomes (TDEs): How to avoid the sting in the tail. Med. Res. Rev. 2020, 40, 385–412. [Google Scholar] [CrossRef] [PubMed]
- Klibi, J.; Niki, T. Blood diffusion and Th1-suppressive effects of galectin-9-containing exosomes released by Epstein-Barr virus-infected nasopharyngeal carcinoma cells. Blood 2009, 113, 1957–1966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.; Jiang, J. Tumor-related exosomes contribute to tumor-promoting microenvironment: An immunological perspective. J. Immunol. Res. 2017, 2017, 1073947. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Chen, J. Nasopharyngeal cancer cell-derived exosomal PD-L1 inhibits CD8+ T cell activity and promotes immune escape. Cancer Sci. 2022. [Google Scholar] [CrossRef]
- Lundholm, M.; Schröder, M. Prostate tumor-derived exosomes down-regulate NKG2D expression on natural killer cells and CD8+ T cells: Mechanism of immune evasion. PLoS ONE. 2014, 9, e108925. [Google Scholar] [CrossRef]
- Han, Q.; Zhao, H. HCC-derived exosomes: Critical player and target for cancer immune escape. Cells 2019, 8, 558. [Google Scholar] [CrossRef] [Green Version]
- Ye, S.B.; Zhang, H. Exosomal miR-24-3p impedes T-cell function by targeting FGF11 and serves as a potential prognostic biomarker for nasopharyngeal carcinoma. J. Pathol. 2016, 240, 329–340. [Google Scholar] [CrossRef]
- Yamada, N.; Kuranaga, Y. Colorectal cancer cell-derived extracellular vesicles induce phenotypic alteration of T cells into tumor-growth supporting cells with transforming growth factor-β1-mediated suppression. Oncotarget 2016, 7, 27033–27043. [Google Scholar] [CrossRef] [Green Version]
- Szajnik, M.; Czystowska, M. Tumor-derived microvesicles induce, expand and up-regulate biological activities of human regulatory T cells (Treg). PLoS ONE 2010, 5, e11469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; An, J. Esophageal cancer-derived microvesicles induce regulatory B cells. Cell Biochem. Funct. 2015, 33, 308–313. [Google Scholar] [CrossRef] [PubMed]
- Ye, L.; Zhang, Q. Tumor-derived exosomal HMGB1 fosters hepatocellular carcinoma immune evasion by promoting TIM-1(+) regulatory B cell expansion. J. Immunother Cancer 2018, 6, 145. [Google Scholar] [CrossRef] [Green Version]
- Fabbri, M.; Paone, A. MicroRNAs bind to Toll-like receptors to induce prometastatic inflammatory response. Proc. Natl. Acad. Sci. USA 2012, 109, E2110–E2116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Vrij, J.; Maas, S.L. Glioblastoma-derived extracellular vesicles modify the phenotype of monocytic cells. Int. J. Cancer 2015, 137, 1630–1642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, L.; Zhang, X. Exosomes derived from gastric cancer cells activate NF-κB pathway in macrophages to promote cancer progression. Tumour Biol. 2016, 37, 12169–12180. [Google Scholar] [CrossRef] [PubMed]
- Pai, S.G.; Carneiro, B.A. Wnt/beta-catenin pathway: Modulating anticancer immune response. J. Hematol. Oncol. 2017, 10, 101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menck, K.; Klemm, F. Induction and transport of Wnt 5a during macrophage-induced malignant invasion is mediated by two types of extracellular vesicles. Oncotarget 2013, 4, 2057–2066. [Google Scholar] [CrossRef] [Green Version]
- Altevogt, P.; Bretz, N.P. Novel insights into exosome-induced, tumor-associated inflammation and immunomodulation. Semin. Cancer Biol. 2014, 28, 51–57. [Google Scholar] [CrossRef]
- Chalmin, F.; Ladoire, S. Membrane-associated Hsp72 from tumor-derived exosomes mediates STAT3-dependent immunosuppressive function of mouse and human myeloid-derived suppressor cells. J. Clin. Investig. 2010, 120, 457–471. [Google Scholar] [CrossRef]
