
Links of Cytoskeletal Integrity with Disease and Aging


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
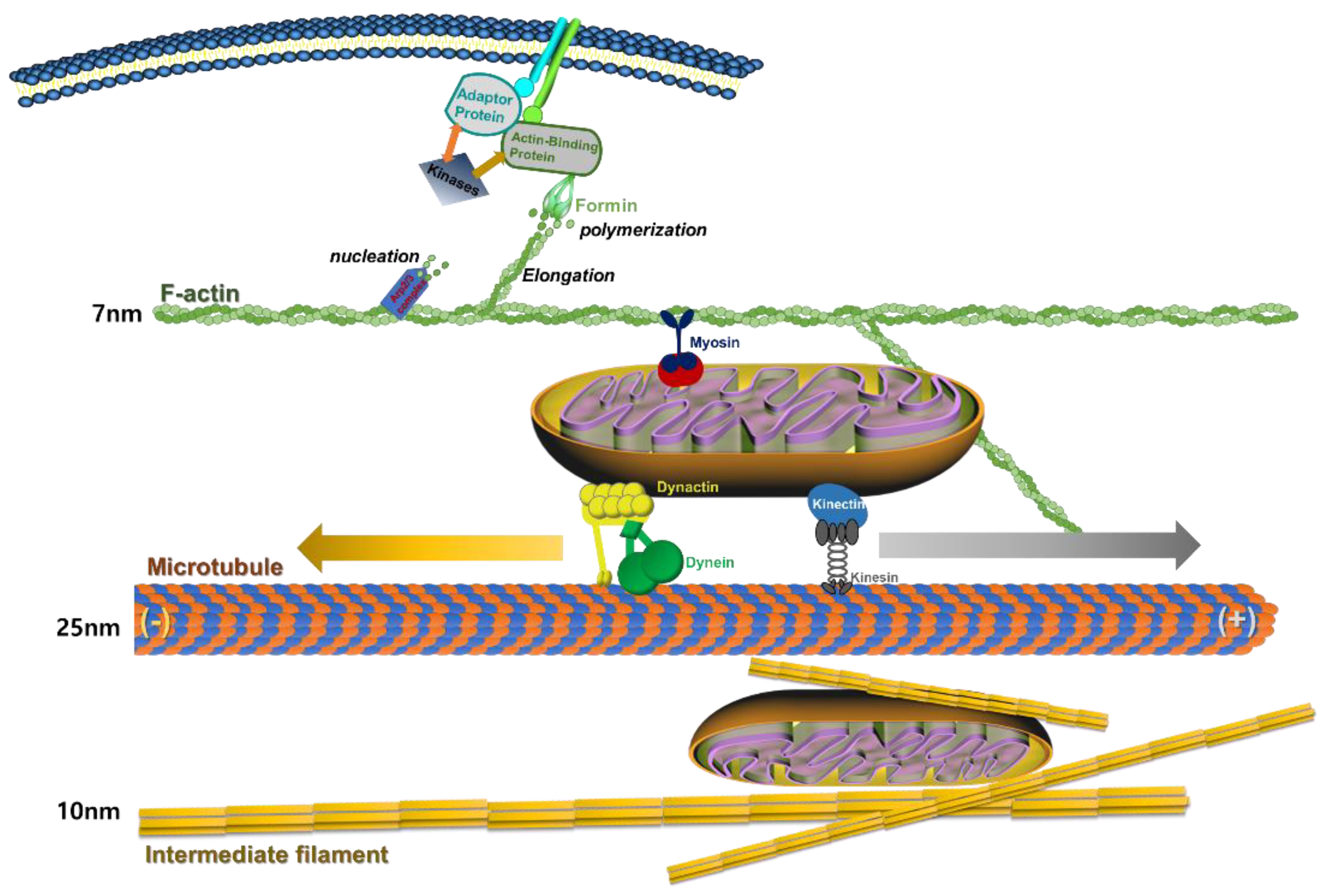
2. Description of the Cytoskeleton
2.1. Actin Cytoskeleton
2.2. Microfilaments
2.3. Microtubules
2.4. Intermediate Filaments
2.5. The Strutural Formation of Actin-Cytoskeleton and Mitochondrial Transfer
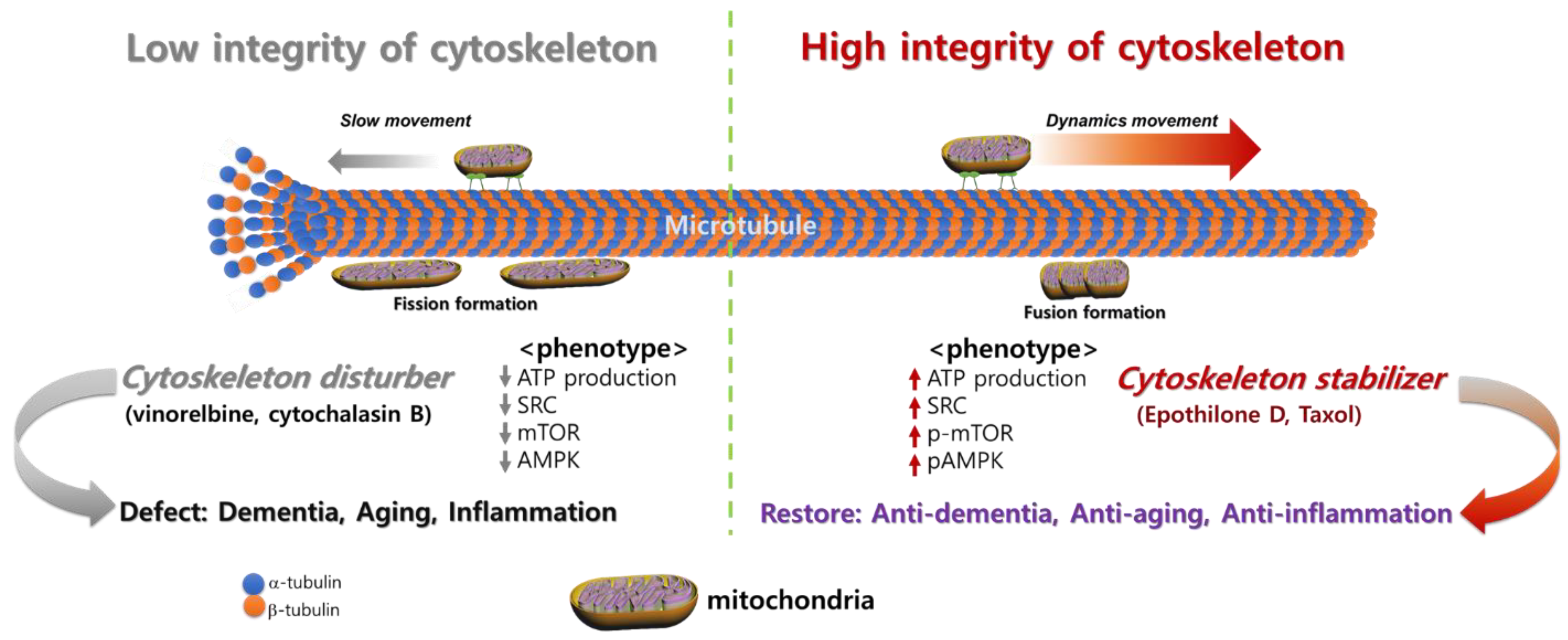
3. Cytoskeletal Integrity Changes with Disease and Aging
3.1. Altered Expression of Actin during Aging
3.2. Damage of Actin by Reactive Oxygen Species (ROS) and Aging
3.3. Alterations in Microtubule Regulation with Aging
3.4. Changes in Intermediate Filaments with Aging
4. Diseases Related to Cytoskeletal Integrity and Cytoskeleton-Targeting Therapy
4.1. Dementia
4.2. Diminished Reproductive Potential in Oocytes with Aging
4.3. COPD
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Boccardi, V. Population Ageing: The Need for a Care Revolution in a World 2.0. Geriatrics 2019, 4, 47. [Google Scholar] [CrossRef] [PubMed]
- Esser, D.E.; Ward, P.S. Ageing as a global public health challenge: From complexity reduction to aid effectiveness. Glob. Public Health 2013, 8, 745–768. [Google Scholar] [CrossRef] [PubMed]
- Jaul, E.; Barron, J. Age-Related Diseases and Clinical and Public Health Implications for the 85 Years Old and Over Population. Front. Public Health 2017, 5, 335. [Google Scholar] [CrossRef] [PubMed]
- Marks, P.W.; Hahn, S. Identifying the Risks of Unproven Regenerative Medicine Therapies. JAMA 2020, 324, 241–242. [Google Scholar] [CrossRef]
- Piscaglia, A.C. Stem cells, a two-edged sword: Risks and potentials of regenerative medicine. World J. Gastroenterol. 2008, 14, 4273–4279. [Google Scholar] [CrossRef]
- Ferraro, K.F.; Kemp, B.R.; Williams, M.M. Diverse Aging and Health Inequality by Race and Ethnicity. Innov. Aging 2017, 1. [Google Scholar] [CrossRef]
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The Hallmarks of Aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef]
- Shammas, M.A. Telomeres, lifestyle, cancer, and aging. Curr. Opin. Clin. Nutr. Metab. Care 2011, 14, 28–34. [Google Scholar] [CrossRef]
- Quazi, S. Telomerase gene therapy: A remission toward cancer. Med Oncol. 2022, 39, 1–20. [Google Scholar] [CrossRef]
- Hahn, A.; Zuryn, S. Mitochondrial Genome (mtDNA) Mutations that Generate Reactive Oxygen Species. Antioxidants 2019, 8, 392. [Google Scholar] [CrossRef] [Green Version]
- Payne, B.A.I.; Chinnery, P.F. Mitochondrial dysfunction in aging: Much progress but many unresolved questions. Biochim. Biophys. Acta 2015, 1847, 1347–1353. [Google Scholar] [CrossRef] [PubMed]
- Yu, R.; Lendahl, U.; Nistér, M.; Zhao, J. Regulation of Mammalian Mitochondrial Dynamics: Opportunities and Challenges. Front. Endocrinol. 2020, 11, 374. [Google Scholar] [CrossRef]
