1. Introduction
The human mast cell (MC) is an immune cell of hematopoietic origin and like a two-faced joker, the MC is well recognized for its role in pathological settings yet acknowledged for its protective role in health [
1]. MCs are abundant in the airway mucosa and epithelium, but also within the septa of the alveolar parenchyma [
2]. However, the role of alveolar MCs is largely unknown. MC subpopulations are binary phenotyped by their protease content in granula; MCs storing tryptase (MC
T) and MCs storing tryptase and chymase (MC
TC) [
1]. In pulmonary baseline conditions MC proteases have a key role in homeostasis and wound healing; enhancing migration, proliferation, and gap closure capacity, but also inducing the release of growth factors associated with tissue repair in fibroblasts and epithelial cells [
3,
4,
5,
6]. However, in an asthmatic context an increase of MCs and change of distribution and phenotype in both bronchial and alveolar compartments often associate with disease severity [
2,
7]. It has also been shown that MC proteases contribute to airway bronchoconstriction, hyperresponsiveness and airway remodelling [
8].
The binding of urokinase plasminogen activator (uPA) to its glycosylphosphatidylinositol-inositol (GPI) anchored plasminogen activator urokinase receptor (uPAR) initiates several cellular pathways that modulate a variety of functions including homeostasis, inflammation, and wound healing [
9,
10]. The uPA/uPAR system has an important role in the regulation of cellular mitogenic effects such as migration and proliferation in relation to events associated with normal cellular development and tissue repair. However, dysregulated cellular proliferation and migration contributes to plural disorders with remodelling and fibrotic characteristics [
9,
11].
uPAR is composed as a three domain (D1, D2 and D3) GPI anchored membrane bound receptor, but also exists as an anchored cleaved form referred to as suPAR, with high affinity for the serine uPA. The zymogen form of uPA, pro-uPA, also binds to uPAR but with less affinity [
9,
12]. The uPA molecule is produced by smooth muscle, endothelial and epithelial cells as wells as fibroblasts and monocytes. It is secreted in its zymogen single chain form and by proteolysis it is actuated into a two-chain active form [
9,
10,
12]. The structure of the uPA molecule is trimeric and divided into amino-terminal “growth factor domain”, the kringle domain and the carboxy-terminal catalytic domain [
9]. The binding of uPA-uPAR occurs between uPA’s “growth factor domain” and uPAR’s domain D1 as well as D3 [
10]. The most frequent description of uPA-uPAR interaction is the role of regulator of the plasminogen activation system, where uPA converts plasminogen to protease plasmin. Plasmin has a key role in degradation of extra cellular matrix components (ECM), lysis of fibrin clots and activation of matrix metalloproteases (MMPs) [
12,
13]. The uPA-uPAR interaction generates a positive feedback loop since plasmin can cleave and activate the zymogen to uPA’s active two chain form. The system is regulated negatively by covalent binding and inhibition of proteolytic activity between serine protease inhibitor plasminogen inhibitor 1 (PAI-1) and plasminogen inhibitor 2 (PAI-2) and their targets [
10,
12].
Studies have shown that the u-PA/u-PAR system is activated during asthmatic conditions as well as in other inflammatory settings [
7,
11,
14,
15]. Further, it has been identified that MC tryptase is a potent activator of pro-uPA and that MC chymase increases levels of suPAR in fibroblasts [
16,
17].
Asthma is a disorder characterized by remodelling and most asthma studies have focused on the large airways. However, MCs are present but less studied in the peripheral and alveolar compartments of the lung [
2,
18,
19]. The uPA-uPAR system has been found to regulate epithelial repair mechanisms [
11]. Given this, it is established that MC proteases and the uPA-uPAR system are implicated in asthma; however, the outcome of their interactions in asthmatic disease has not fully been understood. Therefore, by using human alveolar basal epithelial cells (A549) we were able to examine the effect of MC tryptase and chymase on cellular functional properties and the protease impact of uPAR expression. We hypothesize that MC proteases will have an impact of functional structures such as wound healing, migration, and morphology in alveolar epithelial cells. Further, we also expected an impact of uPA/uPAR expression and uPAR related cellular events such as migration in the presence of MC proteases. Our data can confirm earlier bronchial findings of the role of tryptase in an alveolar context with increased gap closure capacity and enhanced migration. However, an opposite effect was observed in chymase stimulated cells. We can also report that tryptase upregulated the expression of the membrane bound uPAR while chymase increased the release of the soluble (suPAR) receptor. Finally, we identified that dual asthmatic responders had higher concentrations of suPAR compared to single asthmatic responders. However, single asthmatic responders had more suPAR in the alveolar fractions compared with the bronchial fractions. Our findings present the possibility that MC via their proteases may contribute to dual regulation on parameters associated with repair mechanisms that are important features in asthmatic disease.