- Diao, J.; Yang, X. Exosomal Hsp70 mediates immunosuppressive activity of the myeloid-derived suppressor cells via phosphorylation of Stat3. Med. Oncol. 2015, 32, 453. [Google Scholar] [CrossRef]
- Banas, R.; Miller, C. Amnion-derived multipotent progenitor cells inhibit blood monocyte differentiation into mature dendritic cells. Cell Transplant. 2014, 23, 1111–1125. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Cai, Y. Exosomal LGALS9 in the cerebrospinal fluid of glioblastoma patients suppressed dendritic cell antigen presentation and cytotoxic T-cell immunity. Cell Death Dis. 2020, 11, 896. [Google Scholar] [CrossRef] [PubMed]
- Ding, G.; Zhou, L. Pancreatic cancer-derived exosomes transfer miRNAs to dendritic cells and inhibit RFXAP expression via miR-212-3p. Oncotarget 2015, 6, 29877–29888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maus, R.L.G.; Jakub, J.W. Human melanoma-derived extracellular vesicles regulate dendritic cell maturation. Front. Immunol. 2017, 8, 358. [Google Scholar] [CrossRef] [Green Version]
- Shen, Y.; Guo, D. Tumor-derived exosomes educate dendritic cells to promote tumor metastasis via HSP72/HSP105-TLR2/TLR4 pathway. Oncoimmunology 2017, 6, e1362527. [Google Scholar] [CrossRef]
- Wang, L.; Sun, Z. Extracellular vesicles and the regulation of tumor immunity: Current progress and future directions. J. Cell. Biochem. 2021, 122, 760–769. [Google Scholar] [CrossRef]
- Tian, X.; Shen, H. Tumor-derived exosomes, myeloid-derived suppressor cells, and tumor microenvironment. J. Hematol. Oncol. 2019, 12, 84. [Google Scholar] [CrossRef] [Green Version]
- Xia, Y.; Zhang, Q. Negative regulation of tumor-infiltrating NK cell in clear cell renal cell carcinoma patients through the exosomal pathway. Oncotarget 2017, 8, 37783–37795. [Google Scholar] [CrossRef] [Green Version]
- Xiang, X.; Poliakov, A. Induction of myeloid-derived suppressor cells by tumor exosomes. Int. J. Cancer 2009, 124, 2621–2633. [Google Scholar] [CrossRef] [Green Version]
- Kumar, B.V.; Connors, T.J. Human T cell development, localization, and function throughout life. Immunity 2018, 48, 202–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aurrand-Lions, M.; Mancini, S.J.C. Murine bone marrow niches from hematopoietic stem cells to B cells. Int. J. Mol. Sci. 2018, 19, 2353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alemohammad, H.; Najafzadeh, B. The importance of immune checkpoints in immune monitoring: A future paradigm shift in the treatment of cancer. Biomed. Pharmacother. 2022, 146, 112516. [Google Scholar] [CrossRef] [PubMed]
- Dyck, L.; Mills, K.H.G. Immune checkpoints and their inhibition in cancer and infectious diseases. Eur. J. Immunol. 2017, 47, 765–779. [Google Scholar] [CrossRef]
- Dolina, J.S.; Van Braeckel-Budimir, N. CD8+ T cell exhaustion in cancer. Front. Immunol. 2021, 12, 715234. [Google Scholar] [CrossRef]
- Lu, L.; Bai, Y. Elevated T cell activation score is associated with improved survival of breast cancer. Breast Cancer Res. Treat. 2017, 64, 689–696. [Google Scholar] [CrossRef]
- Lu, L.; Huang, H. BRCA1 mRNA expression modifies the effect of T cell activation score on patient survival in breast cancer. BMC Cancer 2019, 19, 387. [Google Scholar] [CrossRef] [Green Version]
- Lu, L.; Gaffney, G.S. Transfer RNA methyltransferase gene NSUN2 mRNA expression modifies the effect of T cell activation score on patient survival in head and neck squamous carcinoma. Oral Oncol. 2020, 101, 104554. [Google Scholar] [CrossRef]
- Keir, M.E.; Butte, M.J. PD-1 and its ligands in tolerance and immunity. Annu. Rev. Immunol. 2008, 26, 677–704. [Google Scholar] [CrossRef] [Green Version]