- Amberg, D.C.; Leadsham, J.E.; Kotiadis, V.; Gourlay, C.W. Cellular Ageing and the Actin Cytoskeleton. Aging Res. Yeast 2011, 57, 331–352. [Google Scholar] [CrossRef]
- Harris, M.J.; Wirtz, D.; Wu, P.-H. Dissecting cellular mechanics: Implications for aging, cancer, and immunity. Semin. Cell Dev. Biol. 2018, 93, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Lai, W.-F.; Wong, W.-T. Roles of the actin cytoskeleton in aging and age-associated diseases. Ageing Res. Rev. 2020, 58, 101021. [Google Scholar] [CrossRef] [PubMed]
- Gourlay, C.; Ayscough, K.R. The actin cytoskeleton: A key regulator of apoptosis and ageing? Nat. Rev. Mol. Cell Biol. 2005, 6, 583–589. [Google Scholar] [CrossRef]
- Cohen, A.A. Complex systems dynamics in aging: New evidence, continuing questions. Biogerontology 2015, 17, 205–220. [Google Scholar] [CrossRef]
- Pegoraro, A.F.; Janmey, P.; Weitz, D.A. Mechanical Properties of the Cytoskeleton and Cells. Cold Spring Harb. Perspect. Biol. 2017, 9, a022038. [Google Scholar] [CrossRef]
- Phillip, J.M.; Aifuwa, I.; Walston, J.; Wirtz, D. The Mechanobiology of Aging. Annu. Rev. Biomed. Eng. 2015, 17, 113–141. [Google Scholar] [CrossRef]
- Starodubtseva, M.N. Mechanical properties of cells and ageing. Ageing Res. Rev. 2011, 10, 16–25. [Google Scholar] [CrossRef]
- Zahn, J.T.; Louban, I.; Jungbauer, S.; Bissinger, M.; Kaufmann, D.; Kemkemer, R.; Spatz, J.P. Age-Dependent Changes in Microscale Stiffness and Mechanoresponses of Cells. Small 2011, 7, 1480–1487. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Dominguez, R. Regulation of actin cytoskeleton dynamics in cells. Mol. Cells 2010, 29, 311–325. [Google Scholar] [CrossRef] [PubMed]
- Bretscher, A.; Drees, B.; Harsay, E.; Schott, D.; Wang, T. What are the basic functions of microfilaments? Insights from studies in budding yeast. J. Cell Biol. 1994, 126, 821–825. [Google Scholar] [CrossRef] [PubMed]
- Dos Remedios, C.G.; Chhabra, D.; Kekic, M.; Dedova, I.V.; Tsubakihara, M.; Berry, D.A.; Nosworthy, N.J. Actin Binding Proteins: Regulation of Cytoskeletal Microfilaments. Physiol. Rev. 2003, 83, 433–473. [Google Scholar] [CrossRef] [PubMed]
- Kitakaze, M. Dysfunction of microtubules induces cardiac dysfunction. eBioMedicine 2018, 37, 3–4. [Google Scholar] [CrossRef]
- Howard, J.; Hyman, A. Dynamics and mechanics of the microtubule plus end. Nature 2003, 422, 753–758. [Google Scholar] [CrossRef]
- Gardner, M.K.; Zanic, M.; Howard, J. Microtubule catastrophe and rescue. Curr. Opin. Cell Biol. 2012, 25, 14–22. [Google Scholar] [CrossRef]
- Ebbinghaus, M.; Santen, L. Theoretical Modeling of Aging Effects in Microtubule Dynamics. Biophys. J. 2011, 100, 832–838. [Google Scholar] [CrossRef]
- Hyder, C.; Isoniemi, K.O.; Torvaldson, E.; Eriksson, J.E. Insights into intermediate filament regulation from development to ageing. J. Cell Sci. 2011, 124, 1363–1372. [Google Scholar] [CrossRef]
