
Isolation of Human Osteoblast Cells Capable for Mineralization and Synthetizing Bone-Related Proteins In Vitro from Adult Bone

 , , ,
, , ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients
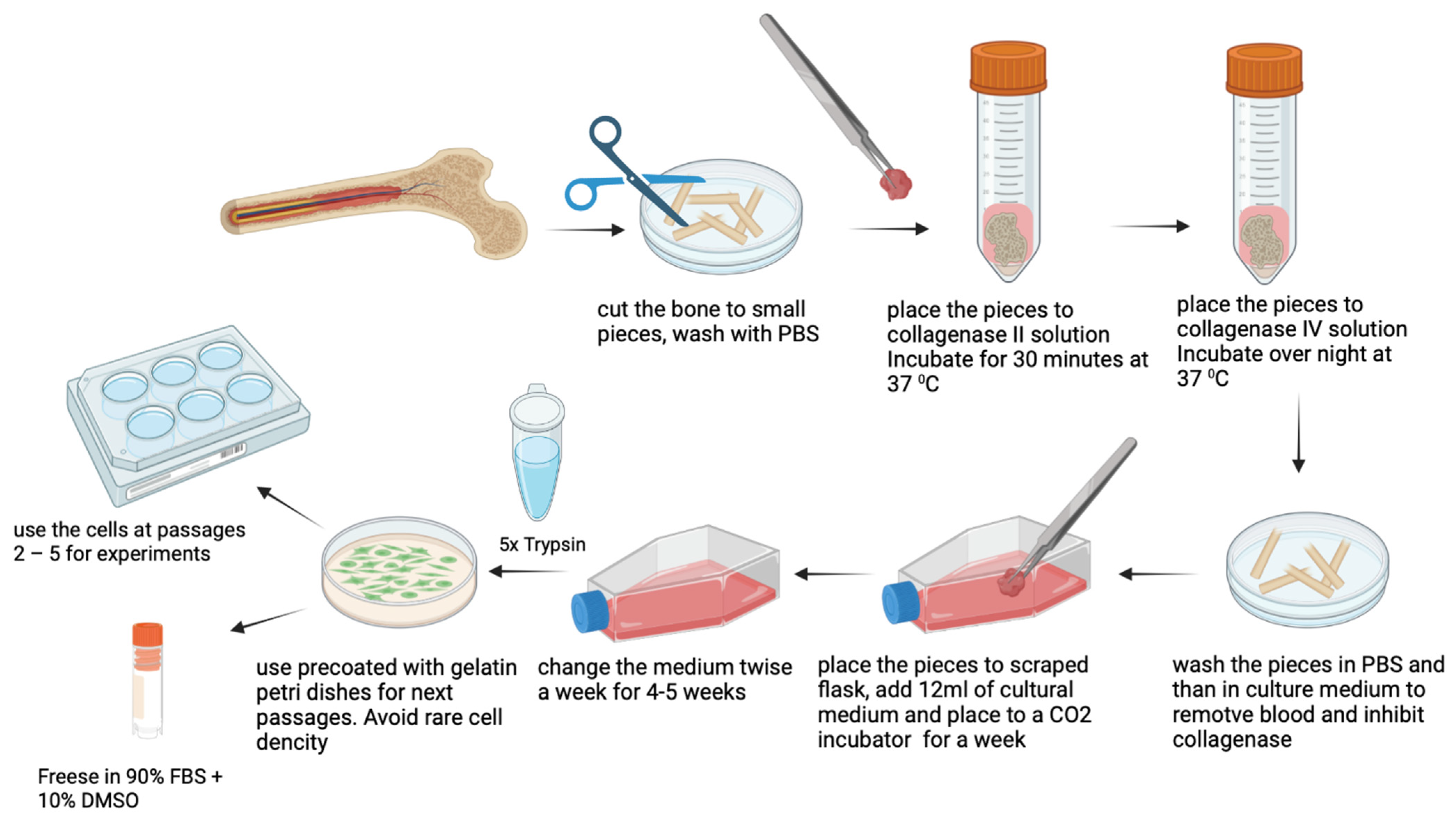
2.2. Protocol of Human OB Cells Isolation from Femur
2.2.1. Reagents and Instruments Preparation
- Autoclave the following items: tissue forceps, steel nippers, scalpel blade, and 0.5 L of PBS (potassium buffer solution). Add to PBS solution (Biolot, Russia) of penicillin/streptomycin (Gibco BRL, Invitrogen, Grand Island, NY, US) in the concentration recommended by the producer.