- Ohue, Y.; Nishikawa, H. Regulatory T (Treg) cells in cancer: Can Treg cells be a new therapeutic target? Cancer Sci. 2019, 110, 2080–2089. [Google Scholar] [CrossRef]
- Mrizak, D.; Martin, N. Effect of nasopharyngeal carcinoma-derived exosomes on human regulatory T cells. J. Natl. Cancer Inst. 2015, 107, 363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orecchioni, M.; Ghosheh, Y. Macrophage polarization: Different gene signatures in M1(LPS+) vs. classically and M2(LPS−) vs. alternatively activated macrophages. Front. Immunol. 2019, 10, 1084. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A.; Sozzani, S. Macrophage polarization: Tumor-associated macrophages as a paradigm for polarized M2 mononuclear phagocytes. Trends Immunol. 2002, 23, 549–555. [Google Scholar] [CrossRef]
- Shapouri-Moghaddam, A.; Mohammadian, S. Macrophage plasticity, polarization, and function in health and disease. J. Cell. Physiol. 2018, 233, 6425–6440. [Google Scholar] [CrossRef]
- Baig, M.S.; Roy, A. Tumor-derived exosomes in the regulation of macrophage polarization. Inflamm. Res. 2020, 69, 435–451. [Google Scholar] [CrossRef]
- Cai, J.; Qiao, B. Oral squamous cell carcinoma-derived exosomes promote M2 subtype macrophage polarization mediated by exosome-enclosed miR-29a-3p. Am. J. Physiol. Cell Physiol. 2019, 316, C731–C740. [Google Scholar] [CrossRef]
- Hsu, Y.L.; Hung, J.Y. Hypoxic lung cancer-secreted exosomal miR-23a increased angiogenesis and vascular permeability by targeting prolyl hydroxylase and tight junction protein ZO-1. Oncogene 2017, 36, 4929–4942. [Google Scholar] [CrossRef]
- Hsu, Y.L.; Hung, J.Y. Hypoxic lung-cancer-derived extracellular vesicle microRNA-103a increases the oncogenic effects of macrophages by targeting PTEN. Mol. Ther. 2018, 26, 568–581. [Google Scholar] [CrossRef] [Green Version]
- Jeong, H.J.; Oh, H.A. The critical role of mast cell-derived hypoxia-inducible factor-1α in human and mice melanoma growth. Int. J. Cancer 2013, 132, 2492–2501. [Google Scholar] [CrossRef]
- Gardner, A.; de Mingo Pulido, Á. Dendritic cells and their role in immunotherapy. Front. Immunol. 2020, 11, 924. [Google Scholar] [CrossRef]
- Wculek, S.K.; Cueto, F.J. Dendritic cells in cancer immunology and immunotherapy. Nat. Rev. Immunol. 2020, 20, 7–24. [Google Scholar] [CrossRef] [PubMed]
- He, T.; Tang, C. Interferon gamma stimulates cellular maturation of dendritic cell line DC2.4 leading to induction of efficient cytotoxic T cell responses and antitumor immunity. Cell. Mol. Immunol. 2007, 4, 105–111. [Google Scholar] [PubMed]
- Chen, M.; Felix, K. Critical role for perforin and Fas-dependent killing of dendritic cells in the control of inflammation. Blood 2012, 119, 127–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tesone, A.J.; Svoronos, N. Pathological mobilization and activities of dendritic cells in tumor-bearing hosts: Challenges and opportunities for immunotherapy of cancer. Front. Immunol. 2013, 4, 435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sombroek, C.C.; Stam, A.G. Prostanoids play a major role in the primary tumor-induced inhibition of dendritic cell differentiation. J. Immunol. 2002, 168, 4333–4343. [Google Scholar] [CrossRef]
- Husain, Z.; Huang, Y. Tumor-derived lactate modifies antitumor immune response: Effect on myeloid-derived suppressor cells and NK cells. J. Immunol. 2013, 191, 1486–1495. [Google Scholar] [CrossRef]
- Valenti, R.; Huber, V. Human tumor-released microvesicles promote the differentiation of myeloid cells with transforming growth factor-beta-mediated suppressive activity on T lymphocytes. Cancer Res. 2006, 66, 9290–9298. [Google Scholar] [CrossRef] [Green Version]