2. Materials and Methods
2.1. Subjects
This study included 15 non-smoking patients with mild to moderate allergic asthma of whom 9 had received a low dose of inhaled corticosteroids and the remainder were steroid naïve. The subjects displayed either early allergic reaction only (termed as single responders) or both early and a late allergic reaction (dual responders) post inhaled allergen. No other significant differences in patient characteristics were found (see
Table 1 for details). All subjects gave their written informed consent and the Regional Ethics Review Board in Lund, Sweden, approved the study (2012/800). The subjects have been used in previously published studies [
20,
21]. Subjects underwent bronchoscopy as previously described [
21] before and 24 h post allergen challenge where 3 × 50 mL phosphate-buffered saline (PBS) was instilled via a bronchoscope to the middle lobe. The first 50 mL was collected separately and referred to as bronchial lavage and the remaining PBS was recovered as alveolar fraction. Dual and single responders were determined by a reading of the FEV
1 between 0–2 h and 4–8 h after an administrated allergen challenge. For the current study, samples collected at baseline were used.
2.2. Cell Culture
Alveolar basal epithelial cells (A549) from ECACC (Porton Down, Salisbury, UK) were cultured in Ham’s F-12 nutrient Mix (Gibco, 21765-037) with 10% fetal bovine serum (FBS) and 1% penicillin-streptomycin from Life Technologies (Stockholm, Sweden). For detachment of cells, trypsin-EDTA 0.25% was used (Gibco, 25200056). For the experimental set-up (RNA, protein and supernatant) cells were cultured in a 6 well Nunc multidish (Nunc Technologies, Carlsbad, CA, USA) at 37 °C in 5% CO
2 and were grown until approximately 80–90% confluency which was determined automatically using a live cell imaging system (Holomonitor Phase Holographic Imaging, Lund, Sweden). Cells were treated with 0.5 μg/mL human lung tryptase (Merck Millipore, Darmstadt, Germany) and 0.5 μg/mL recombinant chymase (Sigma-Aldrich, St. Louis, MO, USA, C8118-50 μg) [
5,
6]. During cellular experiments starvation medium was used (Ham’s F-12 nutrient Mix medium with 1% FBS and 1% penicillin-streptomycin). RNA and supernatants were collected at 6 h and protein and supernatant were collected 24 h post stimulation.
2.3. RNA Extraction and RT2 PCR Array
After 6 h stimulation, mRNA was collected and isolated with Aurum total RNA MiniKit (Bio-Rad, Hercules, CA, USA, 7326820). RNA concentration was measured with nanodrop 2000c Thermo Scientific (Waltham, MA, USA). Using RT2 First Strand Kit from QIAGEN (Hilden, Germany), RNA was transcribed to cDNA. For PCR analysis, RT2 SYBR Green Rox Master Mix (QIAGEN, 330521) and RT2 primer PLAUR (u-PAR) (PPH00797B) were used. For housekeeping, genes ACTB (QIAGEN, PPH00073G) and HPRT1 (QIAGEN, PPH01018C) were used.