- Prepare collagenase type II (Worthington Biochemical Corporation, USA): solve collagenase in the volume of DMEM (Gibco BRL, Invitrogen, Grand Island, NY, US) enough to cover the piece of bone (6 mL should be enough for 1 piece of bone at about 3 × 3 cm2), and the final concentration of collagenase II should be 2 mg/mL. Mix well and sterilize the solution by passing through a 0.45 μm filter, then add penicillin/streptomycin in the concentration recommended by the producer. Prepare the solution just before use. Prewarm the solution to 37 °C before use.
- Prepare collagenase type IV the same way as collagenase type II. Final concentration of collagenase IV (Worthington Biochemical Corporation, USA) should be 2 mg/mL. Prewarm the solution to 37 °C before use.
- Prepare 50 mM (1000x) Ascorbic Acid (Sigma Aldrich, St. Louis, MO, USA) solution: solve Ascorbic Acid in water, filter through a 0.45 μm filter and store at −20 in aliquots.
- OB growth medium: DMEM base medium supplemented with 4.5 g/L glucose 2 mM L-glutamine, sodium pyruvate and penicillin/streptomycin (100 mg/L) (Gibco BRL, Invitrogen, Grand Island, NY, US), 15% FBS (fetal bovine serum, Hy Clone, Cytiva, USA), Ascorbic Acid (0.05 mM). Warm to 37 °C just before use.
2.2.2. Osteoblasts Cells (OB) Isolation
- Place a piece of bone in a new 50 mL falcon and wash with PBS supplied with an antibiotic.
- Place the piece of bone in a Petri dish and add PBS with an antibiotic. Remove extraneous soft connective tissue from the outer surfaces of the bone by scraping it with a sterile scalpel blade. Change the PBS solution to avoid contamination with blood cells.
- Cut the fragment of bone into small (not bigger than 2 mm2) pieces with steel nippers. Change the PBS solution as many times as it is needed to wash the small bone pieces from blood and fat.
- Using the tissue forceps, place the small bone pieces into a new 50 mL falcon and add 6 mL of collagenase type II solution. Incubate for 30 min at 37 °C.
- Remove collagenase type II solution by serological pipette and add 6 mL of collagenase type IV solution. Incubate overnight at 37 °C. After treatment with collagenase II only, it is almost impossible to remove the remnants of blood cells from the bone tissue, while step-by-step treatment with 2 different collagenases makes it feasible.
- Remove collagenase type IV solution by serological pipette and wash the resulting small bone pieces with PBS solution. After collagenase treatment, the pieces will become macroscopically white as all the remnants of bone marrow will be completely removed. Then inactivate the collagenase by washing with 5–6 mL of growth culture medium, supplied with 2–10% of FBS.
- Scrape the cultural flask using tissue forceps. Make 4–6 scrapes. Scrapes are needed for anchoring the pieces of bone on the flask surface. It is sufficient for cell attachment and migration from explants. At the same time, it is necessary to leave enough space for intact plastic for cell growth.
- Add 12 mL of prewarmed cultural medium (37 °C) to the T-75 flask, place the bone pieces inside and distribute evenly on the surface of the flask.
- Place to CO2 incubator and do not move the flask for a week to avoid shift of bone explants.
- At day 7 check if cells are visible and change the growth medium. Then change the medium twice a week until the cells become confluent. A representative picture of OB cell isolation is represented in Figure 2.
2.2.3. Osteoblasts Cells (OB) Expansion and Storage
- When OB cells reach confluency of more than 90%, wash them twice with PBS to remove the excess media and detach the cells by adding the appropriate volume of pre-warmed 5x trypsin reagent (prepare from 10x Gibco Trypsin by solving with PBS) to cover the flask surface. Incubate at 37 °C for 3–7 min. After detaching the cells, inactivate trypsin with a culture medium. Centrifuge cells suspension at 180× g for 5 min to remove trypsin.