- Valenti, R.; Huber, V. Tumor-released microvesicles as vehicles of immunosuppression. Cancer Res. 2007, 67, 2912–2915. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Fei, X. Tumor-derived extracellular vesicles: Regulators of tumor microenvironment and the enlightenment in tumor therapy. Pharmacol. Res. 2020, 159, 105041. [Google Scholar] [CrossRef]
- Olejarz, W.; Dominiak, A. Tumor-derived exosomes in immunosuppression and immunotherapy. J. Immunol. Res. 2020, 2020, 6272498. [Google Scholar] [CrossRef]
- Whiteside, T.L. Immune modulation of T-cell and NK (natural killer) cell activities by TEXs (tumour-derived exosomes). Biochem. Soc. Trans. 2013, 41, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Kugeratski, F.G.; Kalluri, R. Exosomes as mediators of immune regulation and immunotherapy in cancer. FEBS J. 2021, 288, 10–35. [Google Scholar] [CrossRef] [PubMed]
- Hosseini, R.; Sarvnaz, H. Cancer exosomes and natural killer cells dysfunction: Biological roles, clinical significance and implications for immunotherapy. Mol. Cancer 2022, 21, 15. [Google Scholar] [CrossRef] [PubMed]
- Baragaño Raneros, A.; López-Larrea, C. Acute myeloid leukemia and NK cells: Two warriors confront each other. Oncoimmunology 2019, 8, e1539617. [Google Scholar] [CrossRef] [Green Version]
- Schwarzenbach, H.; Gahan, P.B. Exosomes in immune regulation. Non-Coding RNA 2021, 7, 4. [Google Scholar] [CrossRef]
- Liu, C.; Yu, S. Murine mammary carcinoma exosomes promote tumor growth by suppression of NK cell function. J. Immunol. 2006, 176, 1375–1385. [Google Scholar] [CrossRef] [Green Version]
- Groth, C.; Hu, X. Immunosuppression mediated by myeloid-derived suppressor cells (MDSCs) during tumour progression. Br. J. Cancer 2019, 120, 16–25. [Google Scholar] [CrossRef] [Green Version]
- Greten, T.F.; Manns, M.P. Myeloid derived suppressor cells in human diseases. Int. Immunopharmacol. 2011, 11, 802–807. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.; Qiu, W. Glioma exosomes mediate the expansion and function of myeloid-derived suppressor cells through microRNA-29a/Hbp1 and microRNA-92a/Prkar1a pathways. Int. J. Cancer 2019, 144, 3111–3126. [Google Scholar] [CrossRef]
- Wang, M.; Zhang, B. The immunomodulation potential of exosomes in tumor microenvironment. J. Immunol. Res. 2021, 2021, 3710372. [Google Scholar] [CrossRef]
- Li, L.; Cao, B. Microenvironmental oxygen pressure orchestrates an anti- and pro-tumoral γδ T cell equilibrium via tumor-derived exosomes. Oncogene 2019, 38, 2830–2843. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Wu, Y. Infiltration of alternatively activated macrophages in cancer tissue is associated with MDSC and Th2 polarization in patients with esophageal cancer. PLoS ONE 2014, 9, e104453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dou, D.; Ren, X. Cancer-associated fibroblasts-derived exosomes suppress immune cell function in breast cancer via the miR-92/PD-L1 pathway. Front. Immunol. 2020, 11, 2026. [Google Scholar] [CrossRef] [PubMed]
- Vergani, E.; Daveri, E. Extracellular vesicles in anti-tumor immunity. Semin. Cancer Biol. 2021, in press. [CrossRef]
- Sullivan, J.A.; Tomita, Y. Treg-cell-derived IL-35-coated extracellular vesicles promote infectious tolerance. Cell Rep. 2020, 30, 1039–1051.E5. [Google Scholar] [CrossRef]
- Okoye, I.S.; Coomes, S.M. MicroRNA-containing T-regulatory-cell-derived exosomes suppress pathogenic T helper 1 cells. Immunity 2014, 41, 89–103. [Google Scholar] [CrossRef] [Green Version]
- Chersi, A.; Houghten, R.A. Recognition of HLA class II molecules by antipeptide antibodies elicited by synthetic peptides selected from regions of HLA-DP antigens. Z. Naturforsch. C. J. Biosci. 1987, 42, 1313–1318. [Google Scholar] [CrossRef]