2.4. ELISA and LDH Test
Lung lavage fractions from allergic asthmatics and cellular supernatant from A549 cell line and secretion of protein release were analyzed with ELISA DuoSet Kits from R&D systems (Minneapolis, MN, USA) for soluble uPAR (DY807), MMP-9 (DY911-05), PAI-1 (DY1786) and uPA (DY1310). In order to measure possible cytotoxicity, lactate dehydrogenase (LDH) test was used (Roche Diagnostics, Basal, Switzerland). Supernatant was collected at 1 h, 6 h and 24 h and testing was done according to the manufacturer’s protocol.
2.5. Western Blot
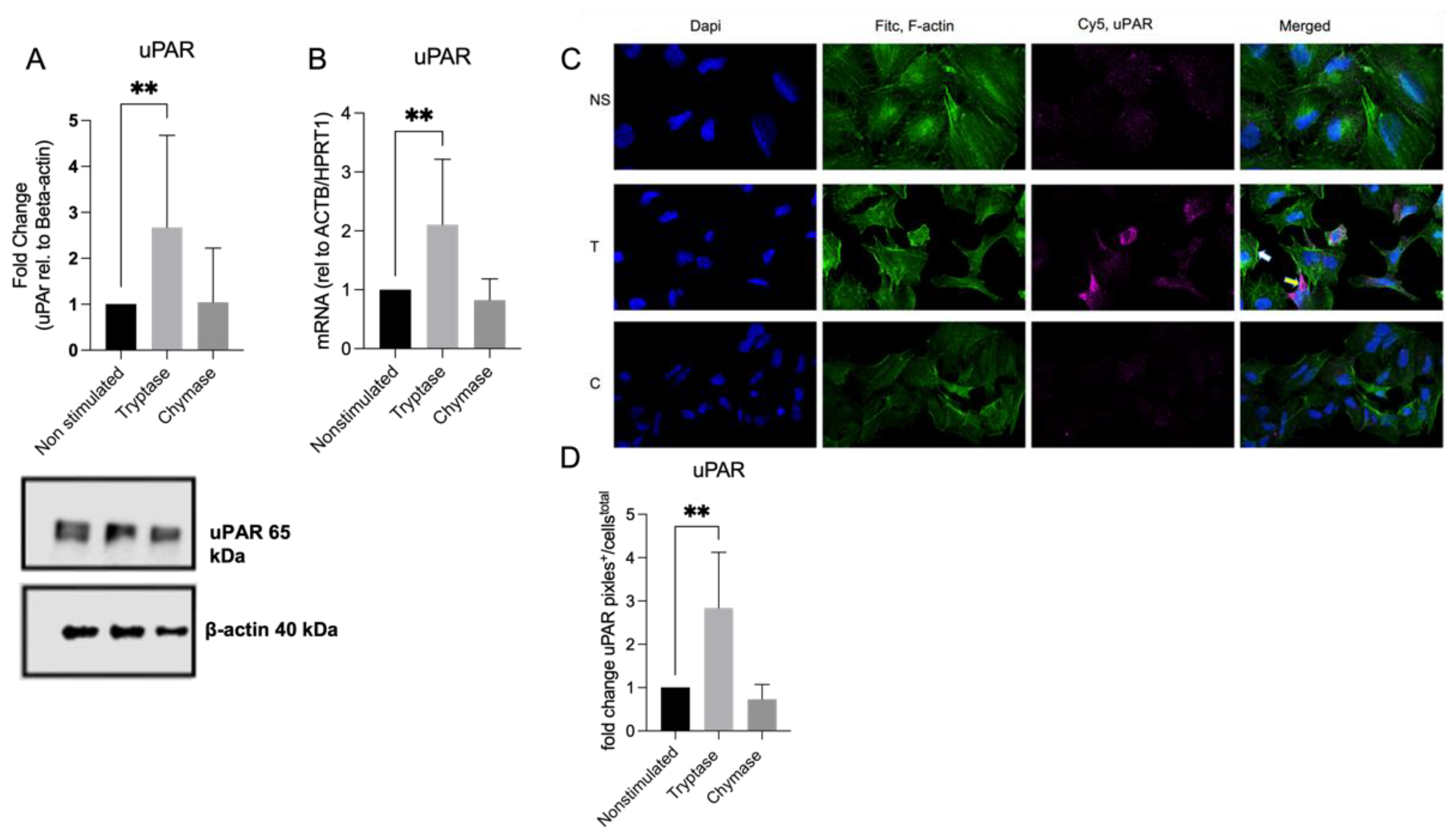
Protein was collected at 24 h and the concentration was quantified using Pierce BCA Protein Assay Kit. Samples were diluted with Laemmeli and boiled at 95 °C for 5 min using QIA Amplifier 96 QIAGEN (Hilden, Germany). Samples were added to Mini Protean TGX stain free gel (Bio-Rad, 4668086) and blotted with Transfer Blot Turbo (Bio-Rad, 1704156). The membranes were incubated with uPAR antibody 1:1000 (Cell Signaling, D4Q5S), and reference gene ß-actin, diluted 1:1000 (Cell signalling, Danvers, MA, USA, D6A8) overnight at 4 °C on a sample rocker. Thereafter, the membranes were incubated with secondary HRP conjugated antibody diluted 1:5000 (Bio-Rad, 5213-2504). For chemiluminescent detection, LI-COR Odyssey Fc and Image analysis was performed with Image Studio ver.5.2 (Lincoln, NE, USA).
2.6. Immunocytochemistry
A549 were cultured in chamber slides (Merck Millipore, Darmstadt, Germany) until 60% confluency or 90% confluency for scratch treatment. Tryptase and chymase were added and samples were fixated in 2% paraformaldehyde for 20 min post 24 h addition. For scratch staining, the samples were wounded with a single cut using 200 μL pipette tip prior to addition of proteases. Thereafter, samples were incubated with 0.1% Tween-20 for 10 min. Samples were immunostained with Primary antibody diluted 1:50, uPAR (Cell Signaling, D7X2N) and thereafter labelled with Alexa Fluor 488 or Alexa Flour 647 diluted 1:200 (Invitrogen, Eugene, OR, USA). For double immunostaining of uPAR in relation to the cytoskeleton arrangement, FITC conjugated phalloidin (Sigma-Aldrich, St Louis, MO, USA) was added and diluted 1:1000 post removal of secondary Alexa Fluor antibody. ProLong Glass antifade with NucblueTM (Thermo Fisher Scientific, Waltham, MA, USA, P36983) were used for mounting and nuclei visualization. A Nikon eclipse 80i combined with Nikon DS-QI1MC was used to view and document images.