- Chesarone, M.A.; Goode, B.L. Actin nucleation and elongation factors: Mechanisms and interplay. Curr. Opin. Cell Biol. 2009, 21, 28–37. [Google Scholar] [CrossRef] [Green Version]
- Breitsprecher, D.; Goode, B.L. Formins at a glance. J. Cell Sci. 2013, 126, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Brunton, V.; MacPherson, I.; Frame, M. Cell adhesion receptors, tyrosine kinases and actin modulators: A complex three-way circuitry. Biochim. Biophys. Acta 2004, 1692, 121–144. [Google Scholar] [CrossRef] [PubMed]
- Papakonstanti, E.; Stournaras, C. Cell responses regulated by early reorganization of actin cytoskeleton. FEBS Lett. 2008, 582, 2120–2127. [Google Scholar] [CrossRef] [PubMed]
- Sato, O.; Sakai, T.; Choo, Y.-Y.; Ikebe, R.; Watanabe, T.M.; Ikebe, M. Mitochondria-associated myosin 19 processively transports mitochondria on actin tracks in living cells. J. Biol. Chem. 2022, 298, 101883. [Google Scholar] [CrossRef]
- Kruppa, A.J.; Buss, F. Motor proteins at the mitochondria–cytoskeleton interface. J. Cell Sci. 2021, 134. [Google Scholar] [CrossRef] [PubMed]
- Logan, C.M.; Menko, A.S. Microtubules: Evolving roles and critical cellular interactions. Exp. Biol. Med. 2019, 244, 1240–1254. [Google Scholar] [CrossRef]
- Hollenbeck, P.J.; Saxton, W.M. The axonal transport of mitochondria. J. Cell Sci. 2005, 118, 5411–5419. [Google Scholar] [CrossRef]
- Schmit, H.L.; Kraft, L.M.; Lee-Smith, C.; Lackner, L.L. The role of mitochondria in anchoring dynein to the cell cortex extends beyond clustering the anchor protein. Cell Cycle 2018, 17, 1345–1357. [Google Scholar] [CrossRef]
- Feinstein, S.C.; Wilson, L. Inability of tau to properly regulate neuronal microtubule dynamics: A loss-of-function mechanism by which tau might mediate neuronal cell death. Biochim. et Biophys. Acta (BBA)—Mol. Basis Dis. 2005, 1739, 268–279. [Google Scholar] [CrossRef]
- Zinsmaier, K.E.; Babic, M.; Russo, G.J. Mitochondrial Transport Dynamics in Axons and Dendrites. Cell Biol. Axon 2009, 48, 361–381. [Google Scholar] [CrossRef]
- Congdon, E.; Sigurdsson, E.M. Tau-targeting therapies for Alzheimer disease. Nat. Rev. Neurol. 2018, 14, 399–415. [Google Scholar] [CrossRef] [PubMed]
- Cash, A.D.; Aliev, G.; Siedlak, S.L.; Nunomura, A.; Fujioka, H.; Zhu, X.; Raina, A.K.; Vinters, H.V.; Tabaton, M.; Johnson, A.B.; et al. Microtubule Reduction in Alzheimer's Disease and Aging Is Independent of τ Filament Formation. Am. J. Pathol. 2003, 162, 1623–1627. [Google Scholar] [CrossRef]
- Lowery, J.; Kuczmarski, E.R.; Herrmann, H.; Goldman, R.D. Intermediate Filaments Play a Pivotal Role in Regulating Cell Architecture and Function. J. Biol. Chem. 2015, 290, 17145–17153. [Google Scholar] [CrossRef] [PubMed]
- Ball, E.H.; Singer, S.J. Mitochondria are associated with microtubules and not with intermediate filaments in cultured fibroblasts. Proc. Natl. Acad. Sci. USA 1982, 79, 123–126. [Google Scholar] [CrossRef]