- Burke, M.; Choksawangkarn, W. Exosomes from myeloid-derived suppressor cells carry biologically active proteins. J. Proteome Res. 2014, 13, 836–843. [Google Scholar] [CrossRef] [Green Version]
- Geis-Asteggiante, L.; Dhabaria, A. Top-down analysis of low mass proteins in exosomes shed by murine myeloid-derived suppressor cells. Int. J. Mass Spectrom. 2015, 378, 264–269. [Google Scholar] [CrossRef] [Green Version]
- Hegde, V.L.; Tomar, S. Distinct microRNA expression profile and targeted biological pathways in functional myeloid-derived suppressor cells induced by Δ9-tetrahydrocannabinol in vivo: Regulation of CCAAT/enhancer-binding protein α by microRNA-690. J. Biol. Chem. 2013, 288, 36810–36826. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Zhang, Y. Regulating tumor myeloid-derived suppressor cells by microRNAs. Cancer Cell Microenviron. 2015, 2, e637. [Google Scholar] [PubMed]
- Parker, K.H.; Beury, D.W. Myeloid-derived suppressor cells: Critical cells driving immune suppression in the tumor microenvironment. Adv. Cancer Res. 2015, 128, 95–139. [Google Scholar] [PubMed] [Green Version]
- Kroesen, B.J.; Teteloshvili, N. Immuno-miRs: Critical regulators of T-cell development, function and ageing. Immunology 2015, 144, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Rashid, M.H.; Borin, T.F. Critical immunosuppressive effect of MDSC-derived exosomes in the tumor microenvironment. Oncol. Rep. 2021, 45, 1171–1181. [Google Scholar] [CrossRef]
- Geis-Asteggiante, L.; Belew, A.T. Differential content of proteins, mRNAs, and miRNAs suggests that MDSC and their exosomes may mediate distinct immune suppressive functions. J. Proteome Res. 2018, 17, 486–498. [Google Scholar] [CrossRef]
- Zöller, M. Janus-Faced Myeloid-derived suppressor cell exosomes for the good and the bad in cancer and autoimmune disease. Front. Immunol. 2018, 9, 137. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.S.; Wu, T.M. M2 macrophage-derived exosomal microRNA-155-5p promotes the immune escape of colon cancer by downregulating ZC3H12B. Mol. Ther. Oncolytics 2021, 20, 484–498. [Google Scholar] [CrossRef]
- Lu, L.; Ling, W. TAM-derived extracellular vesicles containing microRNA-29a-3p explain the deterioration of ovarian cancer. Mol. Ther. Nucleic Acids 2021, 25, 468–482. [Google Scholar] [CrossRef]
- Han, C.; Zhang, C. Exosome-mediated communication between tumor cells and tumor-associated macrophages: Implications for tumor microenvironment. Oncoimmunology 2021, 10, 1887552. [Google Scholar] [CrossRef]
- Kalluri, R. The biology and function of fibroblasts in cancer. Nat. Rev. Cancer 2016, 16, 582–598. [Google Scholar] [CrossRef]
- Mao, X.; Xu, J. Crosstalk between cancer-associated fibroblasts and immune cells in the tumor microenvironment: New findings and future perspectives. Mol. Cancer 2021, 20, 131. [Google Scholar] [CrossRef] [PubMed]
- Smyth, L.A.; Ratnasothy, K. CD73 expression on extracellular vesicles derived from CD4+ CD25+ Foxp3+ T cells contributes to their regulatory function. Eur. J. Immunol. 2013, 43, 2430–2440. [Google Scholar] [CrossRef] [PubMed]
- Turnis, M.E.; Sawant, D.V. Interleukin-35 limits anti-tumor immunity. Immunity 2016, 44, 316–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raposo, G.; Stoorvogel, W. Extracellular vesicles: Exosomes, microvesicles, and friends. J. Cell Biol. 2013, 200, 373–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, Y.; Chen, Y. Exosome: Emerging biomarker in breast cancer. Oncotarget. 2017, 8, 41717–41733. [Google Scholar] [CrossRef] [Green Version]