2.7. Live Cell Imaging
For functional studies of wound gap closure and migration, A549 were cultured in a Sarstedt TC 6-well plate (Nümbrecht, Germany). For study of wound gaps, model cells were applied to culture-insert 2 well (Ibidi, Martinsried, Germany) with the concentration of 5 × 10
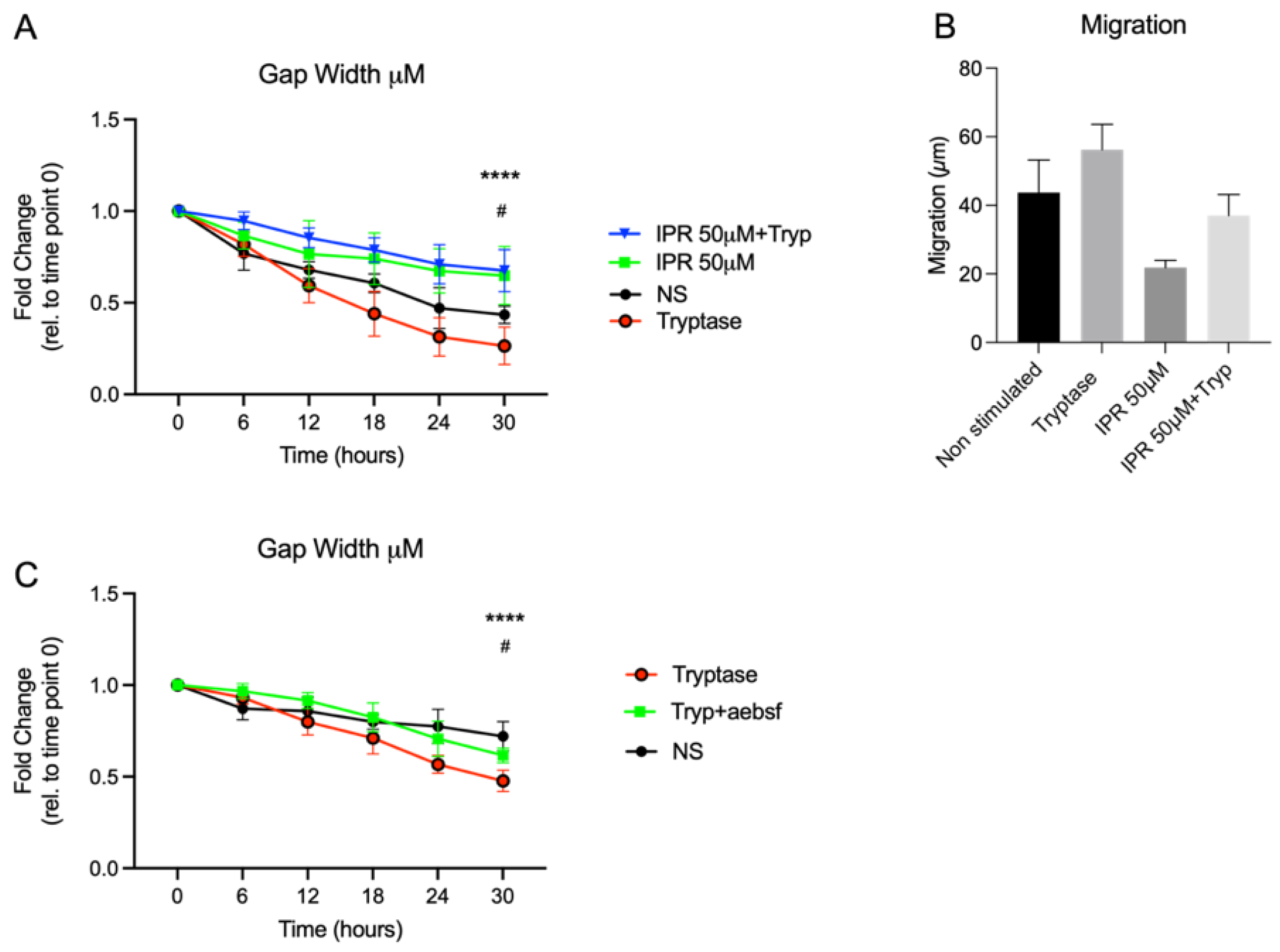
5 cells/mL. After 24 h, the inserts were removed, and cells were treated with tryptase or chymase with the working concentration of 0.5 μg/mL. For studying the uPA-uPAR effect, cells were pre-treated with uPA-uPAR inhibitor IPR-803 (Sollentuna, Sweden, HY-111192) at the concentration of 50 μM for three hours at 37 °C in 5% CO
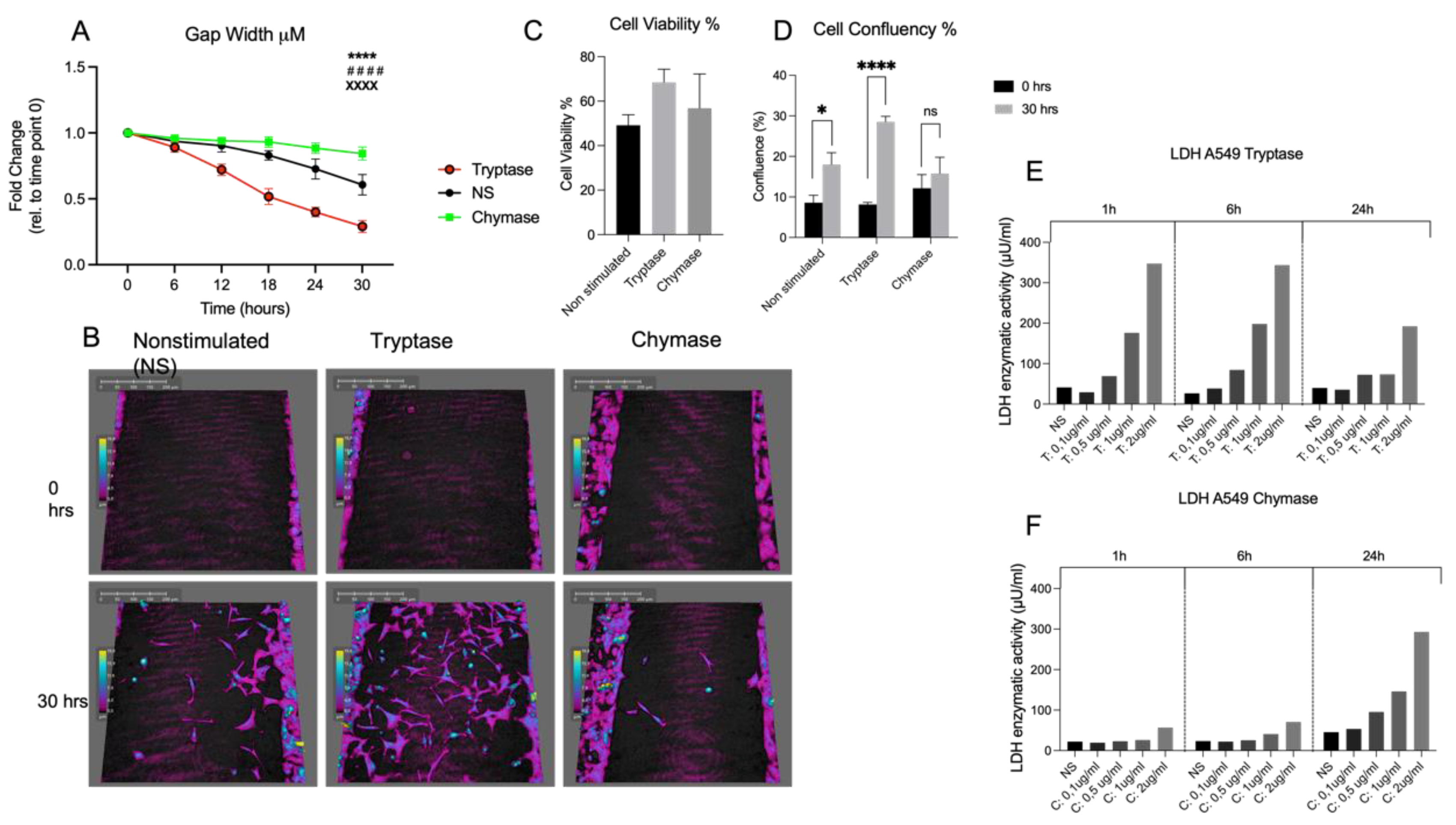
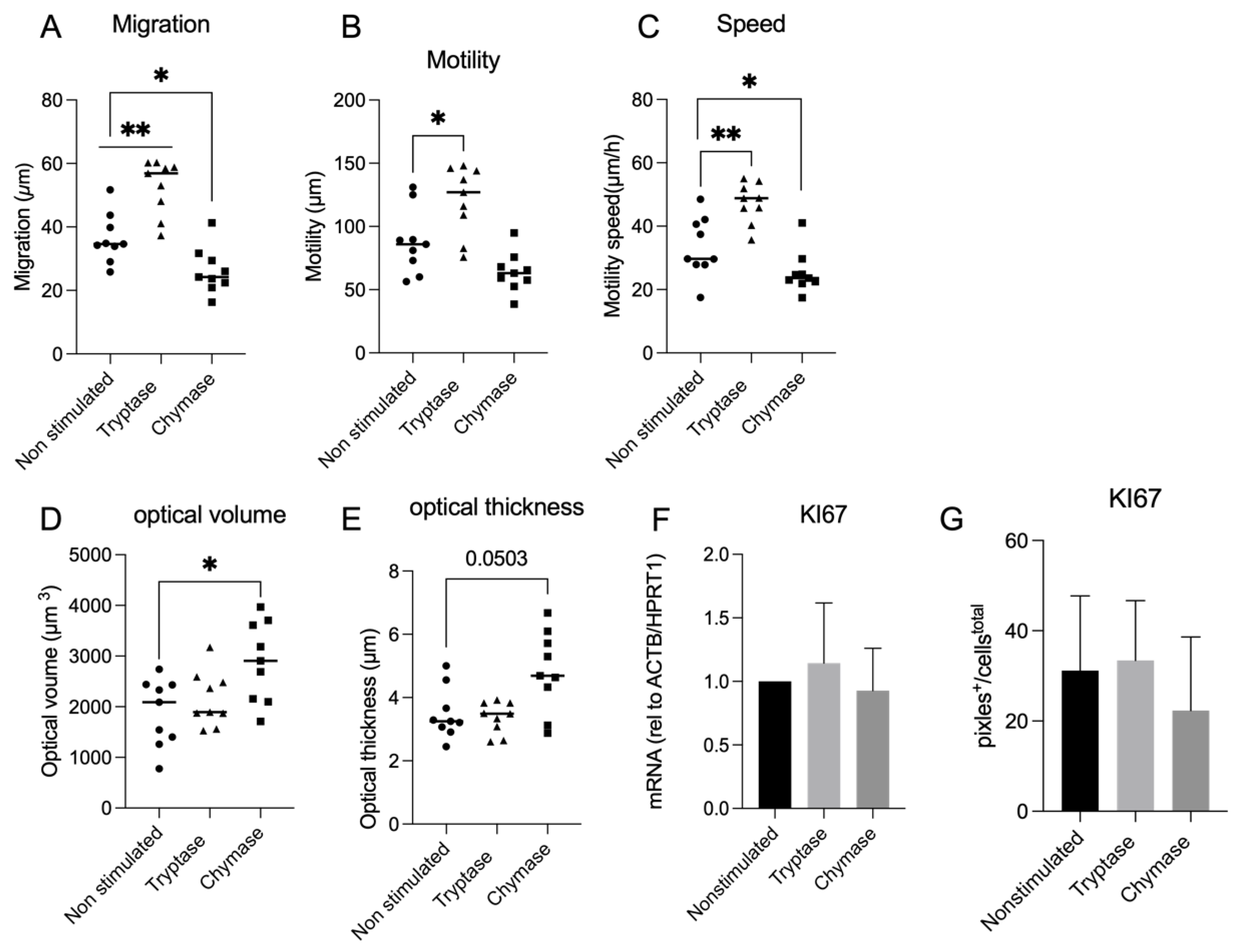
2 and thereafter washed three times with PBS prior to addition of MC proteases. A cell suspension with tryptase was pre-treated for 30 min at 37 °C with 200 μM of a broad serine protease inhibitor, 4-(2-aminoethyl) benzene sulfonyl fluoride hydrochloride (AEBSF) (Sigma-Aldrich, Stockholm, Sweden) in order to confirm the tryptase effect on a functional level. Cells were monitored for 30 h using HoloMonitor M4 live cell imaging system Phase Holographic Imaging (Lund, Sweden). Three randomly chosen positions in each well were selected