- Seed the cells on a gelatin-coated flask (0.02% gelatin solved in PBS) or Petri dish at a density of 1200–1500 cells per mm2. Avoid rare density as OB stop proliferating and change their morphology without cell-to-cell contacts. It is recommended to seed OB cells 1:1 in passage 1 and 1:2 or 1:3 in the next passages. OB cells grow rather slowly and rich confluence in a week. Use the cells between passages 2–5.
- Cryoconservation: Remove cells from the Petri dish as described in step 1. After trypsinization and centrifugation resuspend the cell pellet in the previously prepared mix: 90% of FBS plus 10% of DMSO (Dimethyl sulfoxide Sigma Aldrich, St. Louis, MO, USA) to a cell density of ~1 million cells per 1 mL. Add each ml of the cell suspension to a cryopreservation vial. Place vials in a cell-freezing container. Place the container at −80 °C for 6–72 h or according to the freezing container protocol. Remove vials from −80 °C and transfer to liquid nitrogen for long-term storage.
- For all experiments cells passages between 2–5 are used. After passage 5 the cells usually lose osteogenic potential.
2.3. Osteogenic Differentiation
2.4. Western blotting
2.5. Reverse Transcription-PCR
2.6. Immunocytochemistry
2.7. Flow Cytometry
2.8. Alizarin Staining
2.9. Statistical Analysis
3. Results
3.1. OB Cell Cultures Characteristics
3.2. Comparison of OB Cells Properties in Control Conditions and in Osteogenic Medium
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name of Material/ Equipment | Company | Catalog Number |
|---|---|---|
| 7-AAD Viability Dye | Beckman Coulter | A07704 |
| Alizarin Red | Sigma | TMS-008-C |
| beta-glycerolphosphate | Sigma | G9422-100G |
| BMP-2/4 | Santa Cruz | sc-9003 |
| BSA assay kit | Thermo Scientific | PI23235 |
| CD105-PC7 | Beckman Coulter | B43293 |
| beta-glycerolphosphate | Sigma | G9422-100G |
| BMP-2/4 | Santa Cruz | sc-9003 |
| BSA assay kit | Thermo Scientific | PI23235 |
| CD105-PC7 | Beckman Coulter | B43293 |
| CD117-PE | Beckman Coulter | IM2732U |
| CD14-PC5 | Beckman Coulter | IM2640U |
| CD34-FITC | Beckman Coulter | IM1870U |
| CD44-FITC | Beckman Coulter | IM1219U |
| CD45-PC7 | Beckman Coulter | IM3548U |
| CD73-PE | Beckman Coulter | B68176 |
| CD90-PC5 | Beckman Coulter | IM3703 |
| collagenase type II | Worthington | LS004174 |
| collagenase type IV | Worthington | LS004186 |
| dexamethasone | Sigma | D4902 |
| DMEM, high glucose, pyruvate | Gibco | 11995065 |
| DMSO | Sigma | D2650-100ML |
| FBS | Hy Clone | |
| gelatin | Sigma | G1890-100G |
| L-glutamine | Gibco | 25030081 |
| MMLV RT kit | Eurogen | SK021 |
| osteocalcin | Abcam | ab93876 |
| osteopontin | Santa Cruz | sc-21742 |
| penicillin/streptomycin | Gibco | 15140122 |
| protease inhibitor cocktail | Sigma | 4693116001 |
| RIPA Buffer | Sigma | D4902 |
| RUNX2 | Abcam | ab76956 |
| SYBR Green | Eurogen | PB025S |
| Trizol reagent | Eurogen | BC032 |
| Confocal microscope | Olimpus | FV3000 |
| Flow cytometer | Navios | B86672 |
| Light Cycler 96 instrument | Roche | 5815916001 |
| Varioscan | Thermo Scientific | VL0000D0 |
Appendix B
| Gene | ||
|---|---|---|
| ACTA2 | forward | GTTACTACTGCTGAGCGTGAG |
| reverce | CAGGCAACTCGTAACTCTTC | |