- Melo, S.A.; Luecke, L.B. Glypican-1 identifies cancer exosomes and detects early pancreatic cancer. Nature 2015, 523, 177–182. [Google Scholar] [CrossRef] [Green Version]
- Kok, V.C.; Yu, C.C. Cancer-derived exosomes: Their role in cancer biology and biomarker development. Int. J. Nanomed. 2020, 15, 8019–8036. [Google Scholar] [CrossRef]
- Lunavat, T.R.; Cheng, L. Small RNA deep sequencing discriminates subsets of extracellular vesicles released by melanoma cells--Evidence of unique microRNA cargos. RNA Biol. 2015, 12, 810–823. [Google Scholar] [CrossRef] [Green Version]
- Whiteside, T.L. The potential of tumor-derived exosomes for noninvasive cancer monitoring. Expert Rev. Mol. Diagn. 2015, 15, 1293–1310. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Zeng, H. A circulating exosome RNA signature is a potential diagnostic marker for pancreatic cancer, a systematic study. Cancers 2021, 13, 2565. [Google Scholar] [CrossRef]
- Xu, H.; Dong, X. Serum exosomal hnRNPH1 mRNA as a novel marker for hepatocellular carcinoma. Clin. Chem. Lab. Med. 2018, 56, 479–484. [Google Scholar] [CrossRef] [PubMed]
- Yokota, Y.; Noda, T. Serum exosomal miR-638 is a prognostic marker of HCC via downregulation of VE-cadherin and ZO-1 of endothelial cells. Cancer Sci. 2021, 112, 1275–1288. [Google Scholar] [CrossRef] [PubMed]
- Yugawa, K.; Yoshizumi, T. Cancer-associated fibroblasts promote hepatocellular carcinoma progression through downregulation of exosomal miR-150-3p. Eur. J. Surg. Oncol. 2021, 47, 384–393. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.; Jiang, Y. Decreased levels of serum exosomal miR-638 predict poor prognosis in hepatocellular carcinoma. J. Cell. Biochem. 2018, 119, 4711–4716. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Zhu, H. Serum exosomal long noncoding RNA CRNDE as a prognostic biomarker for hepatocellular carcinoma. J. Clin. Lab. Anal. 2021, 35, e23959. [Google Scholar] [CrossRef]
- Yu, S.; Li, Y. Plasma extracellular vesicle long RNA profiling identifies a diagnostic signature for the detection of pancreatic ductal adenocarcinoma. Gut 2020, 69, 540–550. [Google Scholar] [CrossRef]
- Huang, T.; Deng, C.X. Current progresses of exosomes as cancer diagnostic and prognostic biomarkers. Int. J. Biol. Sci. 2019, 15, 1–11. [Google Scholar] [CrossRef]
- Beccard, I.J.; Hofmann, L. Immune suppressive effects of plasma-derived exosome populations in head and neck cancer. Cancers 2020, 12, 7. [Google Scholar] [CrossRef]
- Sharma, P.; Diergaarde, B. Melanoma cell-derived exosomes in plasma of melanoma patients suppress functions of immune effector cells. Sci. Rep. 2020, 10, 92. [Google Scholar] [CrossRef]
- Wei, Q.; Feng, H. Serum exosomal EphA2 is a prognostic biomarker in patients with pancreatic cancer. Cancer Manag. Res. 2021, 13, 3675–3683. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, Y. Plasma-derived exosomal ALIX as a novel biomarker for diagnosis and classification of pancreatic cancer. Front. Oncol. 2021, 11, 628346. [Google Scholar] [CrossRef] [PubMed]
- Hong, C.S.; Muller, L. Plasma exosomes as markers of therapeutic response in patients with acute myeloid leukemia. Front. Immunol. 2014, 5, 160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Niu, X. Exosomal PD-L1 predicts response with immunotherapy in NSCLC patients. Clin. Exp. Immunol. 2022. [Google Scholar] [CrossRef] [PubMed]
- Momen-Heravi, F. Isolation of extracellular vesicles by ultracentrifugation. Methods Mol. Biol. 2017, 1660, 25–32. [Google Scholar]
- Merchant, M.L.; Powell, D.W. Microfiltration isolation of human urinary exosomes for characterization by MS. Proteomics Clin. Appl. 2010, 4, 84–96. [Google Scholar] [CrossRef]