- Handorf, A.M.; Zhou, Y.; Halanski, M.A.; Li, W.-J. Tissue Stiffness Dictates Development, Homeostasis, and Disease Progression. Organogenesis 2015, 11, 1–15. [Google Scholar] [CrossRef]
- Svitkina, T. The Actin Cytoskeleton and Actin-Based Motility. Cold Spring Harb. Perspect. Biol. 2018, 10, a018267. [Google Scholar] [CrossRef]
- Chou, S.Z.; Pollard, T.D. Mechanism of actin polymerization revealed by cryo-EM structures of actin filaments with three different bound nucleotides. Proc. Natl. Acad. Sci. USA 2019, 116, 4265–4274. [Google Scholar] [CrossRef]
- Pollard, T.D. Actin and Actin-Binding Proteins. Cold Spring Harb. Perspect. Biol. 2016, 8, a018226. [Google Scholar] [CrossRef]
- Tang, D.; Kang, R.; Berghe, T.V.; Vandenabeele, P.; Kroemer, G. The molecular machinery of regulated cell death. Cell Res. 2019, 29, 347–364. [Google Scholar] [CrossRef]
- Santella, L.; Limatola, N.; Chun, J.T. Cellular and molecular aspects of oocyte maturation and fertilization: A perspective from the actin cytoskeleton. Zool. Lett. 2020, 6, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Szarama, K.B.; Gavara, N.; Petralia, R.S.; Kelley, M.W.; Chadwick, R.S. Cytoskeletal changes in actin and microtubules underlie the developing surface mechanical properties of sensory and supporting cells in the mouse cochlea. Development 2012, 139, 2187–2197. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Huff, L.P.; Fujii, M.; Griendling, K.K. Redox regulation of the actin cytoskeleton and its role in the vascular system. Free Radic. Biol. Med. 2017, 109, 84–107. [Google Scholar] [CrossRef] [PubMed]
- Wilson, C.; González-Billault, C. Regulation of cytoskeletal dynamics by redox signaling and oxidative stress: Implications for neuronal development and trafficking. Front. Cell. Neurosci. 2015, 9, 381. [Google Scholar] [CrossRef] [PubMed]
- Islam, S.; Kabir, A.M.R.; Inoue, D.; Sada, K.; Kakugo, A. Enhanced dynamic instability of microtubules in a ROS free inert environment. Biophys. Chem. 2016, 211, 1–8. [Google Scholar] [CrossRef]
- Roy, J.; Galano, J.-M.; Durand, T.; Le Guennec, J.Y.; Lee, J.C.Y. Physiological role of reactive oxygen species as promoters of natural defenses. FASEB J. 2017, 31, 3729–3745. [Google Scholar] [CrossRef]
- Mittal, M.; Siddiqui, M.R.; Tran, K.; Reddy, S.P.; Malik, A.B. Reactive Oxygen Species in Inflammation and Tissue Injury. Antioxid. Redox Signal. 2014, 20, 1126–1167. [Google Scholar] [CrossRef]
- Taulet, N.; Delorme-Walker, V.D.; DerMardirossian, C. Reactive Oxygen Species Regulate Protrusion Efficiency by Controlling Actin Dynamics. PLoS ONE 2012, 7, e41342. [Google Scholar] [CrossRef]
- Lamari, F.; Braut-Boucher, F.; Pongnimitprasert, N.; Bernard, M.; Foglietti, M.-J.; Derappe, C.; Aubery, M. Cell adhesion and integrin expression are modulated by oxidative stress in EA.hy 926 cells. Free Radic. Res. 2007, 41, 812–822. [Google Scholar] [CrossRef]
