
Neuroinflammatory Gene Expression Analysis Reveals Pathways of Interest as Potential Targets to Improve the Recording Performance of Intracortical Microelectrodes


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Microelectrodes
2.3. Surgical Procedure
2.4. Tissue Extraction
2.5. RNA Isolation
2.6. Gene Expression Assay
2.7. Data Visualization and Statistical Analysis
2.7.1. Normalization
2.7.2. Heatmap and Principal Component Analysis
2.7.3. Comparison of Gene Expression at Each Post-Surgical Time Point to Naïve Non-Surgical Control
3. Results
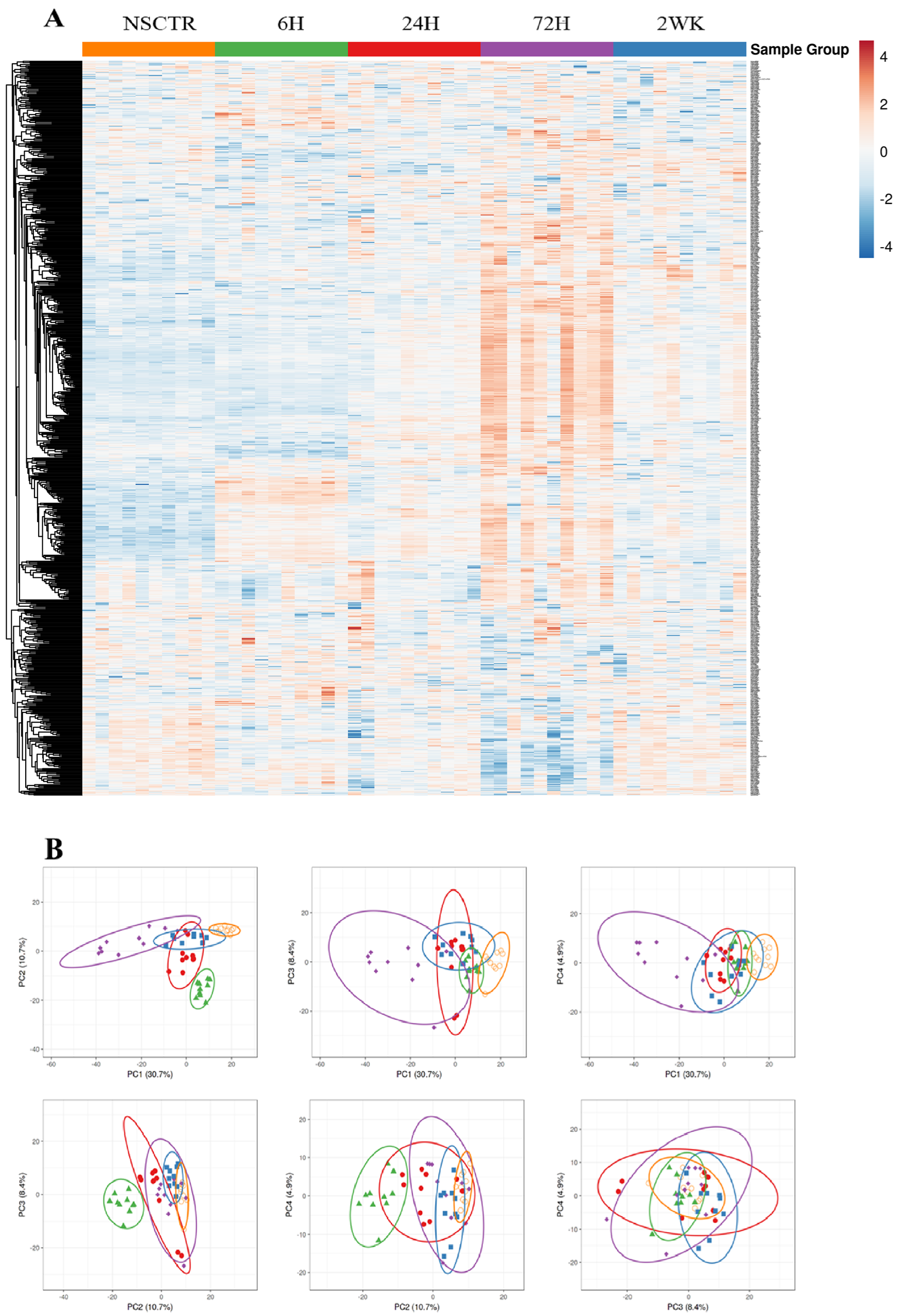
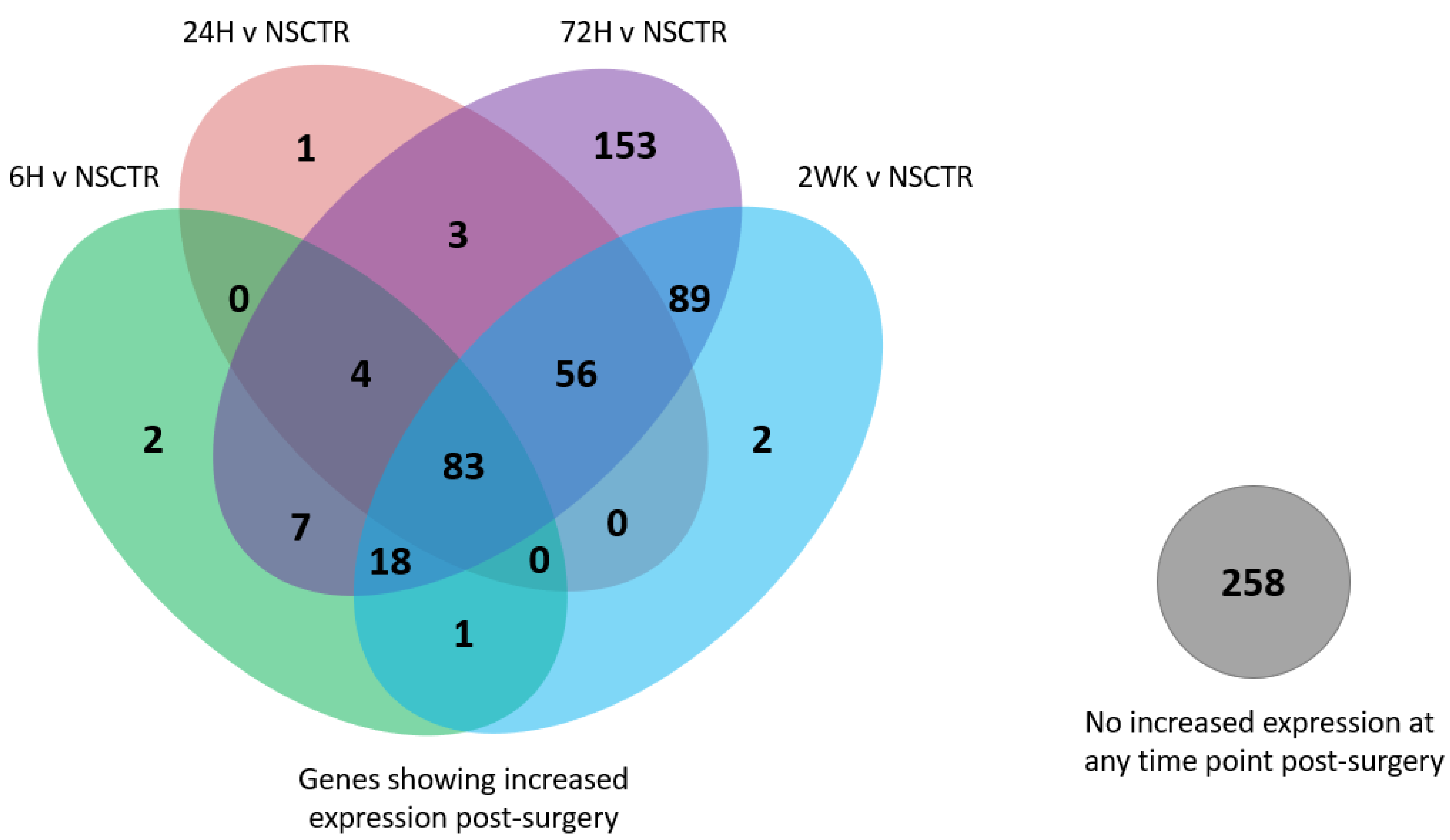
3.1. Overall Gene Expression
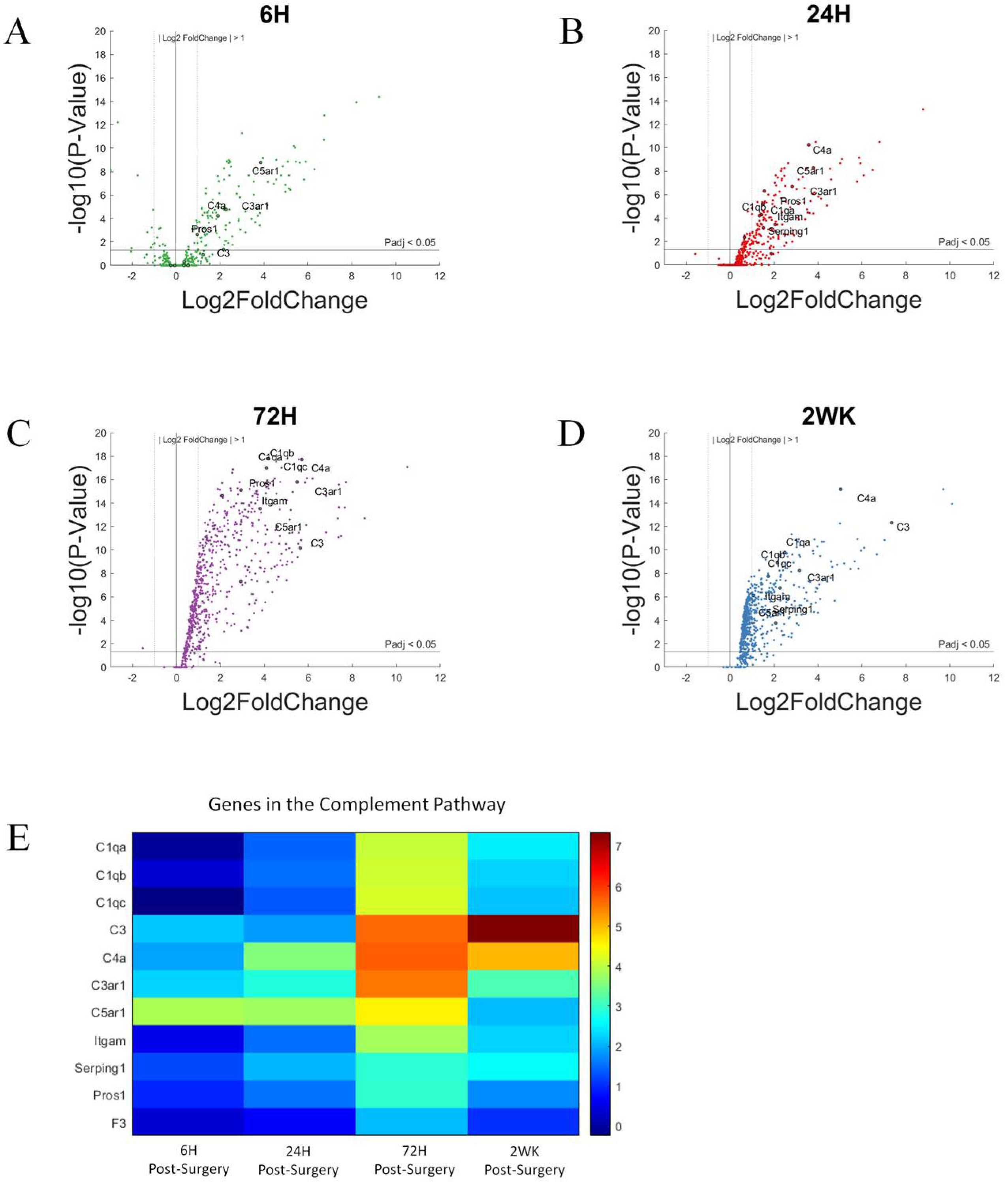
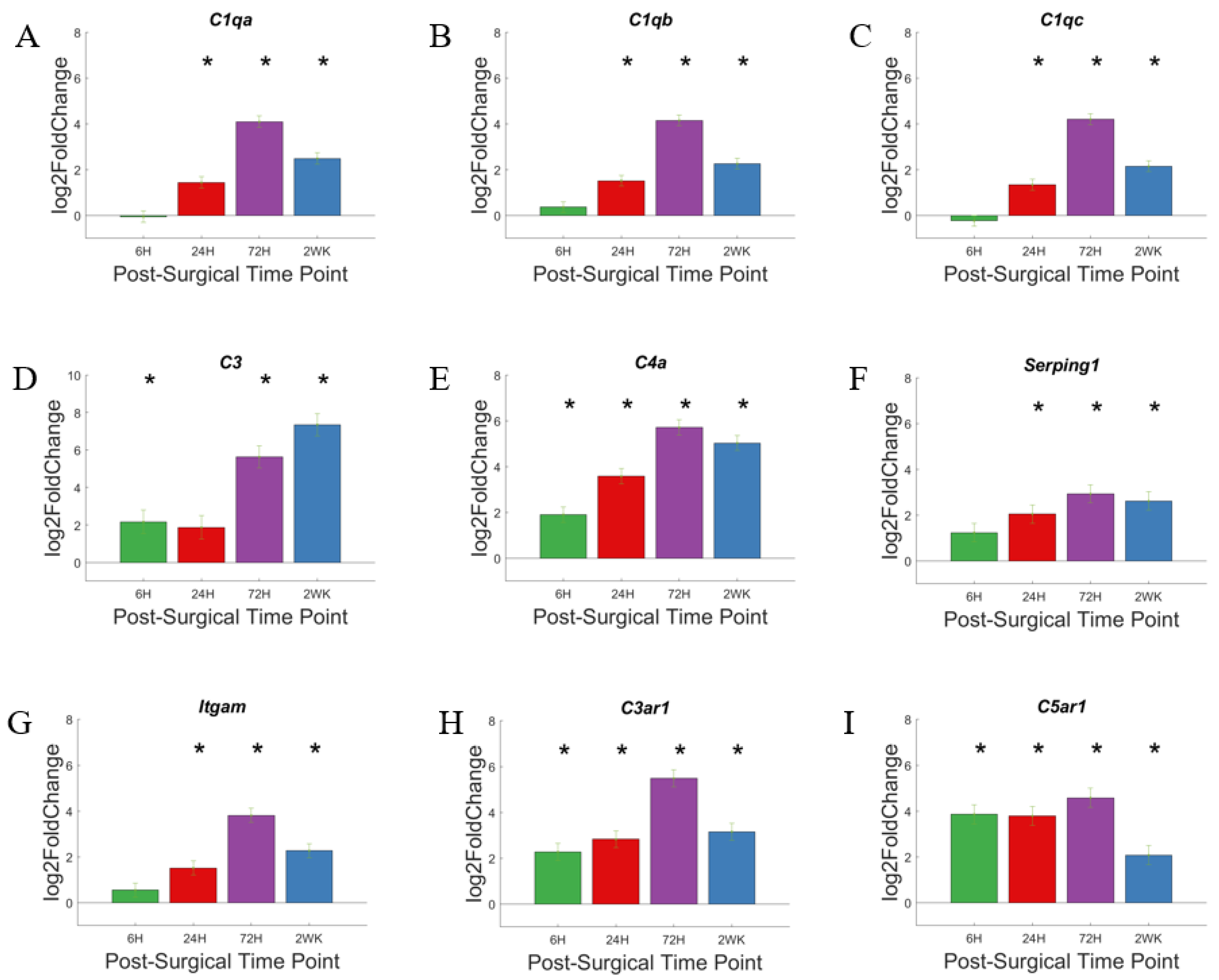
3.2. The Complement Pathway
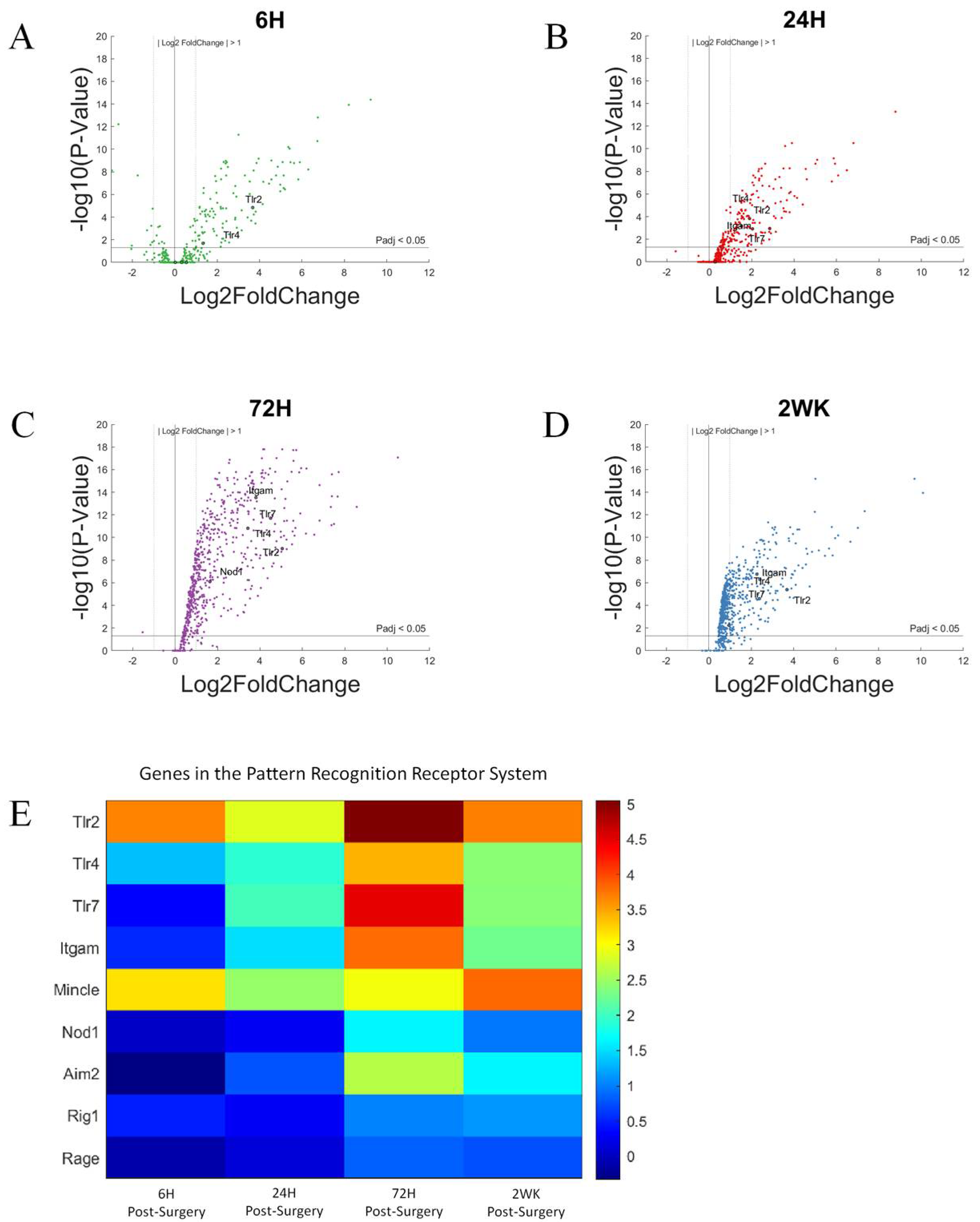
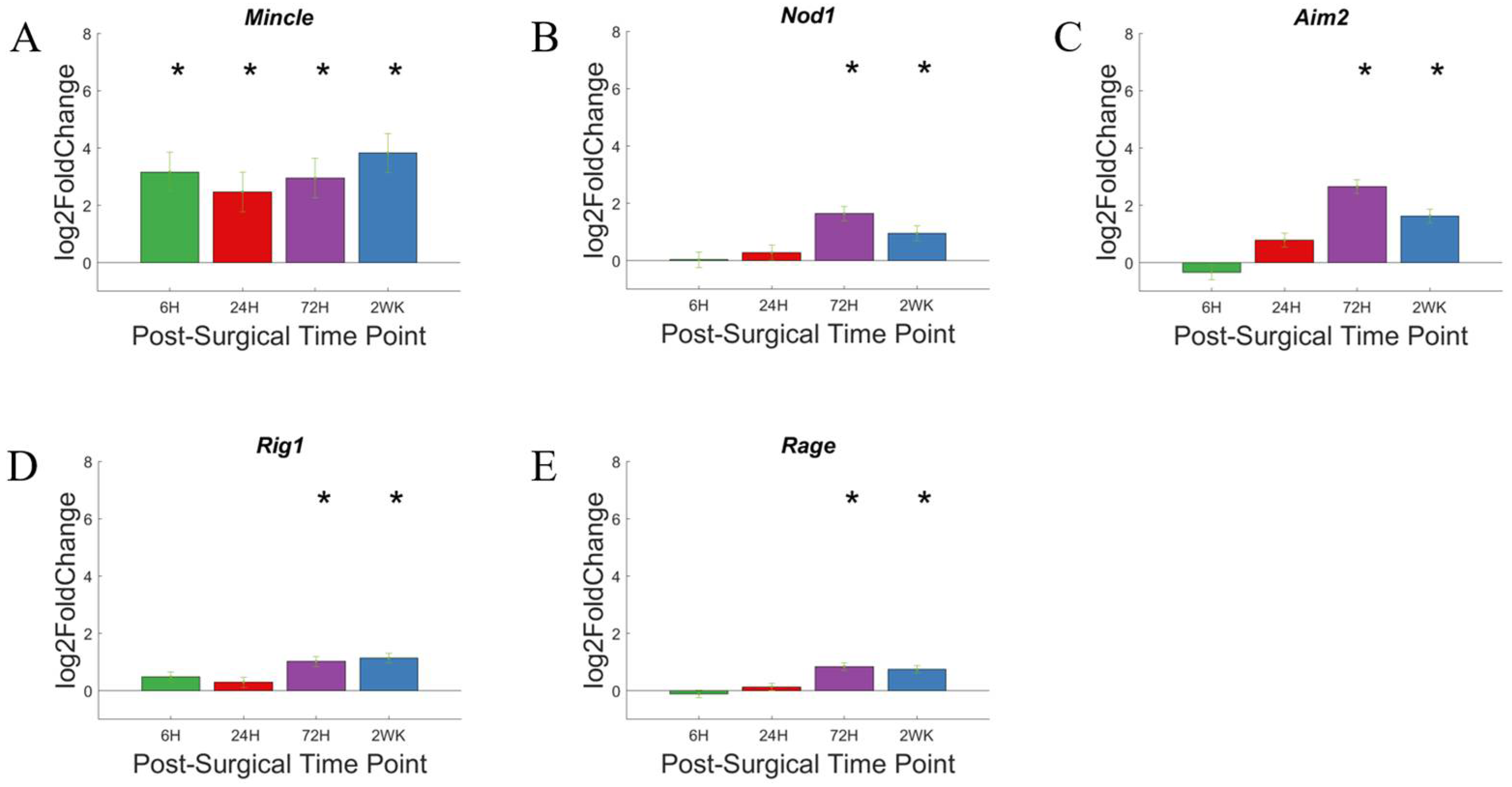
3.3. Pattern Recognition Receptors
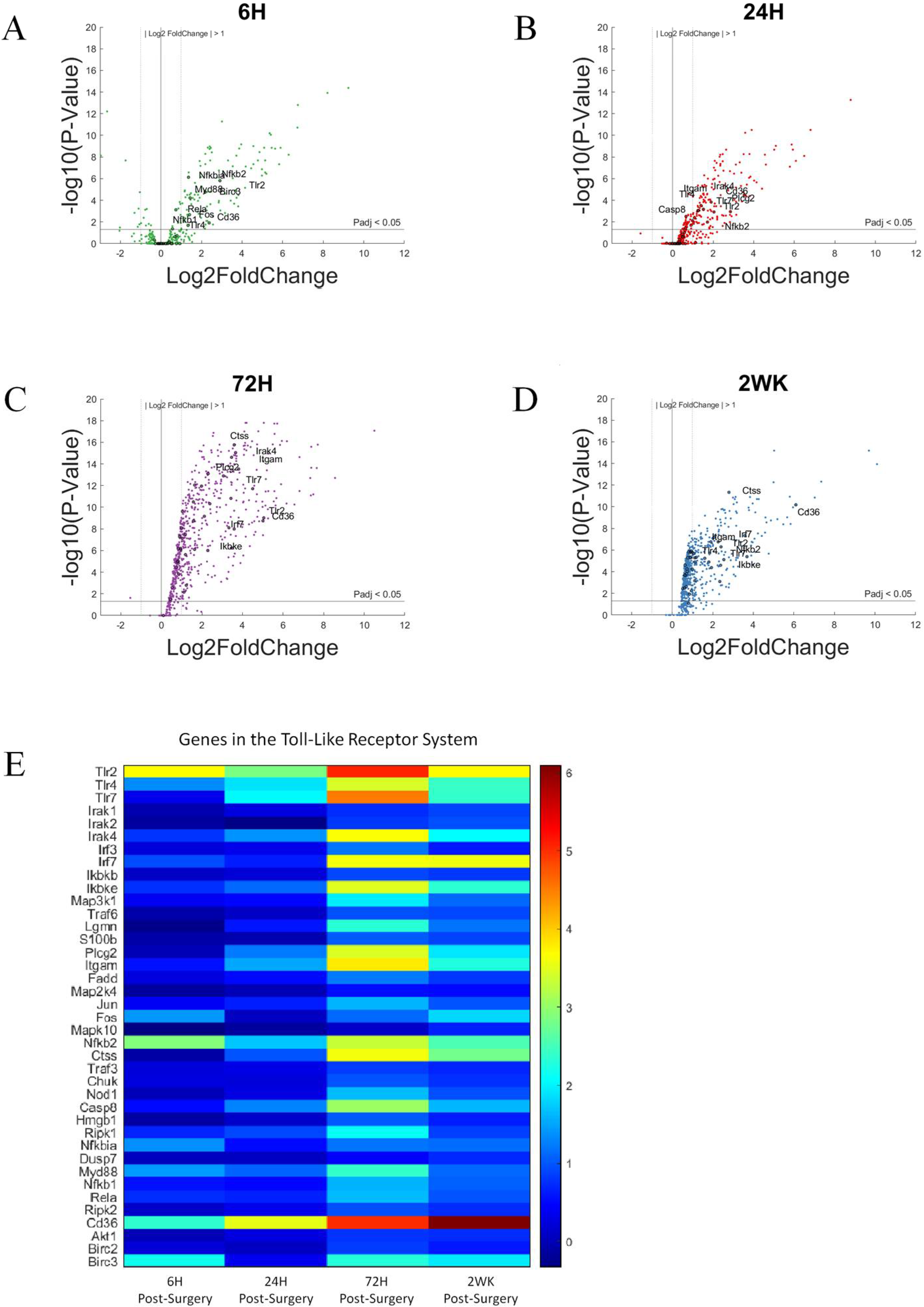
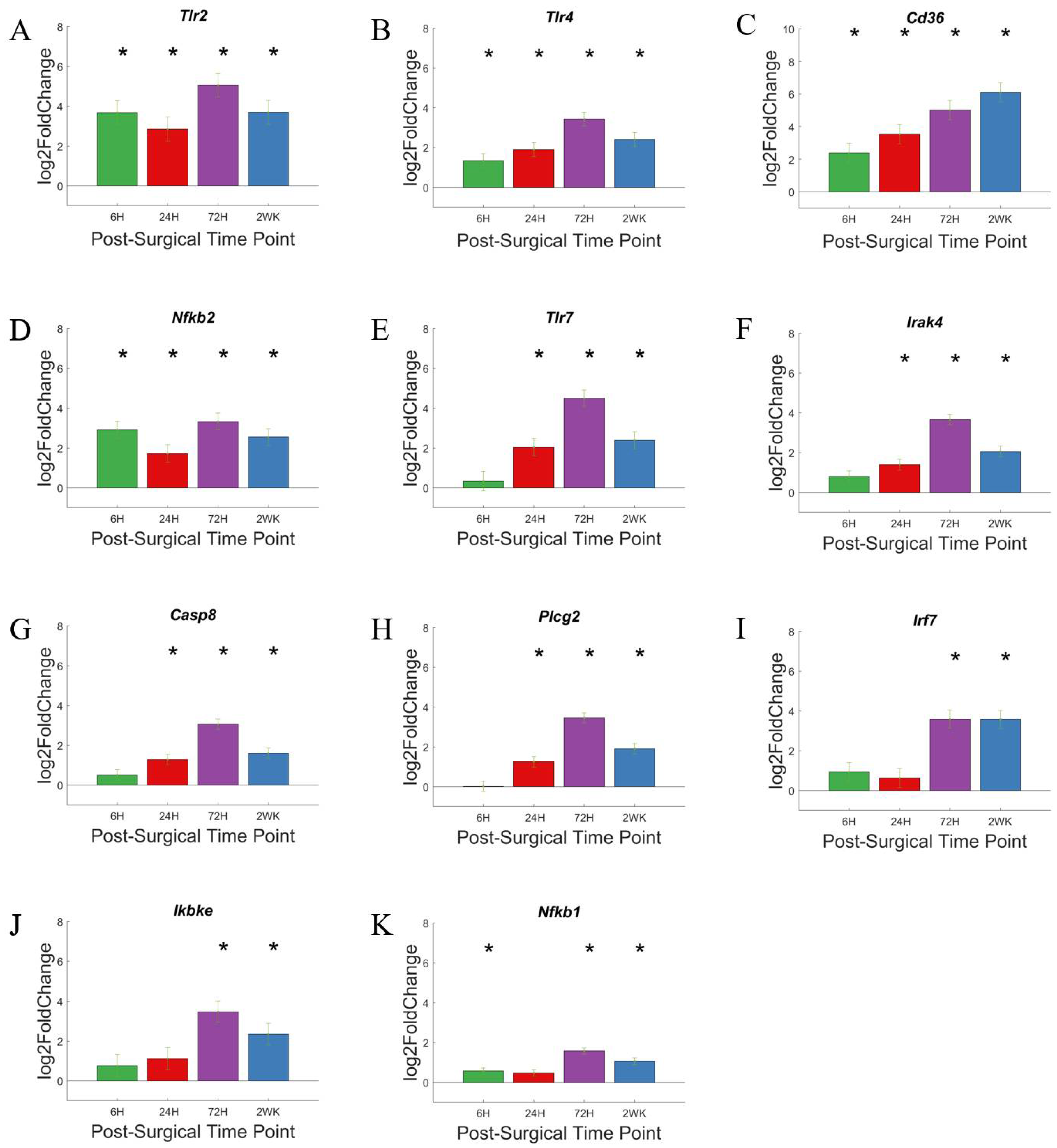
3.4. Toll-Like Receptors and Associated Pathways
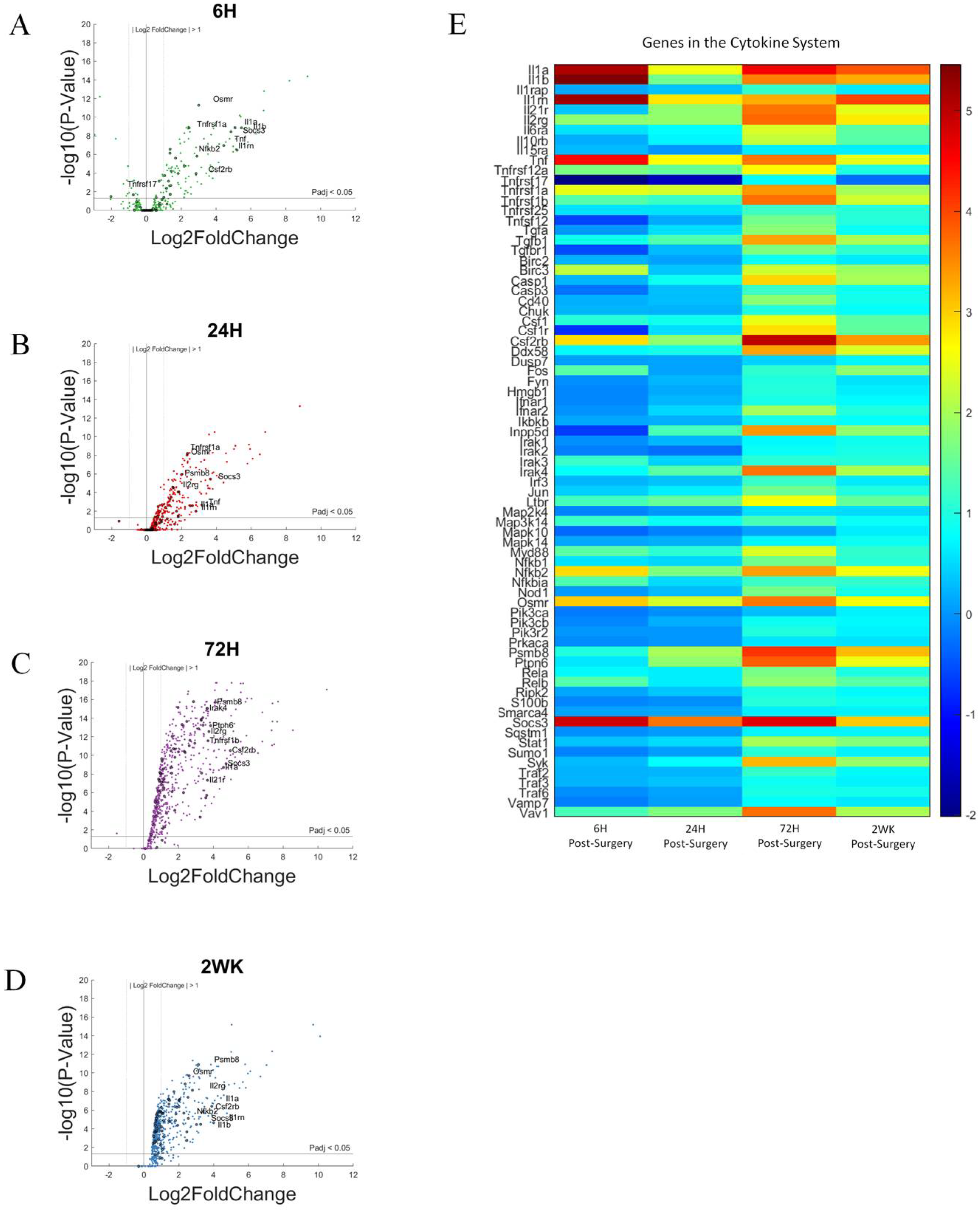
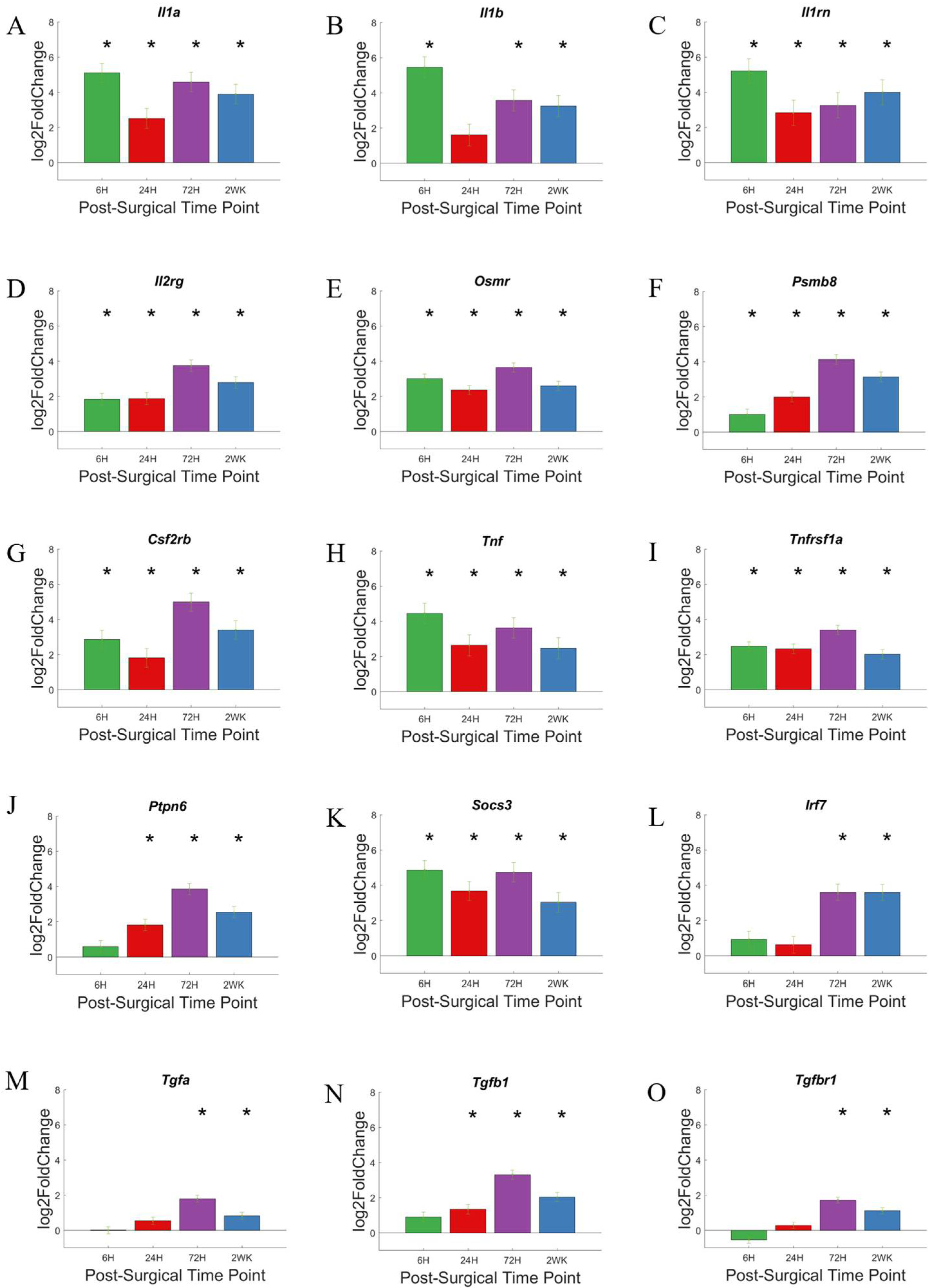
3.5. Cytokine Response
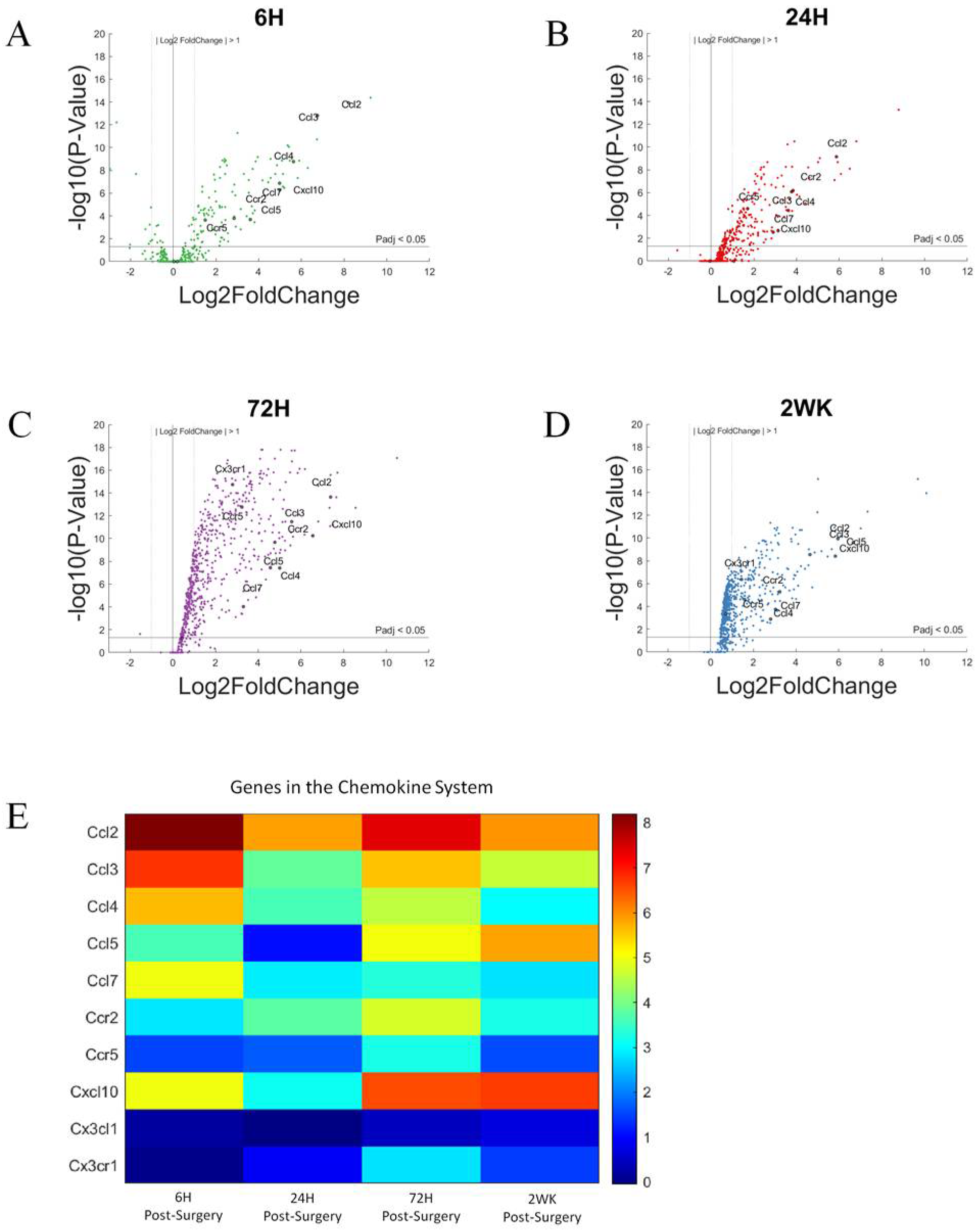
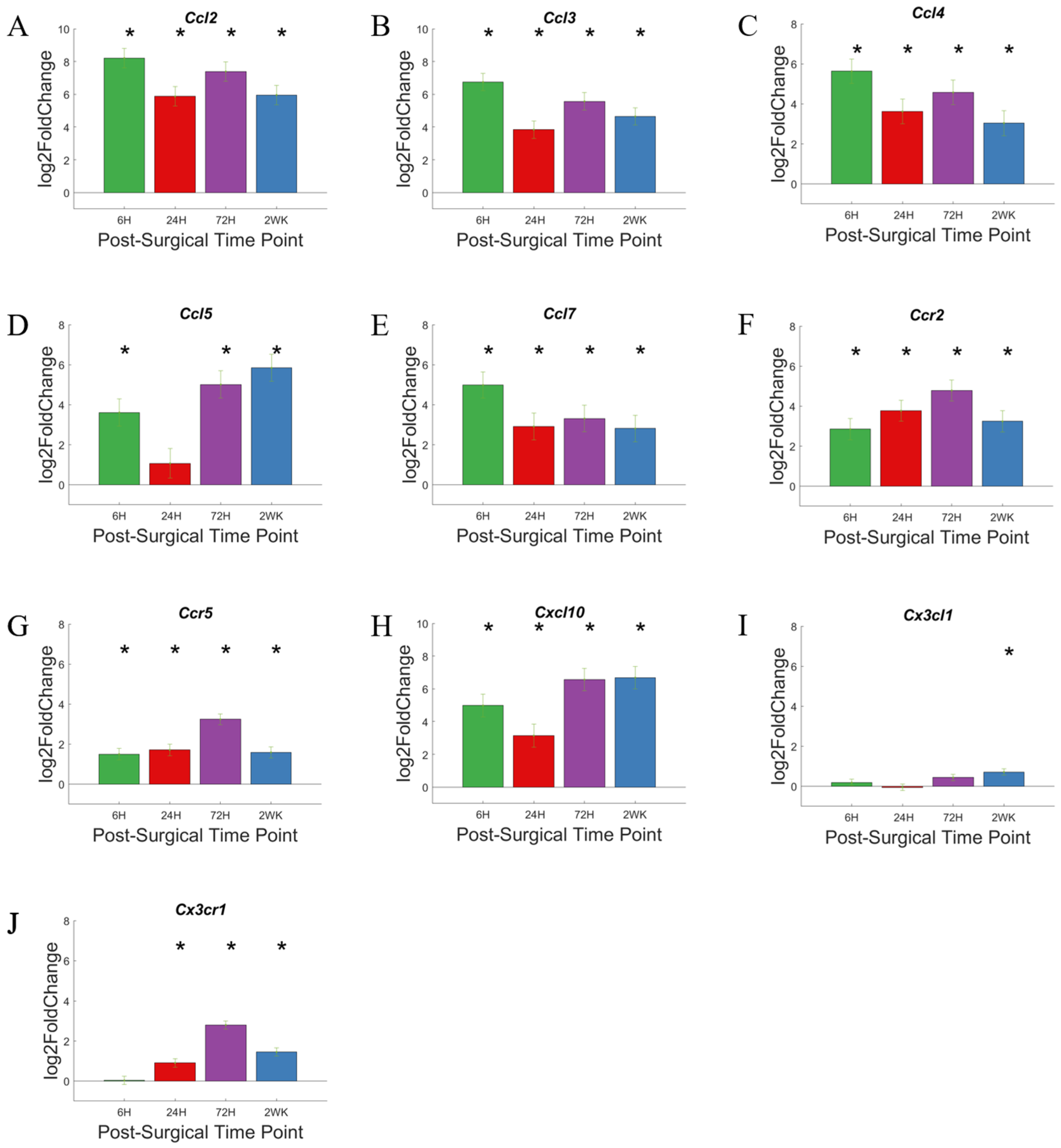
3.6. Chemokines
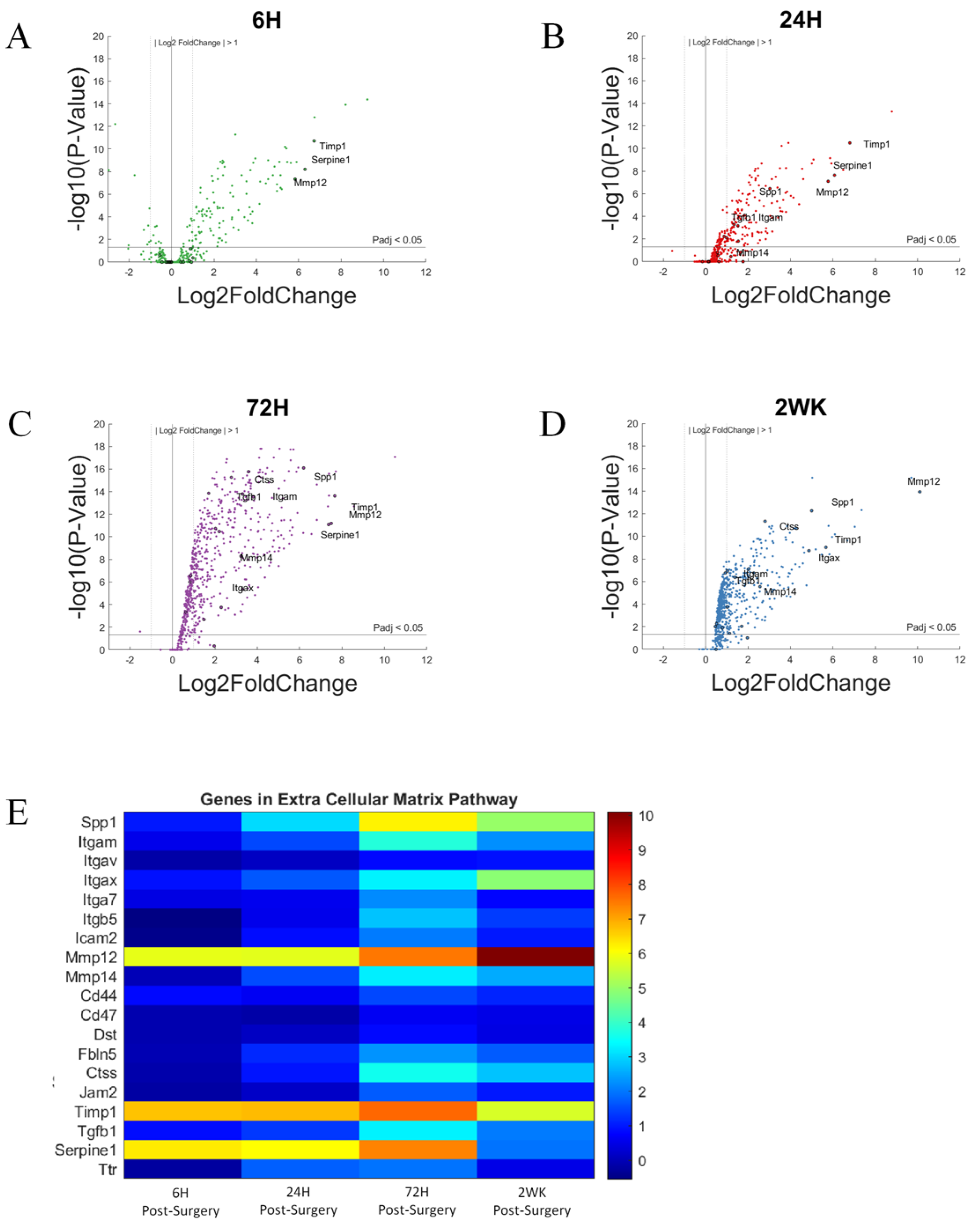
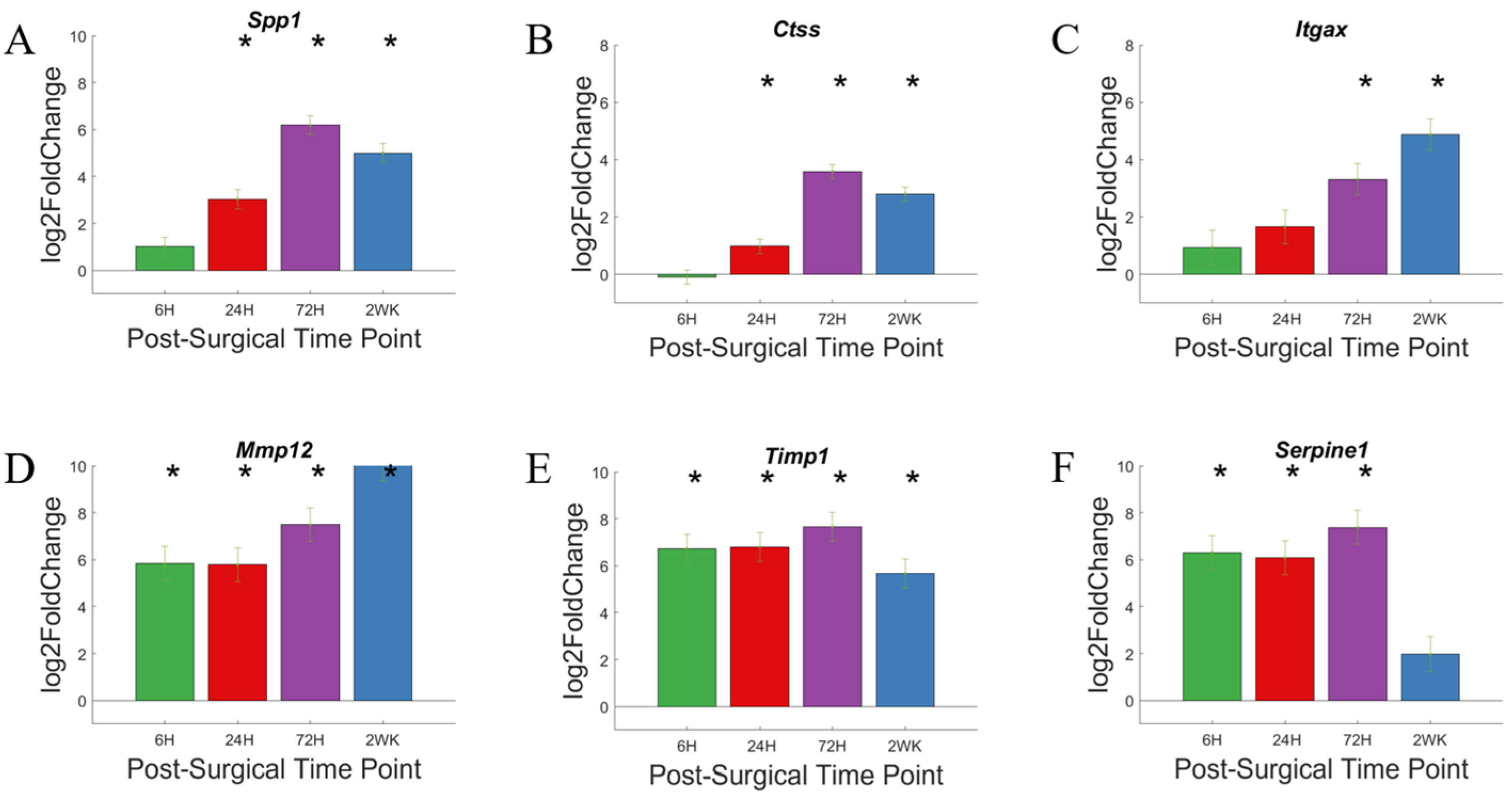
3.7. Extracellular Matrix
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Disclaimer
References
- Donoghue, J. Bridging the Brain to the World: A Perspective on Neural Interface Systems. Neuron 2008, 60, 511–521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicolelis, M.A. Brain-machine interfaces to restore motor function and probe neural circuits. Nat. Rev. Neurosci. 2003, 4, 417–422. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, A.B. Cortical neural prosthetics. Annu. Rev. Neurosci. 2004, 27, 487–507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hubel, D.H. Single unit activity in striate cortex of unrestrained cats. J. Physiol. 1959, 147, 226–238. [Google Scholar] [CrossRef]
- Yuste, R. From the neuron doctrine to neural networks. Nat. Rev. Neurosci. 2015, 16, 487–497. [Google Scholar] [CrossRef]
- Szostak, K.M.; Grand, L.; Constandinou, T.G. Neural Interfaces for Intracortical Recording: Requirements, Fabrication Methods, and Characteristics. Front. Neurosci. 2017, 11, 665. [Google Scholar] [CrossRef] [Green Version]
- Mak, J.N.; Wolpaw, J.R. Clinical Applications of Brain-Computer Interfaces: Current State and Future Prospects. IEEE Rev. Biomed. Eng. 2009, 2, 187–199. [Google Scholar] [CrossRef] [Green Version]
- McFarland, D.J.; Sarnacki, W.A.; Wolpaw, J.R. Electroencephalographic (EEG) control of three-dimensional movement. J. Neural Eng. 2010, 7, 036007. [Google Scholar] [CrossRef] [Green Version]
- Wolpaw, J.R.; McFarland, D.J. Control of a two-dimensional movement signal by a noninvasive brain-computer interface in humans. Proc. Natl. Acad. Sci. USA 2004, 101, 17849–17854. [Google Scholar] [CrossRef] [Green Version]
- Bell, C.J.; Shenoy, P.; Chalodhorn, R.; Rao, R.P. Control of a humanoid robot by a noninvasive brain-computer interface in humans. J. Neural Eng. 2008, 5, 214–220. [Google Scholar] [CrossRef]
- Hochberg, L.R.; Bacher, D.; Jarosiewicz, B.; Masse, N.Y.; Simeral, J.D.; Vogel, J.; Haddadin, S.; Liu, J.; Cash, S.S.; van der Smagt, P.; et al. Reach and grasp by people with tetraplegia using a neurally controlled robotic arm. Nature 2012, 485, 372–375. [Google Scholar] [CrossRef] [Green Version]
- Collinger, J.L.; Wodlinger, B.; Downey, J.E.; Wang, W.; Tyler-Kabara, E.C.; Weber, D.J.; McMorland, A.J.; Velliste, M.; Boninger, M.L.; Schwartz, A.B. High-performance neuroprosthetic control by an individual with tetraplegia. Lancet 2013, 381, 557–564. [Google Scholar] [CrossRef] [Green Version]
- Ajiboye, A.B.; Willett, F.R.; Young, D.R.; Memberg, W.D.; Murphy, B.A.; Miller, J.P.; Walter, B.L.; Sweet, J.A.; Hoyen, H.A.; Keith, M.W.; et al. Restoration of reaching and grasping movements through brain-controlled muscle stimulation in a person with tetraplegia: A proof-of-concept demonstration. Lancet 2017, 398, 1821–1830. [Google Scholar] [CrossRef] [Green Version]
