

Effective, Long-Term, Neutrophil Depletion Using a Murinized Anti-Ly-6G 1A8 Antibody
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mice
2.2. Neutrophil Depletion
2.3. Flow Cytometry
2.4. Neutrophil Attraction Model in IMR32 Tumor Bearing Mice
2.5. Quantification and Statistical Analysis
3. Results
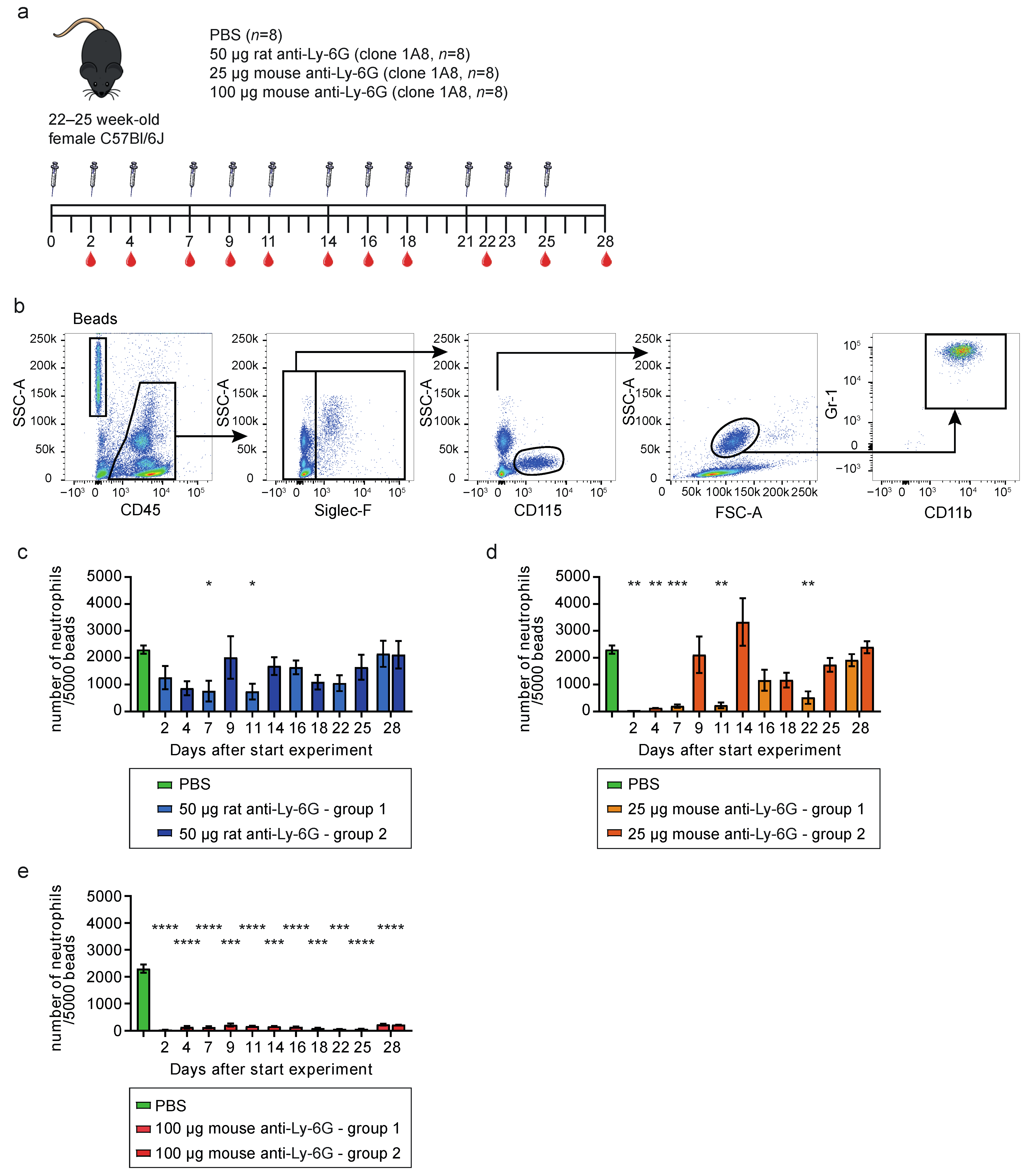
3.1. Mouse-Ly-6G Depletes Neutrophils More Efficiently Than Rat-Ly-6G in C57Bl6/J Mice
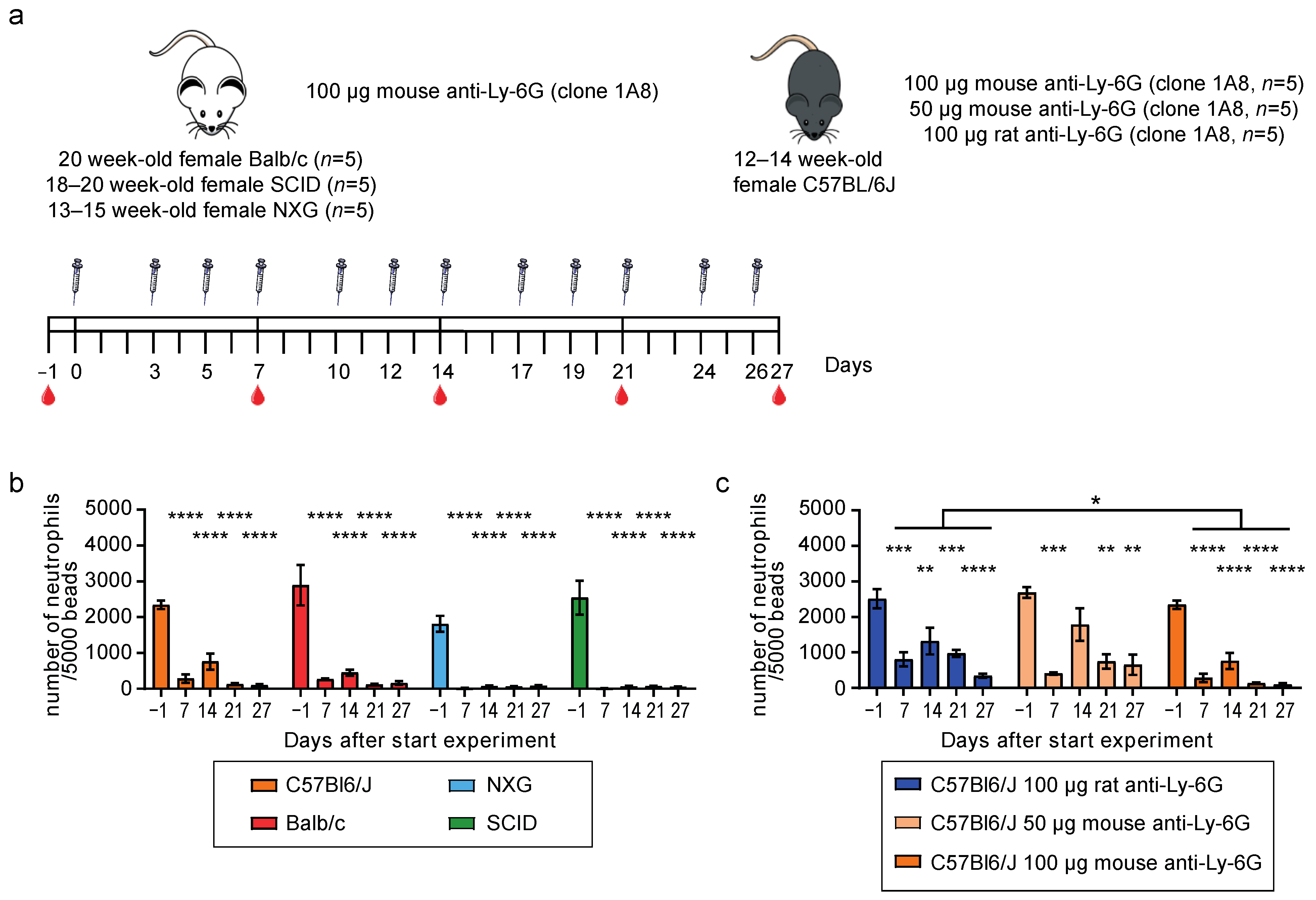
3.2. 100μg Mouse-Ly-6G Efficiently Depletes Neutrophils in C57Bl6/J, Balb/c, NXG, and SCID Mice
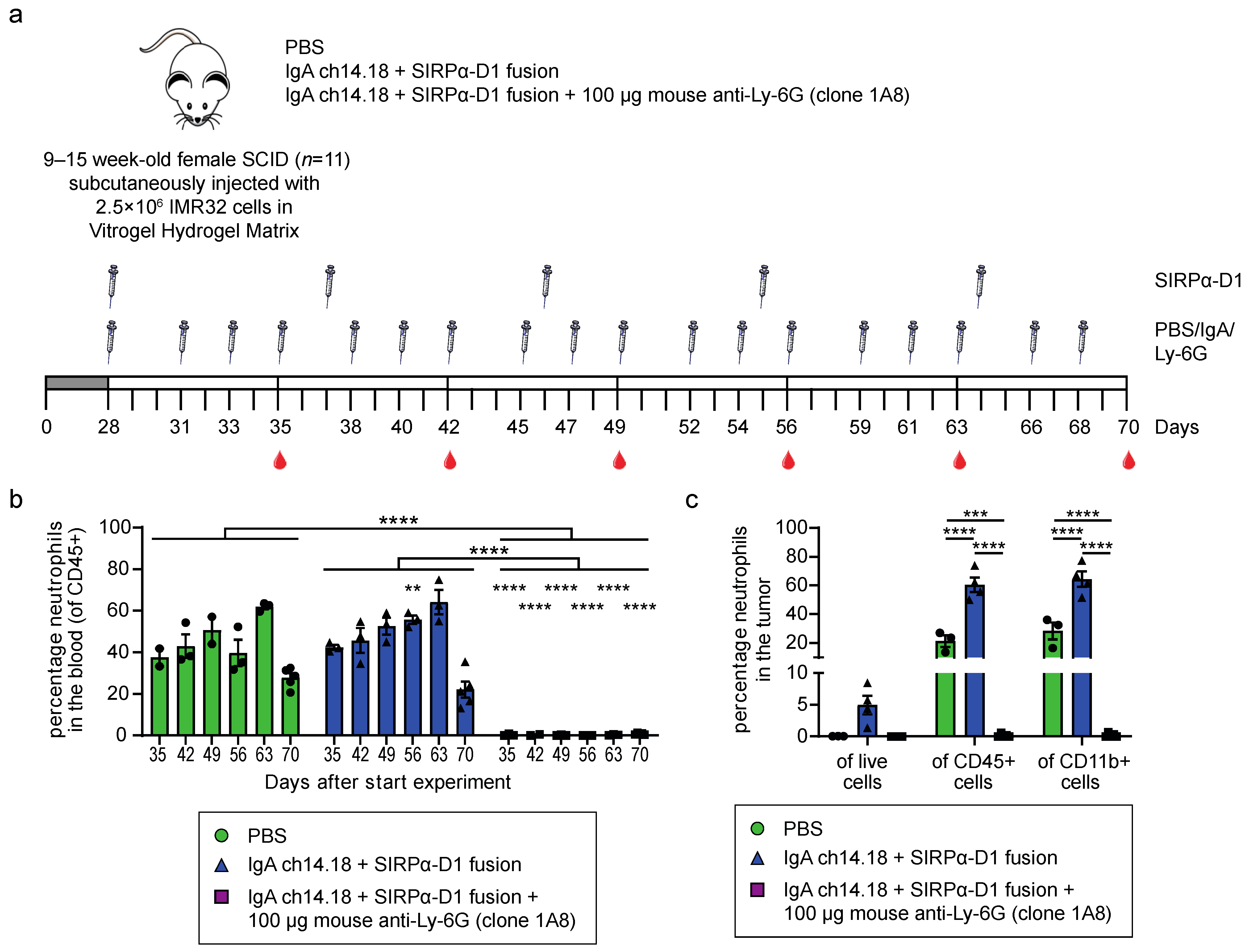
3.3. Neutrophils Can Efficiently Be Depleted Intratumorally in IMR32 Tumor Bearing Mice
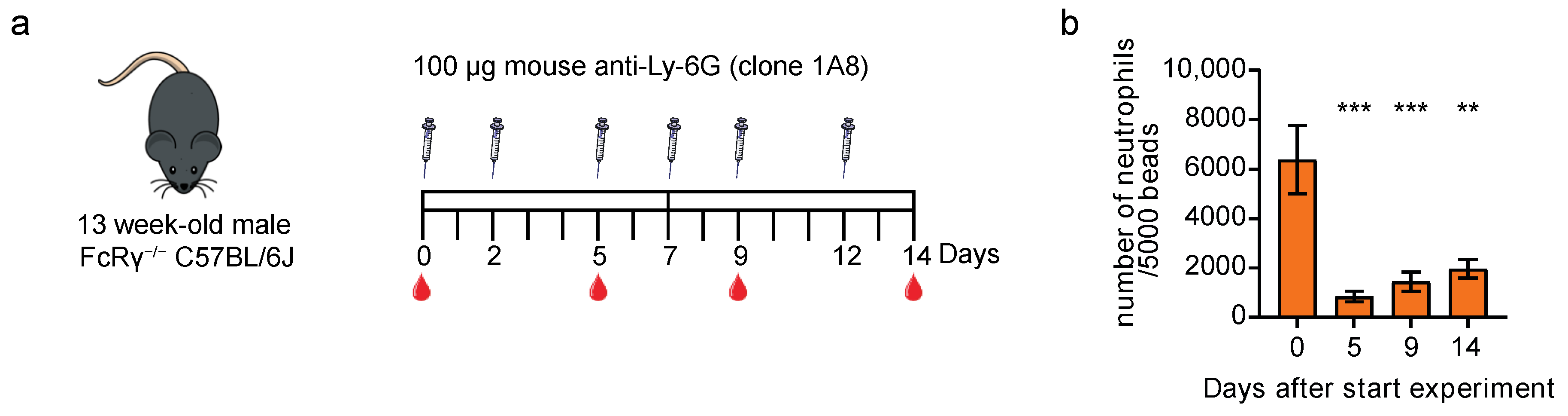
3.4. Neutrophil Depletion with Mouse-Ly-6G Is Not Solely Complement- or Fc Receptor-Mediated
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Masucci, M.T.; Minopoli, M.; Carriero, M.V. Tumor Associated Neutrophils. Their Role in Tumorigenesis, Metastasis, Prognosis and Therapy. Front. Oncol. 2019, 9, 1146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Musiani, P.; Allione, A.; Modica, A.; Lollini, P.L.; Giovarelli, M.; Cavallo, F.; Belardelli, F.; Forni, G.; Modesti, A. Role of neutrophils and lymphocytes in inhibition of a mouse mammary adenocarcinoma engineered to release IL-2, IL-4, IL-7, IL-10, IFN-alpha, IFN-gamma, and TNF-alpha. Lab. Investig. 1996, 74, 146–157. [Google Scholar] [PubMed]