- Sakai, J.; Li, J.; Subramanian, K.K.; Mondal, S.; Bajrami, B.; Hattori, H.; Jia, Y.; Dickinson, B.C.; Zhong, J.; Ye, K.; et al. Reactive Oxygen Species-Induced Actin Glutathionylation Controls Actin Dynamics in Neutrophils. Immunity 2012, 37, 1037–1049. [Google Scholar] [CrossRef]
- Stojkov, D.; Amini, P.; Oberson, K.; Sokollik, C.; Duppenthaler, A.; Simon, H.-U.; Yousefi, S. ROS and glutathionylation balance cytoskeletal dynamics in neutrophil extracellular trap formation. J. Cell Biol. 2017, 216, 4073–4090. [Google Scholar] [CrossRef]
- Yang, F.; Yi, M.; Liu, Y.; Wang, Q.; Hu, Y.; Deng, H. Glutaredoxin-1 Silencing Induces Cell Senescence via p53/p21/p16 Signaling Axis. J. Proteome Res. 2018, 17, 1091–1100. [Google Scholar] [CrossRef]
- Gallogly, M.M.; Shelton, M.D.; Qanungo, S.; Pai, H.V.; Starke, D.W.; Hoppel, C.L.; Lesnefsky, E.J.; Mieyal, J.J. Glutaredoxin Regulates Apoptosis in Cardiomyocytes via NFκB Targets Bcl-2 and Bcl-xL: Implications for Cardiac Aging. Antioxidants Redox Signal. 2010, 12, 1339–1353. [Google Scholar] [CrossRef]
- Gao, X.-H.; Qanungo, S.; Pai, H.V.; Starke, D.W.; Steller, K.M.; Fujioka, H.; Lesnefsky, E.J.; Kerner, J.; Rosca, M.G.; Hoppel, C.L.; et al. Aging-dependent changes in rat heart mitochondrial glutaredoxins—Implications for redox regulation. Redox Biol. 2013, 1, 586–598. [Google Scholar] [CrossRef]
- Gulati, S.; Thannickal, V.J. The Aging Lung and Idiopathic Pulmonary Fibrosis. Am. J. Med Sci. 2019, 357, 384–389. [Google Scholar] [CrossRef] [PubMed]
- Thannickal, V.J. Mechanistic links between aging and lung fibrosis. Biogerontology 2013, 14, 609–615. [Google Scholar] [CrossRef] [PubMed]
- Yanai, H.; Shteinberg, A.; Porat, Z.; Budovsky, A.; Braiman, A.; Zeische, R.; Fraifeld, V.E. Cellular senescence-like features of lung fibroblasts derived from idiopathic pulmonary fibrosis patients. Aging 2015, 7, 664–672. [Google Scholar] [CrossRef]
- Jaffar, J.; Yang, S.-H.; Kim, S.Y.; Kim, H.-W.; Faiz, A.; Chrzanowski, W.; Burgess, J.K.; Yang, S.-H. Greater cellular stiffness in fibroblasts from patients with idiopathic pulmonary fibrosis. Am. J. Physiol. Cell. Mol. Physiol. 2018, 315, L59–L65. [Google Scholar] [CrossRef]
- Apple, E.; Chen, L. Neuronal microtubules impact lifespan. Aging 2019, 11, 6616–6617. [Google Scholar] [CrossRef]
- Spears, W.; Furgerson, M.; Sweetnam, J.M.; Evans, P.; Gearing, M.; Fechheimer, M.; Furukawa, R. Hirano bodies differentially modulate cell death induced by tau and the amyloid precursor protein intracellular domain. BMC Neurosci. 2014, 15, 74. [Google Scholar] [CrossRef]
- Biernat, J.; Gustke, N.; Drewes, G.; Mandelkow, E. Phosphorylation of Ser262 strongly reduces binding of tau to microtubules: Distinction between PHF-like immunoreactivity and microtubule binding. Neuron 1993, 11, 153–163. [Google Scholar] [CrossRef]