for image capture with images taken every 15 min, resulting in a total of 2322 images. Quantitative phase imaging of gap closure was done in Holomonitor App Suite version 3.4.0.158 and quantification is based on the fold change of cell covered area. Analysis of cellular migration was performed as earlier described [
5]. Briefly, it used single cell tracking of a minimum of 9 randomly chosen cells per well, and technical repeats from the recorded images were performed. The software track computed the tracking data and calculated the cellular movements. However, the migration is based on data from the last 6 h of imaging, timepoint 24 h to timepoint 30 h. This to avoid issues related to cells migrating out of the picture, which prevents adequate monitoring of cells from start at 0 to endpoint at 30 h. Wound healing and migration data are based on three technical repeats. Migration was measured in µm and refers to the distance between start position and end position. Motility was also measured in µm and refers to the entire distance including spontaneous movements. Cellular speed was measured in µm/hour and morphology refers to the measured optical volume (µm
3) and thickness (µm) of the cells.
2.8. Confocal Microscopy
The samples were visualized on a Nikon A1+ confocal microscope with a 20× Plan Apo objective (NA 0.75) (Nikon Instruments Inc., Tokyo, Japan). Images were acquired and processed with NIS-elements, version: 4.60.02, (Laboratory Imaging, Nikon, Tokyo, Japan).
2.9. Cell Viability
Alveolar epithelial cells were cultured and stimulated with MC proteases as described above. Cells were removed using TrypLe Express (12604013, Gibco, Thermo Fisher Scientific, Waltham, MA, USA) and cell suspensions were diluted 1:1 with trypan blue (15250061, Gibco Thermo Fisher Scientific, Waltham, MA, USA). Thereafter, 10 μL of the cell suspension was added to an automatised cell counter (DeNovix, Celldrop Brightfield Cell Counter, Wilmington, DE, USA) for quantification of cell viability where dead cells display an entry of trypan blue and are stained compared to live cells that remain unstained.
2.10. Statistical Analysis
Statistical analysis was performed using GraphPad Prism 9.1.2 (GraphPad Software, La Jolla, CA, USA). Nonparametric Mann–Whitney U tests were used to detect differences between 2 groups. For multiple comparison between more than two groups at different time points two-way ANOVA with Tukey’s multiple comparison test was used. Results were considered significant at p ≤ 0.05. The data will be presented as Mean ± SD.