- Barrese, J.C.; Rao, N.; Paroo, K.; Triebwasser, C.; Vargas-Irwin, C.; Franquemont, L.; Donoghue, J.P. Failure mode analysis of silicon-based intracortical microelectrode arrays in non-human primates. J. Neural Eng. 2013, 10, 066014. [Google Scholar] [CrossRef]
- Shoffstall, A.J.; Paiz, J.E.; Miller, D.M.; Rial, G.M.; Willis, M.T.; Menendez, D.M.; Hostler, S.R.; Capadona, J.R. Potential for thermal damage to the blood-brain barrier during craniotomy: Implications for intracortical recording microelectrodes. J. Neural Eng. 2018, 15, 034001. [Google Scholar] [CrossRef]
- Bjornsson, C.S.; Oh, S.J.; Al-Kofahi, Y.A.; Lim, Y.J.; Smith, K.L.; Turner, J.N.; De, S.; Roysam, B.; Shain, W.; Kim, S.J. Effects of insertion conditions on tissue strain and vascular damage during neuroprosthetic device insertion. J. Neural Eng. 2006, 3, 196–207. [Google Scholar] [CrossRef]
- Money, B.; Hess-Dunning, A.; Gloth, P.; Capadona, J.R.; Weder, C. Mechanically adaptive implants fabricated with poly(2-hydroxy¬ethyl methacrylate)-based negative photoresists. J. Mater. Chem. B 2020, 8, 6357–6365. [Google Scholar] [CrossRef]
- Bedell, H.W.; Song, S.; Li, X.; Molinich, E.; Lin, S.; Voit, W.E.; Pancrazio, J.J.; Capadona, J.R. Understanding the effects of both CD14-meditated innate immunity and device/tissue mechanical mismatch in the neuroinflammatory response to intracortical microelectrodes. Front. Neurosci. 2018, 12, 772. [Google Scholar] [CrossRef]
- Nguyen, J.K.; Jorfi, M.; Buchanan, K.L.; Park, D.J.; Foster, E.J.; Tyler, D.J.; Rowan, S.J.; Weder, C.; Capadona, J.R. Influence of resveratrol release on the tissue response to mechanically adaptive cortical implants. Acta Biomater. 2016, 29, 81–93. [Google Scholar] [CrossRef] [Green Version]
- Sridharan, A.; Nguyen, J.K.; Capadona, J.R.; Muthuswamy, J. Compliant intracortical implants reduce strains and strain rates in brain tissue in vivo. J. Neural Eng. 2015, 12, 036002. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, J.K.; Park, D.J.; Skousen, J.L.; Hess-Dunning, A.; Tyler, D.J.; Rowan, S.J.; Weder, C.; Capadona, J.R. Mechanically-compliant intracortical implants reduce the neuroinflammatory response. J. Neural Eng. 2014, 11, 056014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, J.P.; Capadona, J.R.; Miller, R.H.; Healy, B.C.; Shanmuganathan, K.; Rowan, S.J.; Weder, C.; Tyler, D.J. Mechanically adaptive intracortical implants improve the proximity of neuronal cell bodies. J. Neural Eng. 2011, 8, 066011. [Google Scholar] [CrossRef] [PubMed]
- Simon, D.M.; Charkhkar, H.; St John, C.; Rajendran, S.; Kang, T.; Reit, R.; Arreaga-Salas, D.; McHail, D.G.; Knaack, G.L.; Sloan, A.; et al. Design and demonstration of an intracortical probe technology with tunable modulus. J. Biomed. Mater. Res. A 2017, 105, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Ware, T.; Simon, D.; Arreaga-Salas, D.E.; Reeder, J.; Rennaker, R.; Keefer, E.W.; Voit, W. Fabrication of responsive, softening neural interfaces. Adv. Funct. Mater. 2012, 22, 3470–3479. [Google Scholar] [CrossRef]
- Welle, E.J.; Patel, P.R.; Woods, J.E.; Petrossians, A.; Della Valle, E.; Vega-Medina, A.; Richie, J.M.; Cai, D.; Weiland, J.D.; Chestek, C.A. Ultra-small carbon fiber electrode recording site optimization and improved in vivo chronic recording yield. J. Neural Eng. 2020, 17, 026037. [Google Scholar] [CrossRef]
- Ravikumar, M.; Hageman, D.J.; Tomaszewski, W.H.; Chandra, G.M.; Skousen, J.L.; Capadona, J.R. The effect of residual endotoxin contamination on the neuroinflammatory response to sterilized intracortical microelectrodes. J. Mater. Chem. B 2014, 2, 2517–2529. [Google Scholar] [CrossRef] [Green Version]
- Shoffstall, A.J.; Ecker, M.; Danda, V.; Joshi-Imre, A.; Stiller, A.; Yu, M.; Paiz, J.E.; Mancuso, E.; Bedell, H.W.; Voit, W.E.; et al. Characterization of the Neuroinflammatory Response to Thiol-ene Shape Memory Polymer Coated Intracortical Microelectrodes. Micromachines 2018, 9, 486. [Google Scholar] [CrossRef] [Green Version]