- Hong, M.; Zhukareva, V.; Vogelsberg-Ragaglia, V.; Wszolek, Z.; Reed, L.; Miller, B.I.; Geschwind, D.H.; Bird, T.D.; McKeel, D.; Goate, A.; et al. Mutation-Specific Functional Impairments in Distinct Tau Isoforms of Hereditary FTDP-17. Science 1998, 282, 1914–1917. [Google Scholar] [CrossRef] [PubMed]
- Kerr, J.P.; Robison, P.; Shi, G.; Bogush, A.I.; Kempema, A.M.; Hexum, J.K.; Becerra, N.; Harki, D.A.; Martin, S.S.; Raiteri, R.; et al. Detyrosinated microtubules modulate mechanotransduction in heart and skeletal muscle. Nat. Commun. 2015, 6, 8526. [Google Scholar] [CrossRef]
- Han, S.-W.; Simona, P.; Banu, M.; Adachi, T. Real-time monitoring of changes in microtubule mechanical properties in response to microtubule-destabilizing drug treatment. J. Nanosci. Nanotechnol. 2013, 13, 2087–2090. [Google Scholar] [CrossRef] [PubMed]
- Schulze, C.; Wetzel, F.; Kueper, T.; Malsen, A.; Muhr, G.; Jaspers, S.; Blatt, T.; Wittern, K.-P.; Wenck, H.; Käs, J.A. Stiffening of Human Skin Fibroblasts with Age. Biophys. J. 2010, 99, 2434–2442. [Google Scholar] [CrossRef] [PubMed]
- Caporizzo, M.A.; Chen, C.Y.; Bedi, K.; Margulies, K.B.; Prosser, B.L. Microtubules Increase Diastolic Stiffness in Failing Human Cardiomyocytes and Myocardium. Circulation 2020, 141, 902–915. [Google Scholar] [CrossRef] [PubMed]
- Koide, M.; Hamawaki, M.; Narishige, T.; Sato, H.; Nemoto, S.; Defreyte, G.; Zile, M.R.; Cooper, I.G.; Carabello, B.A. Microtubule Depolymerization Normalizes In Vivo Myocardial Contractile Function in Dogs with Pressure-Overload Left Ventricular Hypertrophy. Circulation 2000, 102, 1045–1052. [Google Scholar] [CrossRef]
- Prins, K.W.; Tian, L.; Wu, D.; Thenappan, T.; Metzger, J.M.; Archer, S.L. Colchicine Depolymerizes Microtubules, Increases Junctophilin-2, and Improves Right Ventricular Function in Experimental Pulmonary Arterial Hypertension. J. Am. Heart Assoc. 2017, 6, e006195. [Google Scholar] [CrossRef]
- Scopacasa, B.S.; Teixeira, V.P.A.; Franchini, K.G. Colchicine attenuates left ventricular hypertrophy but preserves cardiac function of aortic-constricted rats. J. Appl. Physiol. 2003, 94, 1627–1633. [Google Scholar] [CrossRef]
- Kim, M.J.; Choi, K.H.; Seo, D.W.; Lee, H.R.; Kong, H.S.; Lee, C.H.; Lee, W.S.; Lee, H.T.; Ko, J.J.; Kim, J.H.; et al. Association Between Functional Activity of Mitochondria and Actin Cytoskeleton Instability in Oocytes from Advanced Age Mice. Reprod. Sci. 2020, 27, 1037–1046. [Google Scholar] [CrossRef]
- Yoshida, H.; Goedert, M. Phosphorylation of microtubule-associated protein tau by AMPK-related kinases. J. Neurochem. 2011, 120, 165–176. [Google Scholar] [CrossRef]
- Asnaghi, L.; Bruno, P.; Priulla, M.; Nicolin, A. mTOR: A protein kinase switching between life and death. Pharmacol. Res. 2004, 50, 545–549. [Google Scholar] [CrossRef] [PubMed]
- Tee, A.R. Fundamental for life: mTOR orchestrates developing biological systems. Semin. Cell Dev. Biol. 2014, 36, 66–67. [Google Scholar] [CrossRef] [PubMed]