4. Discussion
Our study revealed that MC tryptase and chymase have binary impacts on alveolar epithelial gap reduction, migration, and morphology. Further, this study demonstrated a dual impact of tryptase and chymase related to the uPAR expression. Current findings could confirm the mitogenic effect of tryptase in terms of gap closure, migration, motility and cellular speed in an alveolar setting that we and others have previously found in human bronchial epithelial cells and fibroblasts [
5,
6,
22,
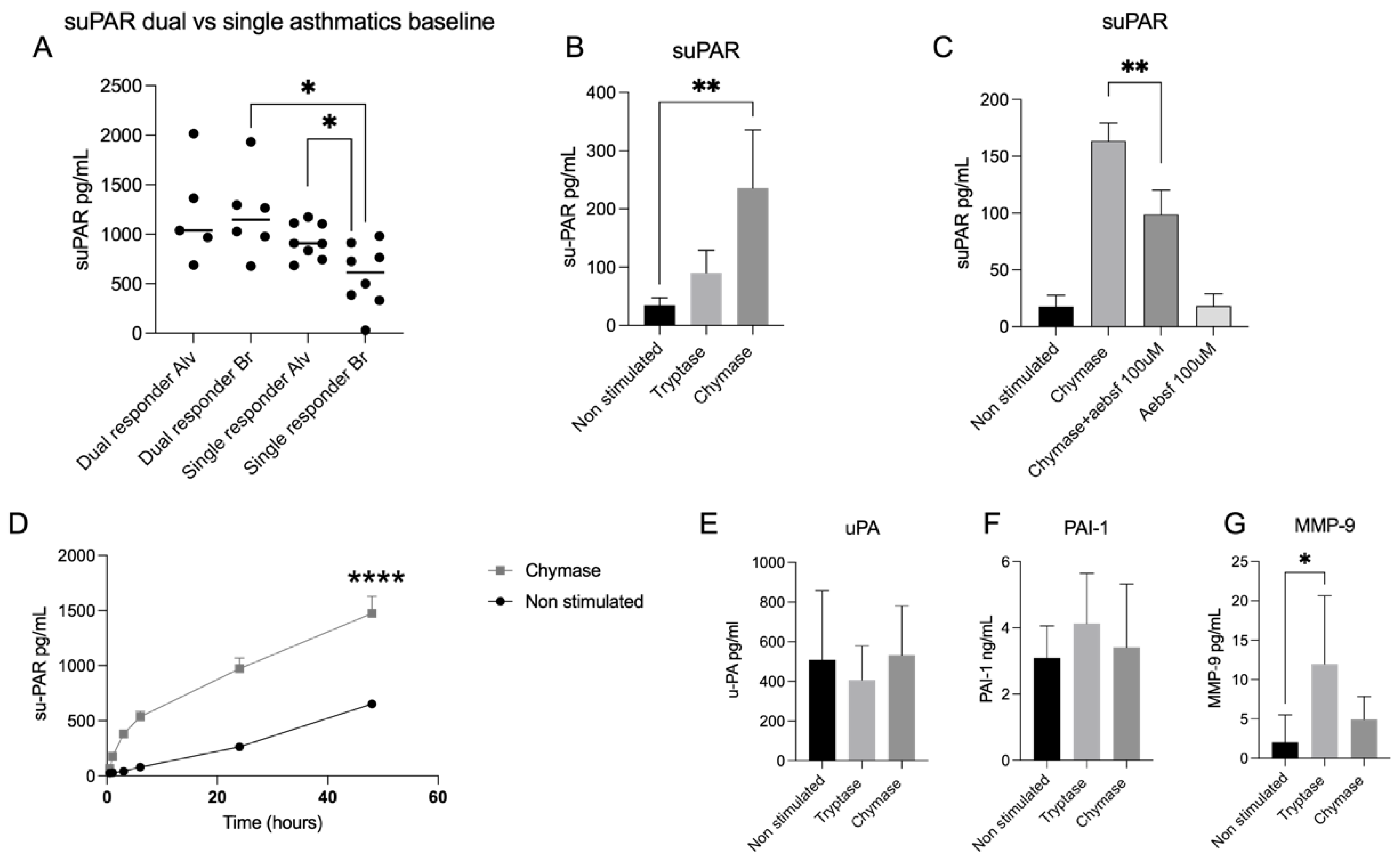
23]. Finally, we can also report an increased released of suPAR from an alveolar lavage fraction compared to bronchial lavage fraction in asthmatic patients with early allergic onset. We also detected a higher release of suPAR in asthmatic patients with both early and late reactions compared with patients with early onset only.
We could also confirm morphological alterations as an effect of chymase which has previously been shown in fibroblasts [
17]. Observed morphological changes in the present study was revealed as a quantitative increase of optical volume and optical thickness. This could possibly be an important observation from the perspective of airway remodelling. Furthermore, tryptase stimulated an increase of MMP-9, which is a metalloproteinase that is strongly related to impacts on the composition of the extra cellular matrix (ECM) [
18]. Apart from increase of MMP-9, tryptase stimulated cells also displayed a prominent pro-migratory effect; both are important features of normal physiological development and re-epithelialization upon injury but can also contribute to structural epithelial changes, which are observed in asthma [
24,
25]. In contrast, our data showed that chymase had an impaired effect on gap closure, migration, motility, and cellular speed. Our data highlight the necessity for a detailed understanding of the influence of tryptase and chymase on a molecular level and how the interaction with epithelial cells might differ depending on the micro localization. This finding may have clinical implications since chymase positive MCs increase in the alveolar compartment in several chronic respiratory diseases such as uncontrolled asthma, COPD and IPF [
2,
26] where parenchymal remodelling is an important feature.