- Baranyai, T.; Herczeg, K. Isolation of exosomes from blood plasma: Qualitative and quantitative comparison of ultracentrifugation and size exclusion chromatography methods. PLoS ONE 2015, 10, e0145686. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.; Sun, H.T. Isolation and characterization of exosomes for cancer research. J. Hematol. Oncol. 2020, 13, 152. [Google Scholar] [CrossRef]
- Shen, M.; Di, K. Progress in exosome associated tumor markers and their detection methods. Mol. Biomed. 2020, 1, 3. [Google Scholar] [CrossRef]
- Ramirez, M.I.; Amorim, M.G. Technical challenges of working with extracellular vesicles. Nanoscale 2018, 10, 881–906. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Zeng, X. Sensitive detection of exosomal proteins via a compact surface plasmon resonance biosensor for cancer diagnosis. ACS Sens. 2018, 3, 1471–1479. [Google Scholar] [CrossRef]
- Carney, R.P.; Hazari, S. Multispectral optical tweezers for biochemical fingerprinting of CD9-positive exosome subpopulations. Anal. Chem. 2017, 89, 5357–5363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Guo, J. Field-free isolation of exosomes from extracellular vesicles by microfluidic viscoelastic flows. ACS Nano 2017, 11, 6968–6976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hosseini, R.; Asef-Kabiri, L. The roles of tumor-derived exosomes in altered differentiation, maturation and function of dendritic cells. Mol. Cancer 2021, 20, 83. [Google Scholar] [CrossRef] [PubMed]
- Datta, A.; Kim, H. Manumycin A suppresses exosome biogenesis and secretion via targeted inhibition of Ras/Raf/ERK1/2 signaling and hnRNP H1 in castration-resistant prostate cancer cells. Cancer Lett. 2017, 408, 73–81. [Google Scholar] [CrossRef]
- Datta, A.; Kim, H. High-throughput screening identified selective inhibitors of exosome biogenesis and secretion: A drug repurposing strategy for advanced cancer. Sci. Rep. 2018, 8, 8161. [Google Scholar] [CrossRef] [Green Version]
- Im, E.J.; Lee, C.H. Sulfisoxazole inhibits the secretion of small extracellular vesicles by targeting the endothelin receptor A. Nat. Commun. 2019, 10, 1387. [Google Scholar] [CrossRef]
- Catalano, M.; O’Driscoll, L. Inhibiting extracellular vesicles formation and release: A review of EV inhibitors. J. Extracell. Vesicles 2020, 9, 1703244. [Google Scholar] [CrossRef] [Green Version]
- Menck, K.; Sönmezer, C. Neutral sphingomyelinases control extracellular vesicles budding from the plasma membrane. J. Extracell. Vesicles 2017, 6, 1378056. [Google Scholar] [CrossRef]
- McKelvey, K.J.; Powell, K.L. Exosomes: Mechanisms of uptake. J. Circ. Biomark. 2015, 4, 7. [Google Scholar] [CrossRef] [Green Version]
- Kirchhausen, T.; Macia, E. Use of dynasore, the small molecule inhibitor of dynamin, in the regulation of endocytosis. Methods Enzymol. 2008, 438, 77–93. [Google Scholar]
- Hayatudin, R.; Fong, Z. Overcoming chemoresistance via extracellular vesicle inhibition. Front. Mol. Biosci. 2021, 8, 629874. [Google Scholar] [CrossRef] [PubMed]
- Christianson, H.C.; Svensson, K.J. Cancer cell exosomes depend on cell-surface heparan sulfate proteoglycans for their internalization and functional activity. Proc. Natl. Acad. Sci. USA 2013, 110, 17380–17385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atai, N.A.; Balaj, L. Heparin blocks transfer of extracellular vesicles between donor and recipient cells. J. Neurooncol. 2013, 115, 343–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Target Cells | Functional Molecules | Effects | References |
|---|---|---|---|
| T cells | Galectin-9 | Apoptosis of EBV-specific CD4 T cells and inhibition on Th1 cell function | [14] |