- Ereifej, E.S.; Rial, G.; Hermann, J.K.; Smith, C.S.; Meade, S.; Rayyan, J.; Chen, K.; Feng, H.; Capadona, J.R. Implantation of Neural Probes in the Brain Elicits Oxidative Stress. Front. Bioeng. Biotechnol. 2018, 6, 1–12. [Google Scholar] [CrossRef]
- Potter, K.A.; Buck, A.C.; Self, W.K.; Callanan, M.E.; Sunil, S.; Capadona, J.R. The effect of resveratrol on neurodegeneration and blood brain barrier stability surrounding intracortical. microelectrodes. Biomaterials 2013, 34, 7001–7015. [Google Scholar] [CrossRef]
- Potter, K.A.; Jorfi, M.; Householder, K.T.; Foster, E.J.; Weder, C.; Capadona, J.R. Curcumin-releasing mechanically-adaptive intracortical implants improve the proximal neuronal density and blood-brain barrier stability. Acta Biomater. 2014, 10, 2209–2222. [Google Scholar] [CrossRef]
- Potter-Baker, K.A.; Capadona, J.R. Reducing the “Stress”: Antioxidative Therapeutic and Material Approaches May Prevent Intracortical Microelectrode Failure. ACS Macro Lett. 2015, 4, 275–279. [Google Scholar] [CrossRef]
- Potter-Baker, K.A.; Nguyen, J.K.; Kovach, K.M.; Gitomer, M.M.; Srail, T.W.; Stewart, W.G.; Skousen, J.L.; Capadona, J.R. Development of Superoxide Dismutase Mimetic Surfaces to Reduce Accumulation of Reactive Oxygen Species Surrounding Intracortical Microelectrodes. J. Mater. Chem. B 2014, 2, 2248–2258. [Google Scholar] [CrossRef] [Green Version]
- Potter-Baker, K.A.; Stewart, W.G.; Tomaszewski, W.H.; Wong, C.T.; Meador, W.D.; Ziats, N.P.; Capadona, J.R. Implications of chronic daily anti-oxidant administration on the inflammatory response to intracortical microelectrodes. J. Neural Eng. 2015, 12, 046002. [Google Scholar] [CrossRef] [Green Version]
- Bennett, C.; Mohammed, F.; Alvarez-Ciara, A.; Nguyen, M.A.; Dietrich, W.D.; Rajguru, S.M.; Streit, W.J.; Prasad, A. Neuroinflammation, oxidative stress, and blood-brain barrier (BBB) disruption in acute Utah electrode array implants and the effect of deferoxamine as an iron chelator on acute foreign body response. Biomaterials 2019, 188, 144–159. [Google Scholar] [CrossRef]
- Ereifej, E.S.; Smith, C.S.; Meade, S.M.; Chen, K.; Feng, H.; Capadona, J.R. The neuroinflammatory response to nanopatterning parallel grooves into the surface structure of intracortical microelectrodes. Adv. Funct. Mater. 2018, 28, 1704420. [Google Scholar] [CrossRef]
- Bedell, H.W.; Schaub, N.J.; Capadona, J.R.; Ereifej, E.S. Differential expression of genes involved in the acute innate immune response to intracortical microelectrodes. Acta Biomater. 2020, 102, 205–219. [Google Scholar] [CrossRef]
- Bedell, H.W.; Hermann, J.K.; Ravikumar, M.; Lin, S.; Rein, A.; Li, X.; Molinich, E.; Smith, P.; Sidik, S.; Taylor, D.M.; et al. Targeting CD14 on blood derived cells improves intracortical microelectrode performance. Biomaterials 2018, 163, 163–173. [Google Scholar] [CrossRef]
- Jersmann, H.P. Time to abandon dogma: CD14 is expressed by non-myeloid lineage cells. Immunol. Cell Biol. 2005, 83, 462–467. [Google Scholar] [CrossRef]
- Haziot, A.; Chen, S.; Ferrero, E.; Low, M.G.; Silber, R.; Goyert, S.M. The monocyte differentiation antigen, CD14, is anchored to the cell membrane by a phosphatidylinositol linkage. J. Immunol. 1988, 141, 547–552. [Google Scholar]
- Tarassishin, L.; Suh, H.S.; Lee, S.C. LPS and IL-1 differentially activate mouse and human astrocytes: Role of CD14. Glia 2014, 62, 999–1013. [Google Scholar] [CrossRef] [Green Version]
- Widera, D.; Martinez Aguilar, R.; Cottrell, G.S. Toll-like receptor 4 and protease-activated receptor 2 in physiology and pathophysiology of the nervous system: More than just receptor cooperation? Neural Regen. Res. 2019, 14, 1196–1201. [Google Scholar] [CrossRef] [PubMed]
- Hermann, J.K.; Ravikumar, M.; Shoffstall, A.J.; Ereifej, E.S.; Kovach, K.M.; Chang, J.; Soffer, A.; Wong, C.; Srivastava, V.; Smith, P.; et al. Inhibition of the cluster of differentiation 14 innate immunity pathway with IAXO-101 improves chronic microelectrode performance. J. Neural Eng. 2018, 15, 025002. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, M.M. Digital Multiplexed Gene Expression Analysis Using the NanoString nCounter System. Curr. Protoc. Mol. Biol. 2011, 94, 25B-10. [Google Scholar]