The uPA-uPAR system participates in normal cellular development as well as in tissue remodelling with plural downstream effects related to ECM degradation but also changes in functional properties such as migration or proliferation. Accumulating evidence suggests a pivotal role for the uPA-uPAR system in chronic respiratory diseases, where it may contribute to epithelial alterations that generate airway remodelling [
11,
12,
27]. Since the accumulation of activated MCs is an established hallmark of asthma, we wanted to investigate the expression of uPAR in alveolar epithelial cells in the presence of MC proteases tryptase and chymase. Our findings reveal that chymase increases the release of suPAR in alveolar epithelial cells. This has previously been shown in fibroblasts [
17] but to the best of our knowledge it has not been found in an alveolar epithelial context before. We can also report that the effect of chymase remained over time (48 h), suggesting not only an enzymatic effect of chymase activity, that has shown to decrease rapidly within the first 6 h of stimulation [
5], but also the involvement of possible other mechanisms. Considering that increased levels of suPAR are connected with asthma prevalence [
27] and that increased alveolar infiltration of MC
TC is a fingerprint of asthmatic disease severity [
2], the chymase effect of suPAR is a highly interesting target for future validation in asthmatic disease. We can in the current study report an increase of suPAR levels in the alveolar lavage fractions compared with the bronchial lavage fractions in allergic asthmatics. Further, we could also detect higher levels of suPAR in dual responders compared with single responders, which indicates that suPAR could be correlated with asthma severity since dual responders display a vast and long-lasting airway inflammation compared to single responders [
20]. Another finding in our study showed that tryptase stimulated cells had higher expression of uPAR both on protein and gene level. Increased levels of uPAR have been shown to contribute to impaired wound healing capacity in asthmatic bronchial epithelium [
11]. We cannot explain why elevated levels of uPAR in our model are associated with improved wound healing. However, since previous studies were conducted on primary bronchial epithelial cells from asthma patients, it is likely that functional and immunological properties may have differed from the commercial cell line of alveolar origin that was used in the current study. It is also important to keep in mind that improved gap closure in vitro could cause events related to tissue remodelling and altered wound healing in an in vivo setting.
Regarding the serine protease ligand uPA, we could detect a decrease of uPA, but not significant, in the tryptase stimulated cells compared to both chymase- and non-stimulated cells. Since it has been shown that tryptase via proteolytic activity is a potent activator of the zymogen form of uPA and that blocking of the uPA-uPAR system attenuates tissue repair [
11,
16], this could be a possible explanation for the decreased levels of uPA and the enhanced gap closure activity in our investigation. Given the fact that tryptase just like uPA is a serine protease, this makes tryptase an interesting target for future investigation of its role and interaction with the uPA-uPAR system.
In our study, tryptase and chymase displayed distinct effects on cellular function but also on expression patterns of mediators of the plasminogen activation system. Since tryptase and chymase might be simultaneously released by certain MC phenotypes, one can speculate on an enhanced dual effect: tryptase induces increased expression of the membrane bound receptor uPAR while chymase cleaves it to its soluble form, suPAR. Previous studies have found increased uPAR expression in the bronchial epithelium of patients with asthma. The study also showed that cells with elevated levels of uPAR showed diminished repair via sequestration of uPA by soluble uPAR [
11]. Since the tryptase and chymase expressing MC phenotype, MC
TC, increases in patients with chronic respiratory disease, including in the alveolar parenchyma in patients with asthma [
2], it might indicate a detrimental role for MC
TC with regard to uPAR/suPAR expression and release. In depth studies of this pathway in primary cells and in vivo settings is, therefore, needed to fully understand the potential role of MC
TC in this context.
It is important to understand the results in our study from the perspective of MC plasticity, where both cellular origin and in vitro environment most likely have great influence on protease activity and cellular effects. The protease working concentration in our study was set at 0.5µg/mL for both tryptase and chymase. These doses were established since they were non-toxic, while yet displaying an impact on alveolar functional properties. Nevertheless, it should be emphasized that it is difficult to establish a concentration that would be representative for in vivo release considering that MCs contain a high content of bioactive products with a powerful impact on the surrounding environment [
28]. The A549 cell line we worked with in the current study has been used earlier to study MC interaction as well as remodelling in an asthma context [
29,
30]. However, the true and complete nature of the human MC and its proteases in relation to alveolar epithelial cells remains unanswered; hence, future investigations, in clinically relevant settings, are warranted.

 ,
, 





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}