- Schmielau, J.; Finn, O.J. Activated granulocytes and granulocyte-derived hydrogen peroxide are the underlying mechanism of suppression of t-cell function in advanced cancer patients. Cancer Res. 2001, 61, 4756–4760. [Google Scholar] [PubMed]
- Serafini, P.; Borrello, I.; Bronte, V. Myeloid suppressor cells in cancer: Recruitment, phenotype, properties, and mechanisms of immune suppression. Semin. Cancer Biol. 2006, 16, 53–65. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Greten, T.F. Editorial: “Invisible” MDSC in tumor-bearing individuals after antibody depletion: Fact or fiction? J. Leukoc. Biol. 2016, 99, 794. [Google Scholar] [CrossRef] [Green Version]
- Moses, K.; Klein, J.C.; Männ, L.; Klingberg, A.; Gunzer, M.; Brandau, S. Survival of residual neutrophils and accelerated myelopoiesis limit the efficacy of antibody-mediated depletion of Ly-6G+ cells in tumor-bearing mice. J. Leukoc. Biol. 2016, 99, 811–823. [Google Scholar] [CrossRef] [Green Version]
- Pollenus, E.; Malengier-Devlies, B.; Vandermosten, L.; Pham, T.-T.; Mitera, T.; Possemiers, H.; Boon, L.; Opdenakker, G.; Matthys, P.; Steen, P.E.V.D. Limitations of neutrophil depletion by anti-Ly6G antibodies in two heterogenic immunological models. Immunol. Lett. 2019, 212, 30–36. [Google Scholar] [CrossRef]