| BGLAP | forward | CCTCACACTCCTCGCCCTAT |
| reverce | CTTGGACACAAAGGCTGCAC | |
| BMP2 | forward | GCCAAGCCGAGCCAACAC |
| reverce | CCCACTCGTTTCTGGTAGTTCTTC | |
| Col1A1 | forward | GACCTAAAGGTGCTGCTGGAG |
| reverce | CTTGTTCACCTCTCTCGCCA | |
| ENPP1 | TaqMan | |
| PERIOSTIN | forward | CCCAGCAGTTTTGCCCATT |
| reverce | TGTGGTGGCTCCCACGAT | |
| RUNX2 | forward | TGGATCACCTGAAATGCTG |
| reverce | CGAAATCCCAAACTCCGATA | |
| SPP1 | forward | TCACCTGTGCCATACCAGTTAAA |
| reverce | TGGGTATTTGTTGTAAAGCTGCTT |
References
- Grabowski, P. Physiology of bone. In Calcium and Bone Disorders in Children and Adolescents; Karger: Basel, Switzerland, 2015; Volume 28, pp. 33–55. [Google Scholar]
- Ytteborg, E.; Torgersen, J.; Baeverfjord, G.; Takle, H. The Atlantic salmon (Salmo salar) vertebra and cellular pathways to vertebral deformities. Health Environ. Aquac. 2012, 329–358. [Google Scholar]
- Hayashi, M.; Ono, T.; Nakashima, T. Signaling in Osteoblast Differentiation. In Encyclopedia of Bone Biology; Zaidi, M., Ed.; Academic Press: Oxford, UK, 2020; pp. 416–426. [Google Scholar]
- Czekanska, E.; Stoddart, M.; Richards, R.; Hayes, J. In search of an osteoblast cell model for in vitro research. Eur. Cell Mater. 2012, 24, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Rickard, D.J.; Kassem, M.; Hefferan, T.E.; Sarkar, G.; Spelsberg, T.C.; Riggs, B.L. Isolation and characterization of osteoblast precursor cells from human bone marrow. J. Bone Miner. Res. 1996, 11, 312–324. [Google Scholar] [CrossRef] [PubMed]
- Ochiai-Shino, H.; Kato, H.; Sawada, T.; Onodera, S.; Saito, A.; Takato, T.; Shibahara, T.; Muramatsu, T.; Azuma, T. A novel strategy for enrichment and isolation of osteoprogenitor cells from induced pluripotent stem cells based on surface marker combination. PLoS ONE 2014, 9, e99534. [Google Scholar] [CrossRef] [Green Version]
- Kartsogiannis, V.; Ng, K.W. Cell lines and primary cell cultures in the study of bone cell biology. Mol. Cell. Endocrinol. 2004, 228, 79–102. [Google Scholar] [CrossRef]
- Fujita, K.; Roforth, M.M.; Atkinson, E.; Peterson, J.; Drake, M.; McCready, L.; Farr, J.; Monroe, D.; Khosla, S. Isolation and characterization of human osteoblasts from needle biopsies without in vitro culture. Osteoporos. Int. 2014, 25, 887–895. [Google Scholar] [CrossRef] [Green Version]
- Jonsson, K.B.; Frost, A.; Nilsson, O.; Ljunghall, S.; Ljunggren, Ö. Three isolation techniques for primary culture of human osteoblast-like cells: A comparison. Acta Orthop. Scand. 1999, 70, 365–373. [Google Scholar] [CrossRef]
- Gartland, A.; Rumney, R.M.H.; Dillon, J.P.; Gallagher, J.A. Isolation and culture of human osteoblasts. In Human Cell Culture Protocols; Springer: Berlin/Heidelberg, Germany, 2012; pp. 337–355. [Google Scholar]
- Jaiswal, N.; Haynesworth, S.E.; Caplan, A.I.; Bruder, S.P. Osteogenic differentiation of purified, culture-expanded human mesenchymal stem cells in vitro. J. Cell. Biochem. 1997, 64, 295–312. [Google Scholar] [CrossRef]
- Langenbach, F.; Handschel, J. Effects of dexamethasone, ascorbic acid and β-glycerophosphate on the osteogenic differentiation of stem cells in vitro. Stem Cell Res. Ther. 2013, 4, 117. [Google Scholar] [CrossRef] [Green Version]