| FasL | Apoptosis of T cells | [15] | |
| PD-L1 | Inhibited proliferation and functions of T cells | [16] | |
| ULBP/MICA | Inhibition on the NKG2D signaling pathway | [17] | |
| TGF-β | Downregulation of NKG2D on CD8 T cells | [18] | |
| MiRNAs | Inhibition on the differentiation of Th cells | [19] | |
| Tregs | TGF-β | Upregulation of Treg-related genes | [20] |
| IL-10/IL-2 | Increased amount and enhanced function of Tregs | [21] | |
| Bregs | —— | Differentiation into TGF-β-producing Bregs | [22] |
| HMGB1 | Increased TIM-1 Breg cells | [23] | |
| Macrophages | miRNAs | Promotion on the IL-6 secretion of immune cells | [24] |
| CSPG4, EGFR, and integrins | Increased M2 macrophages | [25] | |
| —— | Increased proinflammatory factors | [26] | |
| Wnt5a | Improvement on the invasion ability of tumor | [27,28] | |
| DCs | IL-6, HSP70, and HSP72 | Activation of STAT3 | [29,30,31] |
| HLA-G | Suppression on T cells, NK cells, and DCs | [32] | |
| Galectin-9 | Inhibition on the maturation and failure of activating cytotoxic T cells | [33] | |
| miRNAs | Inhibited RFXAP and toll-like receptor 4 (TLR4) expression in DCs | [34] | |
| S100A9 | Decreased costimulatory molecules on DCs | [35] | |
| HSP72 and HSP105 | Increased secretion of IL-6 of DCs | [36] | |
| NK cells | TGF-β | Downregulation of NKG2D and reduced cytolytic activity of NK cells | [37] |
| MDSCs | miRNAs | Activation of STAT1 and STAT3 pathways and expansion of MDSCs | [38,39] |
| —— | Release of NO from MDSCs | [15] | |
| HSP72/TLR2 | Autocrine production of IL-6 | [30] | |
| PGE2 and TGF-β | Differentiation of MDSCs from myeloid cells | [40] |
| Origin of Exosomes | Target Cells | Functional Molecules | Mechanisms | References |
|---|---|---|---|---|
| Cancer-associated fibroblasts | PD-L1 | miR-92 | Promote YAP1 nuclear translocation and increase PD-L1 transcription | [83] |
| Tregs | T cells and B cells | IL-35 | Induce peripheral tolerance of T cells and B cells by transferring IL-35 | [84,85] |
| T cells and DCs | miRNAs | Inhibit proliferation and cytokine release of Th1 cells and DCs | [86,87] | |
| MDSCs | MDSCs | S100A8 and S100A9 | Mediate accumulation and immunosuppressive function of MDSCs | [88,89] |
| MDSCs | miRNAs | Promote expansion, differentiation survival, and IL-10 production of MDSCs | [90,91,92,93] | |
| T cells | —— | Induce apoptosis of T cells | [94,95] | |
| Macrophages | —— | Induce decline of M1 macrophages and expansion of M2 macrophages | [96] | |
| Tumor-associated macrophages | —— | miR-155-5p | Impair stability of IL-6 | [97] |
| Ovarian cancer cells | miR-29a-3p | Increase expression of PD-L1 | [98] | |
| T cells | —— | Inhibit cell cytotoxicity of CD8 T cells | [99] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, Q.-H.; Zheng, J.-Q.; Ding, J.-Y.; Wu, Y.-F.; Liu, L.; Yu, Z.-L.; Chen, G. Exosome-Mediated Immunosuppression in Tumor Microenvironments. Cells 2022, 11, 1946. https://doi.org/10.3390/cells11121946
Xie Q-H, Zheng J-Q, Ding J-Y, Wu Y-F, Liu L, Yu Z-L, Chen G. Exosome-Mediated Immunosuppression in Tumor Microenvironments. Cells. 2022; 11(12):1946. https://doi.org/10.3390/cells11121946
Chicago/Turabian StyleXie, Qi-Hui, Ji-Qi Zheng, Jia-Yi Ding, Yu-Fei Wu, Luisa Liu, Zi-Li Yu, and Gang Chen. 2022. "Exosome-Mediated Immunosuppression in Tumor Microenvironments" Cells 11, no. 12: 1946. https://doi.org/10.3390/cells11121946
APA StyleXie, Q. -H., Zheng, J. -Q., Ding, J. -Y., Wu, Y. -F., Liu, L., Yu, Z. -L., & Chen, G. (2022). Exosome-Mediated Immunosuppression in Tumor Microenvironments. Cells, 11(12), 1946. https://doi.org/10.3390/cells11121946