- Hestdal, K.; Ruscetti, F.W.; Ihle, J.N.; Jacobsen, S.E.W.; Dubois, C.M.; Kopp, W.C.; Longo, D.L.; Keller, J.R. Characterization and regulation of RB6-8C5 antigen expression on murine bone marrow cells. J. Immunol. 1991, 147, 22–28. [Google Scholar]
- Fleming, T.J.; Fleming, M.L.; Malek, T.R. Selective expression of Ly-6G on myeloid lineage cells in mouse bone marrow. RB6-8C5 mAb to granulocyte-differentiation antigen (Gr-1) detects members of the Ly-6 family. J. Immunol. 1993, 151, 2399–2408. [Google Scholar]
- Daley, J.M.; Thomay, A.A.; Connolly, M.D.; Reichner, J.S.; Albina, J.E. Use of Ly6G-specific monoclonal antibody to deplete neutrophils in mice. J. Leukoc. Biol. 2008, 83, 64–70. [Google Scholar] [CrossRef] [Green Version]
- Morton, J.; Coles, B.; Wright, K.; Gallimore, A.; Morrow, J.D.; Terry, E.S.; Anning, P.B.; Morgan, P.; Dioszeghy, V.; Kühn, H.; et al. Circulating neutrophils maintain physiological blood pressure by suppressing bacteria and IFNγ-dependent iNOS expression in the vasculature of healthy mice. Blood 2008, 111, 5187–5194. [Google Scholar] [CrossRef] [PubMed]
- Abbitt, K.B.; Cotter, M.J.; Ridger, V.C.; Crossman, D.C.; Hellewell, P.G.; Norman, K.E. Antibody ligation of murine Ly-6G induces neutropenia, blood flow cessation, and death via complement-dependent and independent mechanisms. J. Leukoc. Biol. 