- Gene Expression Data Analysis Guidelines. Available online: https://nanostring.com/wp-content/uploads/Gene_Expression_Data_Analysis_Guidelines.pdf (accessed on 20 June 2022).
- nSolver™ 4.0 Analysis Software User Manual. Available online: https://nanostring.com/wp-content/uploads/MAN-C0019-08_nSolver_4.0_Analysis_Software_User_Manual.pdf (accessed on 20 June 2022).
- NanoString Technologies®. Inc. nCounter Advanced Analysis 2.0 Plugin for nSolver Software. Available online: https://nanostring.com/wp-content/uploads/MAN-10030-03_nCounter_Advanced_Analysis_2.0_User_Manual.pdf (accessed on 20 June 2022).
- Brown, E.A.; Lautz, J.D.; Davis, T.R.; Gniffke, E.P.; VanSchoiack, A.A.W.; Neier, S.C.; Tashbook, N.; Nicolini, C.; Fahnestock, M.; Schrum, A.G.; et al. Clustering the autisms using glutamate synapse protein interaction networks from cortical and hippocampal tissue of seven mouse models. Mol. Autism 2018, 9, 48. [Google Scholar] [CrossRef]
- Sarkozy, C.; Chong, L.; Takata, K.; Chavez, E.A.; Miyata-Takata, T.; Duns, G.; Telenius, A.; Boyle, M.; Slack, G.W.; Laurent, C.; et al. Gene expression profiling of gray zone lymphoma. Blood Adv. 2020, 4, 2523–2535. [Google Scholar] [CrossRef]
- Metsalu, T.; Vilo, J. ClustVis: A web tool for visualizing clustering of multivariate data using Principal Component Analysis and heatmap. Nucleic Acids Res. 2015, 43, W566–W570. [Google Scholar] [CrossRef]
- Falcone, J.D.; Carroll, S.L.; Saxena, T.; Mandavia, D.; Clark, A.; Yarabarla, V.; Bellamkonda, R.V. Correlation of mRNA Expression and Signal Variability in Chronic Intracortical Electrodes. Front. Bioeng. Biotechnol. 2018, 6, 26. [Google Scholar] [CrossRef] [Green Version]
- Thompson, C.H.; Saxena, A.; Heelan, N.; Salatino, J.; Purcell, E.K. Spatiotemporal patterns of gene expression around implanted silicon electrode arrays. J. Neural Eng. 2021, 18, 045005. [Google Scholar] [CrossRef]
- Song, S.G.; Kim, S.; Koh, J.; Yim, J.; Han, B.; Kim, Y.A.; Jeon, Y.K.; Chung, D.H. Comparative analysis of the tumor immune-microenvironment of primary and brain metastases of non-small-cell lung cancer reveals organ-specific and EGFR mutation-dependent unique immune landscape. Cancer Immunol. Immunother. 2021, 70, 2035–2048. [Google Scholar] [CrossRef]
- Anderson, J.M.; Rodriguez, A.; Chang, D.T. Foreign body reaction to biomaterials. Semin. Immunol. 2008, 20, 86–100. [Google Scholar] [CrossRef] [Green Version]
- Park, J.E.; Barbul, A. Understanding the role of immune regulation in wound healing. Am. J. Surg. 2004, 187, 11S–16S. [Google Scholar] [CrossRef]
- Lempka, S.F.; Miocinovic, S.; Johnson, M.D.; Vitek, J.L.; McIntyre, C.C. In vivo impedance spectroscopy of deep brain stimulation electrodes. J. Neural Eng. 2009, 6, 046001. [Google Scholar] [CrossRef] [Green Version]
- Orsini, F.; De Blasio, D.; Zangari, R.; Zanier, E.R.; De Simoni, M.G. Versatility of the complement system in neuroinflammation, neurodegeneration and brain homeostasis. Front. Cell. Neurosci. 2014, 8, 380. [Google Scholar] [CrossRef] [Green Version]
- Garcia, B.L.; Zwarthoff, S.A.; Rooijakkers, S.H.; Geisbrecht, B.V. Novel Evasion Mechanisms of the Classical Complement Pathway. J. Immunol. 2016, 197, 2051–2060. [Google Scholar] [CrossRef]
- Lubbers, R.; van Essen, M.F.; van Kooten, C.; Trouw, L.A. Production of complement components by cells of the immune system. Clin. Exp. Immunol. 2017, 188, 183–194. [Google Scholar] [CrossRef] [Green Version]
- Mathern, D.R.; Heeger, P.S. Molecules Great and Small: The Complement System. Clin. J. Am. Soc. Nephrol. 2015, 10, 1636–1650. [Google Scholar] [CrossRef] [Green Version]
- Moghimi, S.M.; Andersen, A.J.; Ahmadvand, D.; Wibroe, P.P.; Andresen, T.L.; Hunter, A.C. Material properties in complement activation. Adv. Drug Deliv. Rev. 2011, 63, 1000–1007. [Google Scholar] [CrossRef]
- Arima, Y.; Toda, M.; Iwata, H. Surface plasmon resonance in monitoring of complement activation on biomaterials. Adv. Drug Deliv. Rev. 2011, 63, 988–999. [Google Scholar] [CrossRef] [Green Version]