- Bratic, A.; Larsson, N.-G. The role of mitochondria in aging. J. Clin. Investig. 2013, 123, 951–957. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Coulombe, P.A. Intermediate filament scaffolds fulfill mechanical, organizational, and signaling functions in the cytoplasm. Genes Dev. 2007, 21, 1581–1597. [Google Scholar] [CrossRef] [PubMed]
- Langlois, B.; Belozertseva, E.; Parlakian, A.; Bourhim, M.; Gao-Li, J.; Blanc, J.; Tian, L.; Coletti, D.; Labat, C.; Ramdame-Cherif, Z.; et al. Vimentin knockout results in increased expression of sub-endothelial basement membrane components and carotid stiffness in mice. Sci. Rep. 2017, 7, 11628. [Google Scholar] [CrossRef]
- Lacolley, P.; Regnault, V.; Avolio, A.P. Smooth muscle cell and arterial aging: Basic and clinical aspects. Cardiovasc. Res. 2018, 114, 513–528. [Google Scholar] [CrossRef]
- Danielsson, F.; Peterson, M.K.; Araújo, H.C.; Lautenschläger, F.; Gad, A.K.B. Vimentin Diversity in Health and Disease. Cells 2018, 7, 147. [Google Scholar] [CrossRef]
- Van Engeland, N.C.A.; Rodriguez, F.S.; Rivero-Müller, A.; Ristori, T.; Duran, C.L.; Stassen, O.M.J.A.; Antfolk, D.; Driessen, R.C.H.; Ruohonen, S.; Ruohonen, S.T.; et al. Vimentin regulates Notch signaling strength and arterial remodeling in response to hemodynamic stress. Sci. Rep. 2019, 9, 12415. [Google Scholar] [CrossRef]
- Kueper, T.; Grune, T.; Prahl, S.; Lenz, H.; Welge, V.; Biernoth, T.; Vogt, Y.; Muhr, G.-M.; Gaemlich, A.; Jung, T.; et al. Vimentin Is the Specific Target in Skin Glycation. J. Biol. Chem. 2007, 282, 23427–23436. [Google Scholar] [CrossRef]
- Chahine, N.O.; Blanchette, C.; Thomas, C.B.; Lu, J.; Haudenschild, D.; Loots, G.G. Effect of Age and Cytoskeletal Elements on the Indentation-Dependent Mechanical Properties of Chondrocytes. PLoS ONE 2013, 8, e61651. [Google Scholar] [CrossRef] [Green Version]
- Steklov, N.; Srivastava, A.; Sung, K.L.P.; Chen, P.C.; Lotz, M.K.; D'Lima, D.D. Aging-related differences in chondrocyte viscoelastic properties. Mol. Cell Biomech. 2009, 6, 113–119. [Google Scholar] [PubMed]
- Song, S.; Landsbury, A.; Dahm, R.; Liu, Y.; Zhang, Q.; Quinlan, R.A. Functions of the intermediate filament cytoskeleton in the eye lens. J. Clin. Investig. 2009, 119, 1837–1848. [Google Scholar] [CrossRef] [PubMed]
- Michael, R.; Bron, A.J. The ageing lens and cataract: A model of normal and pathological ageing. Philos. Trans. R. Soc. B Biol. Sci. 2011, 366, 1278–1292. [Google Scholar] [CrossRef] [PubMed]
- Jakobs, P.M.; Hess, J.F.; FitzGerald, P.G.; Kramer, P.; Weleber, R.G.; Litt, M. Autosomal-Dominant Congenital Cataract Associated with a Deletion Mutation in the Human Beaded Filament Protein Gene BFSP2. Am. J. Hum. Genet. 2000, 66, 1432–1436. [Google Scholar] [CrossRef]
- Yang, Y.-H.K. Aging of mesenchymal stem cells: Implication in regenerative medicine. Regen. Ther. 2018, 9, 120–122. [Google Scholar] [CrossRef]