- Mildmay-White, A.; Khan, W. Cell surface markers on adipose-derived stem cells: A systematic review. Curr. Stem Cell Res. Ther. 2017, 12, 484–492. [Google Scholar] [CrossRef]
- Zoch, M.L.; Clemens, T.L.; Riddle, R.C. New insights into the biology of osteocalcin. Bone 2016, 82, 42–49. [Google Scholar] [CrossRef]
- Rutkovskiy, A.; Stensløkken, K.-O.; Vaage, I.J. Osteoblast Differentiation at a Glance. Med. Sci. Monit. Basic Res. 2016, 22, 95. [Google Scholar] [CrossRef] [Green Version]
- Gorski, J.P. Biomineralization of bone: A fresh view of the roles of non-collagenous proteins. Front. Biosci. (Landmark Ed.) 2011, 16, 2598. [Google Scholar] [CrossRef] [Green Version]
- Harmey, D.; Hessle, L.; Narisawa, S.; Johnson, K.A.; Terkeltaub, R.; Millán, J.L. Concerted regulation of inorganic pyrophosphate and osteopontin by akp2, enpp1, and ank: An integrated model of the pathogenesis of mineralization disorders. Am. J. Pathol. 2004, 164, 1199–1209. [Google Scholar] [CrossRef]
- Lobov, A.; Malashicheva, A. Osteogenic differentiation: A universal cell program of heterogeneous mesenchymal cells or a similar extracellular matrix mineralizing phenotype? Biol. Commun. 2022, 67, 32–48. [Google Scholar] [CrossRef]
- Bernhardt, A.; Wolf, S.; Weiser, E.; Vater, C.; Gelinsky, M. An improved method to isolate primary human osteocytes from bone. Biomed. Eng. Biomed. Tech. 2020, 65, 107–111. [Google Scholar] [CrossRef]
- Shah, M.; Gburcik, V.; Reilly, P.; Sankey, R.; Emery, R.; Clarkin, C.; Pitsillides, A. Local origins impart conserved bone type-related differences in human osteoblast behaviour. Eur. Cells Mater. 2015, 29, 155–176. [Google Scholar] [CrossRef]
- Bourne, L.E.; Wheeler-Jones, C.P.; Orriss, I.R. Regulation of mineralisation in bone and vascular tissue: A comparative review. J. Endocrinol. 2021, 248, R51–R65. [Google Scholar] [CrossRef]
- Bonnet, N.; Standley, K.N.; Bianchi, E.N.; Stadelmann, V.; Foti, M.; Conway, S.J.; Ferrari, S.L. The matricellular protein periostin is required for sost inhibition and the anabolic response to mechanical loading and physical activity. J. Biol. Chem. 2009, 284, 35939–35950. [Google Scholar] [CrossRef] [Green Version]
- Massy, E.; Rousseau, J.; Gueye, M.; Bonnelye, E.; Brevet, M.; Chambard, L.; Duruisseaux, M.; Borel, O.; Roger, C.; Guelminger, R. Serum total periostin is an independent marker of overall survival in bone metastases of lung adenocarcinoma. J. Bone Oncol. 2021, 29, 100364. [Google Scholar] [CrossRef]
- Yagiela, J.A.; Woodbury, D.M. Enzymatic isolation of osteoblasts from fetal rat calvaria. Anat. Rec. 1977, 188, 287–305. [Google Scholar] [CrossRef] [PubMed]
- Patel, J.J.; Bourne, L.E.; Davies, B.K.; Arnett, T.R.; MacRae, V.E.; Wheeler-Jones, C.P.; Orriss, I.R. Differing calcification processes in cultured vascular smooth muscle cells and osteoblasts. Exp. Cell Res. 2019, 380, 100–113. [Google Scholar] [CrossRef] [PubMed]
- Kostina, A.; Lobov, A.; Semenova, D.; Kiselev, A.; Klausen, P.; Malashicheva, A. Context-Specific Osteogenic Potential of Mesenchymal Stem Cells. Biomedicines 2021, 9, 673. [Google Scholar] [CrossRef] [PubMed]