- Bergseth, G.; Lambris, J.D.; Mollnes, T.E.; Lappegard, K.T. Artificial surface-induced inflammation relies on complement factor 5: Proof from a deficient person. Ann. Thorac. Surg. 2011, 91, 527–533. [Google Scholar] [CrossRef] [Green Version]
- Nilsson, B.; Ekdahl, K.N.; Mollnes, T.E.; Lambris, J.D. The role of complement in biomaterial-induced inflammation. Mol. Immunol. 2007, 44, 82–94. [Google Scholar] [CrossRef]
- Tang, L.; Liu, L.; Elwing, H.B. Complement activation and inflammation triggered by model biomaterial surfaces. J. Biomed. Mater. Res. 1998, 41, 333–340. [Google Scholar] [CrossRef]
- Ekdahl, K.N.; Lambris, J.D.; Elwing, H.; Ricklin, D.; Nilsson, P.H.; Teramura, Y.; Nicholls, I.A.; Nilsson, B. Innate immunity activation on biomaterial surfaces: A mechanistic model and coping strategies. Adv. Drug Deliv. Rev. 2011, 63, 1042–1050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lappegard, K.T.; Fung, M.; Bergseth, G.; Riesenfeld, J.; Lambris, J.D.; Videm, V.; Mollnes, T.E. Effect of complement inhibition and heparin coating on artificial surface-induced leukocyte and platelet activation. Ann. Thorac. Surg. 2004, 77, 932–941. [Google Scholar] [CrossRef]
- Lappegard, K.T.; Fung, M.; Bergseth, G.; Riesenfeld, J.; Mollnes, T.E. Artificial surface-induced cytokine synthesis: Effect of heparin coating and complement inhibition. Ann. Thorac. Surg. 2004, 78, 38–44; discussion 35–44. [Google Scholar] [CrossRef]
- Watkins, N.J.; Braidley, P.; Bray, C.J.; Savill, C.M.; White, D.J. Coating of human decay accelerating factor (hDAF) onto medical devices to improve biocompatibility. Immunopharmacology 1997, 38, 111–118. [Google Scholar] [CrossRef]
- Kumar, H.; Kawai, T.; Akira, S. Pathogen recognition by the innate immune system. Int. Rev. Immunol. 2011, 30, 16–34. [Google Scholar] [CrossRef]
- Lupfer, C.R.; Anand, P.K.; Qi, X.; Zaki, H. Editorial: Role of NOD-Like Receptors in Infectious and Immunological Diseases. Front. Immunol. 2020, 11, 923. [Google Scholar] [CrossRef]
- Nakaya, Y.; Lilue, J.; Stavrou, S.; Moran, E.A.; Ross, S.R. AIM2-Like Receptors Positively and Negatively Regulate the Interferon Response Induced by Cytosolic DNA. mBio 2017, 8, e00944-17. [Google Scholar] [CrossRef] [Green Version]
- Fekete, T.; Bencze, D.; Szabo, A.; Csoma, E.; Biro, T.; Bacsi, A.; Pazmandi, K. Regulatory NLRs Control the RLR-Mediated Type I Interferon and Inflammatory Responses in Human Dendritic Cells. Front. Immunol. 2018, 9, 2314. [Google Scholar] [CrossRef] [Green Version]
- Geijtenbeek, T.B.; Gringhuis, S.I. Signalling through C-type lectin receptors: Shaping immune responses. Nat. Rev. Immunol. 2009, 9, 465–479. [Google Scholar] [CrossRef]
- Arumugam, T.V.; Manzanero, S.; Furtado, M.; Biggins, P.J.; Hsieh, Y.H.; Gelderblom, M.; MacDonald, K.P.; Salimova, E.; Li, Y.I.; Korn, O.; et al. An atypical role for the myeloid receptor Mincle in central nervous system injury. J. Cereb. Blood Flow Metab. 2017, 37, 2098–2111. [Google Scholar] [CrossRef] [Green Version]
- Fitzgerald, K.A.; Kagan, J.C. Toll-like Receptors and the Control of Immunity. Cell 2020, 180, 1044–1066. [Google Scholar] [CrossRef]
- Hermann, J.K.; Lin, S.; Soffer, A.; Wong, C.; Srivastava, V.; Chang, J.; Sunil, S.; Sudhakar, S.; Tomaszewski, W.H.; Protasiewicz, G.; et al. The Role of Toll-Like Receptor 2 and 4 Innate Immunity Pathways in Intracortical Microelectrode-Induced Neuroinflammation. Front. Bioeng. Biotechnol. 2018, 6, 113. [Google Scholar] [CrossRef]
- Dinarello, C.A. Historical insights into cytokines. Eur. J. Immunol. 2007, 37 (Suppl. S1), S34–S45. [Google Scholar] [CrossRef] [Green Version]
- Jorfi, M.; Skousen, J.L.; Weder, C.; Capadona, J.R. Progress towards biocompatible intracortical microelectrodes for neural interfacing applications. J. Neural Eng. 2015, 12, 011001. [Google Scholar] [CrossRef]
- Kaplani, K.; Koutsi, S.; Armenis, V.; Skondra, F.G.; Karantzelis, N.; Champeris Tsaniras, S.; Taraviras, S. Wound healing related agents: Ongoing research and perspectives. Adv. Drug Deliv. Rev. 2018, 129, 242–253. [Google Scholar] [CrossRef]