- Swaab, D.; Salehi, A. The pathogenesis of Alzheimer disease: An alternative to the amyloid hypothesis. J. Neuropathol. Exp. Neurol. 1997, 56, 216. [Google Scholar] [CrossRef]
- Pavlović, D.M.; Pavlović, A.M.; Zugic, S. Pathogenesis of Alzheimer disease. Vojn. Pregl. 2007, 64, 765–772. [Google Scholar] [CrossRef]
- Mercado, N.; Ito, K.; Barnes, P.J. Accelerated ageing of the lung in COPD: New concepts. Thorax 2015, 70, 482–489. [Google Scholar] [CrossRef]
- D'Anna, C.; Cigna, D.; Di Sano, C.; Di Vincenzo, S.; Dino, P.; Ferraro, M.; Bini, L.; Bianchi, L.; Di Gaudio, F.; Gjomarkaj, M.; et al. Exposure to cigarette smoke extract and lipopolysaccharide modifies cytoskeleton organization in bronchial epithelial cells. Exp. Lung Res. 2017, 43, 347–358. [Google Scholar] [CrossRef]
- Heijink, I.H.; Noordhoek, J.A.; Timens, W.; van Oosterhout, A.J.M.; Postma, D.S. Abnormalities in Airway Epithelial Junction Formation in Chronic Obstructive Pulmonary Disease. Am. J. Respir. Crit. Care Med. 2014, 189, 1439–1442. [Google Scholar] [CrossRef]
- Bidan, C.; Veldsink, A.C.; Meurs, H.; Gosens, R. Airway and Extracellular Matrix Mechanics in COPD. Front. Physiol. 2015, 6, 346. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.; Kim, Y.; Cho, M.; Kim, G.; Kim, S.; Kim, M.; Ko, J.; Lee, J. MIT-001 Restores Human Placenta-Derived Mesenchymal Stem Cells by Enhancing Mitochondrial Quiescence and Cytoskeletal Organization. Int. J. Mol. Sci. 2021, 22, 5062. [Google Scholar] [CrossRef] [PubMed]
- Frey, A.; Lunding, L.P.; Ehlers, J.C.; Weckmann, M.; Zissler, U.M.; Wegmann, M. More Than Just a Barrier: The Immune Functions of the Airway Epithelium in Asthma Pathogenesis. Front. Immunol. 2020, 11, 761. [Google Scholar] [CrossRef] [PubMed]
- Nishida, K.; Brune, K.A.; Putcha, N.; Mandke, P.; O’Neal, W.K.; Shade, D.; Srivastava, V.; Wang, M.; Lam, H.; An, S.; et al. Cigarette smoke disrupts monolayer integrity by altering epithelial cell-cell adhesion and cortical tension. Am. J. Physiol. Cell. Mol. Physiol. 2017, 313, L581–L591. [Google Scholar] [CrossRef] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, Y.J.; Cho, M.J.; Yu, W.D.; Kim, M.J.; Kim, S.Y.; Lee, J.H. Links of Cytoskeletal Integrity with Disease and Aging. Cells 2022, 11, 2896. https://doi.org/10.3390/cells11182896
Kim YJ, Cho MJ, Yu WD, Kim MJ, Kim SY, Lee JH. Links of Cytoskeletal Integrity with Disease and Aging. Cells. 2022; 11(18):2896. https://doi.org/10.3390/cells11182896
Chicago/Turabian StyleKim, Yu Jin, Min Jeong Cho, Won Dong Yu, Myung Joo Kim, Sally Yunsun Kim, and Jae Ho Lee. 2022. "Links of Cytoskeletal Integrity with Disease and Aging" Cells 11, no. 18: 2896. https://doi.org/10.3390/cells11182896
APA StyleKim, Y. J., Cho, M. J., Yu, W. D., Kim, M. J., Kim, S. Y., & Lee, J. H. (2022). Links of Cytoskeletal Integrity with Disease and Aging. Cells, 11(18), 2896. https://doi.org/10.3390/cells11182896