- Ignatieva, E.; Kostina, D.; Irtyuga, O.; Uspensky, V.; Golovkin, A.; Gavriliuk, N.; Moiseeva, O.; Kostareva, A.; Malashicheva, A. Mechanisms of Smooth Muscle Cell Differentiation Are Distinctly Altered in Thoracic Aortic Aneurysms Associated with Bicuspid or Tricuspid Aortic Valves. Front. Physiol. 2017, 8, 536. [Google Scholar] [CrossRef] [Green Version]
- Kostina, A.; Shishkova, A.; Ignatieva, E.; Irtyuga, O.; Bogdanova, M.; Levchuk, K.; Golovkin, A.; Zhiduleva, E.; Uspenskiy, V.; Moiseeva, O. Different Notch signaling in cells from calcified bicuspid and tricuspid aortic valves. J. Mol. Cell. Cardiol. 2018, 114, 211–219. [Google Scholar] [CrossRef]
- Bruderer, M.; Richards, R.; Alini, M.; Stoddart, M.J. Role and regulation of RUNX2 in osteogenesis. Eur. Cell Mater. 2014, 28, 269–286. [Google Scholar] [CrossRef]






| Donor | Bigger Cell Size,% | Cell Viability,% | CD 90,% | CD 105,% | CD 73,% | CD 44,% | CD14, CD45, CD34, CD117, HLA-DR |
|---|---|---|---|---|---|---|---|
| Donor 1 | P2-12 | P2-95.2 | P2-99 | P2-100 | P2-99.8 | P2-99.2 | All negative |
| P4–13 | P4-91.3 | P4-99.5 | P4-100 | P4-99.5 | P4-98.6 | ||
| Donor 2 | P2-38 | P2-86.8 | P2-98.5 | P2-99.7 | P2-98.1 | P2-98.7 | All negative |
| P4-22 | P4-89.9 | P4-99 | P4-99.7 | P4-99.2 | P4-99.1 | ||
| Donor 3 | P2-18 | P2-89.8 | P2-97.4 | P2-99.9 | P2-99.2 | P2-99.6 | All negative |
| P4-5 | P4-91.3 | P4-98.8 | P4-99.9 | P4-99.2 | P4-98.6 | ||
| Donor 4 | P2-41 | P2-89.4 | P2-98.2 | P2-99.6 | P2-99.3 | P2-99.6 | All negative |
| Donor 5 | P2-37 | P2-95.3 | P2-98.4 | P2-99.9 | P2-99.5 | P2-99.7 | All negative |
| Median ± st deviation | P2-37 ± 12 | P2-90 ± 3.4 | P2-98.5 ± 0.5 | P2-99.9 ± 0.1 | P2-99.3 ± 0.6 | P2-99.6 ± 0.4 | |
| P4-13 ± 7 | P4-91 ± 0.95 | P4-99 ± 0.3 | P4-99.9 ± 0.1 | P4-99.2 ± 0.2 | P4-98.6 ± 0.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kostina, D.; Lobov, A.; Klausen, P.; Karelkin, V.; Tikhilov, R.; Bozhkova, S.; Sereda, A.; Ryumina, N.; Enukashvily, N.; Malashicheva, A. Isolation of Human Osteoblast Cells Capable for Mineralization and Synthetizing Bone-Related Proteins In Vitro from Adult Bone. Cells 2022, 11, 3356. https://doi.org/10.3390/cells11213356
Kostina D, Lobov A, Klausen P, Karelkin V, Tikhilov R, Bozhkova S, Sereda A, Ryumina N, Enukashvily N, Malashicheva A. Isolation of Human Osteoblast Cells Capable for Mineralization and Synthetizing Bone-Related Proteins In Vitro from Adult Bone. Cells. 2022; 11(21):3356. https://doi.org/10.3390/cells11213356
Chicago/Turabian StyleKostina, Daria, Arseniy Lobov, Polina Klausen, Vitaly Karelkin, Rashid Tikhilov, Svetlana Bozhkova, Andrey Sereda, Nadezhda Ryumina, Natella Enukashvily, and Anna Malashicheva. 2022. "Isolation of Human Osteoblast Cells Capable for Mineralization and Synthetizing Bone-Related Proteins In Vitro from Adult Bone" Cells 11, no. 21: 3356. https://doi.org/10.3390/cells11213356
APA StyleKostina, D., Lobov, A., Klausen, P., Karelkin, V., Tikhilov, R., Bozhkova, S., Sereda, A., Ryumina, N., Enukashvily, N., & Malashicheva, A. (2022). Isolation of Human Osteoblast Cells Capable for Mineralization and Synthetizing Bone-Related Proteins In Vitro from Adult Bone. Cells, 11(21), 3356. https://doi.org/10.3390/cells11213356