- Hughes, C.E.; Nibbs, R.J.B. A guide to chemokines and their receptors. FEBS J. 2018, 285, 2944–2971. [Google Scholar] [CrossRef]
- Miller, M.C.; Mayo, K.H. Chemokines from a Structural Perspective. Int. J. Mol. Sci. 2017, 18, 2088. [Google Scholar] [CrossRef] [Green Version]
- White, G.E.; Iqbal, A.J.; Greaves, D.R. CC chemokine receptors and chronic inflammation—Therapeutic opportunities and pharmacological challenges. Pharmacol. Rev. 2013, 65, 47–89. [Google Scholar] [CrossRef] [Green Version]
- Antonelli, A.; Ferrari, S.M.; Giuggioli, D.; Ferrannini, E.; Ferri, C.; Fallahi, P. Chemokine (C-X-C motif) ligand (CXCL)10 in autoimmune diseases. Autoimmun. Rev. 2014, 13, 272–280. [Google Scholar] [CrossRef]
- Han, X. Constitutively active chemokine CXC receptors. Adv. Pharmacol. 2014, 70, 265–301. [Google Scholar] [CrossRef]
- Song, I.; Dityatev, A. Crosstalk between glia, extracellular matrix and neurons. Brain Res. Bull. 2018, 136, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Krishnaswamy, V.R.; Benbenishty, A.; Blinder, P.; Sagi, I. Demystifying the extracellular matrix and its proteolytic remodeling in the brain: Structural and functional insights. Cell. Mol. Life Sci. 2019, 76, 3229–3248. [Google Scholar] [CrossRef] [PubMed]
- Morawski, M.; Filippov, M.; Tzinia, A.; Tsilibary, E.; Vargova, L. ECM in brain aging and dementia. Prog. Brain Res. 2014, 214, 207–227. [Google Scholar] [CrossRef] [PubMed]
- Tsien, R.Y. Very long-term memories may be stored in the pattern of holes in the perineuronal net. Proc. Natl. Acad. Sci. USA 2013, 110, 12456–12461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsilibary, E.; Tzinia, A.; Radenovic, L.; Stamenkovic, V.; Lebitko, T.; Mucha, M.; Pawlak, R.; Frischknecht, R.; Kaczmarek, L. Neural ECM proteases in learning and synaptic plasticity. Prog. Brain Res. 2014, 214, 135–157. [Google Scholar] [CrossRef] [PubMed]
- Soleman, S.; Filippov, M.A.; Dityatev, A.; Fawcett, J.W. Targeting the neural extracellular matrix in neurological disorders. Neuroscience 2013, 253, 194–213. [Google Scholar] [CrossRef]
- Jang, D.G.; Sim, H.J.; Song, E.K.; Kwon, T.; Park, T.J. Extracellular matrixes and neuroinflammation. BMB Rep. 2020, 53, 491–499. [Google Scholar] [CrossRef]
- Suttkus, A.; Morawski, M.; Arendt, T. Protective Properties of Neural Extracellular Matrix. Mol. Neurobiol. 2016, 53, 73–82. [Google Scholar] [CrossRef]
- Chelluboina, B.; Nalamolu, K.R.; Klopfenstein, J.D.; Pinson, D.M.; Wang, D.Z.; Vemuganti, R.; Veeravalli, K.K. MMP-12, a Promising Therapeutic Target for Neurological Diseases. Mol. Neurobiol. 2018, 55, 1405–1409. [Google Scholar] [CrossRef]
- Hahn, J.N.; Kaushik, D.K.; Mishra, M.K.; Wang, J.; Silva, C.; Yong, V.W. Impact of Minocycline on Extracellular Matrix Metalloproteinase Inducer, a Factor Implicated in Multiple Sclerosis Immunopathogenesis. J. Immunol. 2016, 197, 3850–3860. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, H.; Sugimura, Y.; Iwama, S.; Suzuki, H.; Nobuaki, O.; Nagasaki, H.; Arima, H.; Sawada, M.; Oiso, Y. Minocycline prevents osmotic demyelination syndrome by inhibiting the activation of microglia. J. Am. Soc. Nephrol. 2010, 21, 2090–2098. [Google Scholar] [CrossRef] [Green Version]
- Rennaker, R.L.; Miller, J.; Tang, H.; Wilson, D.A. Minocycline increases quality and longevity of chronic neural recordings. J. Neural Eng. 2007, 4, L1–L5. [Google Scholar] [CrossRef] [Green Version]
- Moore, C.S.; Crocker, S.J. An alternate perspective on the roles of TIMPs and MMPs in pathology. Am. J. Pathol. 2012, 180, 12–16. [Google Scholar] [CrossRef]
- Gardner, J.; Ghorpade, A. Tissue inhibitor of metalloproteinase (TIMP)-1: The TIMPed balance of matrix metalloproteinases in the central nervous system. J. Neurosci. Res. 2003, 74, 801–806. [Google Scholar] [CrossRef] [Green Version]
- Lin, M.; Griessenauer, C.J.; Starke, R.M.; Tubbs, R.S.; Shoja, M.M.; Foreman, P.M.; Vyas, N.A.; Walters, B.C.; Harrigan, M.R.; Hendrix, P.; et al. Haplotype analysis of SERPINE1 gene: Risk for aneurysmal subarachnoid hemorrhage and clinical outcomes. Mol. Genet. Genom. Med. 2019, 7, e737. [Google Scholar] [CrossRef] [Green Version]