2009, 85, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Bruhn, K.W.; Dekitani, K.; Nielsen, T.B.; Pantapalangkoor, P.; Spellberg, B. Ly6G-mediated depletion of neutrophils is dependent on macrophages. Results Immunol. 2016, 6, 5–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boivin, G.; Faget, J.; Ancey, P.-B.; Gkasti, A.; Mussard, J.; Engblom, C.; Pfirschke, C.; Contat, C.; Pascual, J.; Vazquez, J.; et al. Durable and controlled depletion of neutrophils in mice. Nat. Commun. 2020, 11, 2762. [Google Scholar] [CrossRef]
- Stackowicz, J.; Jönsson, F.; Reber, L.L. Mouse Models and Tools for the in vivo Study of Neutrophils. Front. Immunol. 2019, 10, 3130. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.; Wu, H.Y.; Wesselschmidt, R.; Kornaga, T.; Link, D.C. Impaired Production and Increased Apoptosis of Neutrophils in Granulocyte Colony-Stimulating Factor Receptor–Deficient Mice. Immunity 1996, 5, 491–501. [Google Scholar] [CrossRef] [Green Version]
- Eash, K.J.; Greenbaum, A.; Gopalan, P.K.; Link, D.C. CXCR2 and CXCR4 antagonistically regulate neutrophil trafficking from murine bone marrow. J. Clin. Investig. 2010, 120, 2423–2431. [Google Scholar] [CrossRef] [Green Version]
- Karsunky, H.; Zeng, H.; Schmidt, T.; Zevnik, B.; Kluge, R.; Schmid, K.W.; Dührsen, U.; Möröy, T. Inflammatory reactions and severe neutropenia in mice lacking the transcriptional repressor Gfi1. Nat. Genet. 2002, 30, 295–300. [Google Scholar] [CrossRef]