- Bonneh-Barkay, D.; Wiley, C.A. Brain extracellular matrix in neurodegeneration. Brain Pathol. 2009, 19, 573–585. [Google Scholar] [CrossRef]
- Gur-Wahnon, D.; Mizrachi, T.; Wald-Altman, S.; Al-Roof Higazi, A.; Brenner, T. Tissue plasminogen activator involvement in experimental autoimmune myasthenia gravis: Aggravation and therapeutic potential. J. Autoimmun. 2014, 52, 36–43. [Google Scholar] [CrossRef]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Abcc3 | Bnip3l | Cdc7 | Dock2 | Gfap | Il1rl2 | Lcn2 | Myc | Plcg2 | Ripk1 | Spib | Topbp1 |
| Abcc8 | Bok | Cdk20 | Dot1l | Gja1 | Il1rn | Ldha | Myct1 | Pld1 | Ripk2 | Spint1 | Tpd52 |
| Abl1 | Bola2 | Cdkn1a | Dst | Gjb1 | Il21r | Ldlrad3 | Myd88 | Pld2 | Rnf8 | Spp1 | Tpsb2 |
| Adamts16 | Braf | Cdkn1c | Duoxa1 | Gna15 | Il2rg | Lfng | Myrf | Plekhb1 | Rpa1 | Sqstm1 | Tradd |
| Ago4 | Brca1 | Ceacam3 | Dusp7 | Gpr183 | Il3 | Lgmn | Nbn | Plekhm1 | Rpl28 | Srgn | Traf1 |
| Agt | Brd2 | Cflar | E2f1 | Gpr34 | Il36ra | Lig1 | Ncaph | Pllp | Rpl29 | Srxn1 | Traf2 |
| AI464131 | Brd3 | Ch25h | Eed | Gpr62 | Il3ra | Lilrb4a | Ncf1 | Plp1 | Rpl36al | St3gal6 | Traf3 |
| Aim2 | Brd4 | Chek1 | Eef2k | Gpr84 | Il6ra | Lingo1 | Ncor1 | Plxdc2 | Rpl9 | St8sia6 | Traf6 |
| Ak1 | Btk | Chek2 | Egfr | Grap | iNos | Lmna | Ncor2 | Plxnb3 | Rps10 | Stat1 | Trat1 |
| Akt1 | C1qa | Chn2 | Egr1 | Gria1 | Inpp5d | Lmnb1 | Ncr1 | Pmp22 | Rps2 | Steap4 | Trem1 |
| Akt2 | C1qb | Chst8 | Ehd2 | Gria2 | Iqsec1 | Lrg1 | Nefl | Pms2 | Rps21 | Stmn1 | Trem2 |
| Aldh1l1 | C1qc | Chuk | Ehmt2 | Gria4 | Irak1 | Lrrc25 | Nfe2l2 | Pnoc | Rps3 | Stx18 | Trem3 |
| Ambra1 | C3 | Cidea | Eif1 | Grin2a | Irak2 | Lrrc3 | Nfkb1 | Pole | Rps9 | Sumo1 | Trim47 |
| Amigo2 | C3ar1 | Cideb | Emcn | Grin2b | Irak3 | Lsr | Nfkb2 | Ppfia4 | Rrm2 | Suv39h1 | Trp53 |
| Anapc15 | C4a | Cks1b | Emp1 | Grm2 | Irak4 | Lst1 | Nfkbia | Ppp3ca | Rsad2 | Suv39h2 | Trp53bp2 |
| Anxa1 | C5ar1 | Clcf1 | eNos | Grm3 | Irf1 | Lta | Nfkbie | Ppp3cb | Rtn4rl1 | Suz12 | Trp73 |
| Apc | C6 | Cldn5 | Enpp6 | Grn | Irf2 | Ltb | Ngf | Ppp3r1 | S100a10 | Syk | Trpa1 |
| Apex1 | Cables1 | Clec7a | Entpd2 | Gsn | Irf3 | Ltbr | Ngfr | Ppp3r2 | S100b | Syn2 | Trpm4 |
| Apoe | Calcoco2 | Clic4 | Eomes | Gstm1 | Irf4 | Ltc4s | Ninj2 | Prdx1 | S1pr3 | Syp | Tspan18 |
| App | Calr | Cln3 | Ep300 | Gzma | Irf6 | Ly6a | Nkg7 | Prf1 | S1pr4 | Tarbp2 | Ttr |
| Aqp4 | Camk4 | Clstn1 | Epcam | Gzmb | Irf7 | Ly6g | Nlgn1 | Prkaca | S1pr5 | Tbc1d4 | Tubb3 |
| Arc | Casp1 | Cnn2 | Epg5 | H2afx | Irf8 | Ly9 | Nlgn2 | Prkacb | Sall1 | Tbr1 | Tubb4a |
| Arg1 | Casp2 | Cnp | Epsti1 | H2-T23 | Islr2 | Lyn | Nlrp2 | Prkar1a | Scd1 | Tbx21 | Txnrd1 |
| Arhgap24 | Casp3 | Cntnap2 | Erbb3 | Hat1 | Itga6 | Mafb | Nlrp3 | Prkar2a | Sell | Tcirg1 | Tyrobp |
| Arid1a | Casp4 | Coa5 | Ercc2 | Hcar2 | Itga7 | Maff | nNos | Prkar2b | Serpina3n | Tcl1 | Ugt8a |
| Asb2 | Casp6 | Col6a3 | Ercc6 | Hdac1 | Itgam | Mag | Nod1 | Prkce | Serpine1 | Tet1 | Ulk1 |
| Ash2l | Casp7 | Cotl1 | Esam | Hdac2 | Itgav | Mal | Nostrin | Prkcq | Serpinf1 | Tfg | Ung |
| Asph | Casp8 | Cox5b | Ets2 | Hdac4 | Itgax | Man2b1 | Noxa1 | Prkdc | Serping1 | Tgfa | Uty |
| Atf3 | Casp9 | Cp | Exo1 | Hdac6 | Itgb5 | Map1lc3a | Npl | Prnp | Sesn1 | Tgfb1 | Vamp7 |
| Atg14 | Cass4 | Cpa3 | Ezh1 | Hdc | Jag1 | Map2k1 | Npnt | Pros1 | Sesn2 | Tgfbr1 | Vav1 |
| Atg3 | Ccl2 | Creb1 | Ezh2 | Hells | Jam2 | Map2k4 | Nptx1 | Psen2 | Setd1a | Tgm1 | Vegfa |
| Atg5 | Ccl3 | Crebbp | F3 | Hif1a | Jarid2 | Map3k1 | Nqo1 | Psmb8 | Setd1b | Tgm2 | Vim |
| Atg7 | Ccl4 | Crem | Fa2h | Hilpda | Jun | Map3k14 | Nrgn | Pten | Setd2 | Tie1 | Vps4a |
| Atg9a | Ccl5 | Crip1 | Fabp5 | Hira | Kat2a | Mapk10 | Nrm | Ptger3 | Setd7 | Timeless | Vps4b |
| Atm | Ccl7 | Cryba4 | Fadd | Hist1h1d | Kat2b | Mapk12 | Nrp2 | Ptger4 | Setdb1 | Timp1 | Was |
| Atp6v0e | Ccng2 | Csf1 | Fancc | Hmgb1 | Kcnd1 | Mapk14 | Nthl1 | Ptgs2 | Sftpd | Tle3 | Wdr5 |
| Atp6v1a | Ccni | Csf1r | Fancd2 | Hmox1 | Kcnj10 | Mapt | Nwd1 | Ptms | Sh2d1a | Tlr2 | Xcl1 |
| Atr | Ccr2 | Csf2rb | Fancg | Homer1 | Kcnk13 | Marco | Oas1g | Ptpn6 | Shank3 | Tlr4 | Xiap |
| Axl | Ccr5 | Csf3r | Fas | Hpgds | Kdm1a | Mavs | Ogg1 | Ptprc | Siglec1 | Tlr7 | Xrcc6 |
| B3gnt5 | Cd109 | Csk | Fasl | Hprt | Kdm1b | Mb21d1 | Olfml3 | Pttg1 | Siglecf | Tm4sf1 | Zbp1 |
| Bad | Cd14 | Cst7 | Fbln5 | Hps4 | Kdm2a | Mbd2 | Opalin | Ptx3 | Sin3a | Tmc7 | Zfp367 |
| Bag3 | Cd163 | Ctse | Fcer1g | Hrk | Kdm2b | Mbd3 | Optn | Pycard | Sirt1 | Tmcc3 | Aars |
| Bag4 | Cd19 | Ctsf | Fcgr1 | Hsd11b1 | Kdm3a | Mcm2 | Osgin1 | Rab6b | Slamf8 | Tmem100 | Asb10 |
| Bak1 | Cd209e | Ctss | Fcgr2b | Hspb1 | Kdm3b | Mcm5 | Osmr | Rab7 | Slamf9 | Tmem119 | Ccdc127 |
| Bard1 | Cd244 | Ctsw | Fcgr3 | Hus1 | Kdm4a | Mcm6 | P2rx7 | Rac1 | Slc10a6 | Tmem144 | Cnot10 |
| Bax | Cd24a | Cx3cl1 | Fcrla | Icam2 | Kdm4b | Mdc1 | P2ry12 | Rac2 | Slc17a6 | Tmem173 | Csnk2a2 |
| Bbc3 | Cd300lf | Cx3cr1 | Fcrlb | Ifi30 | Kdm4c | Mdm2 | Pacsin1 | Rad1 | Slc17a7 | Tmem204 | Fam104a |
| Bcas1 | Cd33 | Cxcl10 | Fcrls | Ifih1 | Kdm4d | Mef2c | Padi2 | Rad17 | Slc1a3 | Tmem206 | Gusb |
| Bcl10 | Cd36 | Cxcl9 | Fdxr | Ifitm2 | Kdm5a | Mertk | Pak1 | Rad50 | Slc2a1 | Tmem37 | Lars |
| Bcl2 | Cd3d | Cycs | Fen1 | Ifitm3 | Kdm5b | Mfge8 | Parp1 | Rad51 | Slc2a5 | Tmem64 | Mto1 |
| Bcl2a1a | Cd3e | Cyp27a1 | Fgd2 | Ifnar1 | Kdm5c | Mgmt | Parp2 | Rad51b | Slc44a1 | Tmem88b | Supt7l |
| Bcl2l1 | Cd3g | Cyp7b1 | Fgf13 | Ifnar2 | Kdm5d | Mincle | Pcna | Rad51c | Slc6a1 | Tnf | Tada2b |
| Bcl2l11 | Cd40 | Cytip | Fgl2 | Igf1 | Kdm6a | Mmp12 | Pdpn | Rad9a | Slco2b1 | Tnfrsf10b | Tbp |
| Bcl2l2 | Cd44 | Dab2 | Fkbp5 | Igf1r | Kif2c | Mmp14 | Pecam1 | Rag1 | Slfn8 | Tnfrsf11b | Xpnpep1 |
| Bdnf | Cd47 | Dapk1 | Flt1 | Igf2r | Kir3dl1 | Mobp | Pex14 | Rage | Smarca4 | Tnfrsf12a | |
| Becn1 | Cd6 | Ddb2 | Fos | Igsf10 | Kir3dl2 | Mog | Pik3ca | Rala | Smarca5 | Tnfrsf13c | |
| Bid | Cd68 | Ddx58 | Foxp3 | Igsf6 | Kit | Mpeg1 | Pik3cb | Ralb | Smarcd1 | Tnfrsf17 | |
| Bik | Cd69 | Dicer1 | Fpr1 | Ikbkb | Klrb1 | Mpg | Pik3cd | Rapgef3 | Smc1a | Tnfrsf1a | |
| Bin1 | Cd70 | Dlg1 | Fscn1 | Ikbke | Klrd1 | Mr1 | Pik3cg | Rb1cc1 | Snca | Tnfrsf1b | |
| Birc2 | Cd72 | Dlg4 | Fyn | Ikbkg | Klrk1 | Mre11a | Pik3r1 | Rbfox3 | Socs3 | Tnfrsf25 | |
| Birc3 | Cd74 | Dlx1 | Gadd45a | Il10rb | Kmt2a | Ms4a1 | Pik3r2 | Rela | Sod1 | Tnfrsf4 | |
| Birc5 | Cd83 | Dlx2 | Gadd45g | Il15ra | Kmt2c | Ms4a2 | Pik3r5 | Relb | Sod2 | Tnfsf10 | |
| Blk | Cd84 | Dna2 | Gal3st1 | Il1a | Lacc1 | Ms4a4a | Pilra | Reln | Sod3 | Tnfsf12 | |
| Blm | Cd86 | Dnmt1 | Gba | Il1b | Lag3 | Msh2 | Pilrb1 | Reserved | Sox10 | Tnfsf13b | |
| Blnk | Cd8a | Dnmt3a | Gbp2 | Il1r1 | Lair1 | Msn | Pink1 | Rgl1 | Sox4 | Tnfsf4 | |
| Bmi1 | Cd8b1 | Dnmt3b | Gclc | Il1r2 | Lamp1 | Msr1 | Pla2g4a | Rhoa | Sox9 | Tnfsf8 | |
| Bnip3 | Cdc25a | Dock1 | Gdpd2 | Il1rap | Lamp2 | Mvp | Pla2g5 | Rig1 | Sphk1 | Top2a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, S.; Regan, B.; Ereifej, E.S.; Chan, E.R.; Capadona, J.R. Neuroinflammatory Gene Expression Analysis Reveals Pathways of Interest as Potential Targets to Improve the Recording Performance of Intracortical Microelectrodes. Cells 2022, 11, 2348. https://doi.org/10.3390/cells11152348
Song S, Regan B, Ereifej ES, Chan ER, Capadona JR. Neuroinflammatory Gene Expression Analysis Reveals Pathways of Interest as Potential Targets to Improve the Recording Performance of Intracortical Microelectrodes. Cells. 2022; 11(15):2348. https://doi.org/10.3390/cells11152348
Chicago/Turabian StyleSong, Sydney, Brianna Regan, Evon S. Ereifej, E. Ricky Chan, and Jeffrey R. Capadona. 2022. "Neuroinflammatory Gene Expression Analysis Reveals Pathways of Interest as Potential Targets to Improve the Recording Performance of Intracortical Microelectrodes" Cells 11, no. 15: 2348. https://doi.org/10.3390/cells11152348
APA StyleSong, S., Regan, B., Ereifej, E. S., Chan, E. R., & Capadona, J. R. (2022). Neuroinflammatory Gene Expression Analysis Reveals Pathways of Interest as Potential Targets to Improve the Recording Performance of Intracortical Microelectrodes. Cells, 11(15), 2348. https://doi.org/10.3390/cells11152348