- Bruhns, P. Properties of mouse and human IgG receptors and their contribution to disease models. Blood 2012, 119, 5640–5649. [Google Scholar] [CrossRef]
- Leatherbarrow, R.J.; Dwek, R.A. Binding of complement subcomponent Clq to mouse IgGl, IgG2a AND IgG2b: A novel Clq binding assay. Mol. Immunol. 1984, 21, 321–327. [Google Scholar] [CrossRef]
- Boross, P.; Lohse, S.; Nederend, M.; Jansen, J.H.M.; van Tetering, G.; Dechant, M.; Peipp, M.; Royle, L.; Liew, L.P.; Boon, L.; et al. Ig A EGFR antibodies mediate tumour killing in vivo. EMBO Mol. Med. 2013, 5, 1213–1226. [Google Scholar] [CrossRef] [PubMed]
- Takai, T.; Li, M.; Sylvestre, D.; Clynes, R.; Ravetch, J.V. FcR gamma chain deletion results in pleiotrophic effector cell defects. Cell 1994, 76, 519–529. [Google Scholar] [CrossRef]
- Meyer, S.; Nederend, M.; Jansen, J.M.; Reiding, K.R.; Jacobino, S.R.; Meeldijk, J.; Bovenschen, N.; Wuhrer, M.; Valerius, T.; Ubink, R.; et al. Improved in vivo anti-tumor effects of IgA-Her2 antibodies through half-life extension and serum exposure enhancement by FcRn targeting. mAbs 2016, 8, 87–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evers, M.; Rösner, T.; Duenkel, A.; Jansen, J.H.M.M.; Baumann, N.; Broeke, T.T.; Nederend, M.; Eichholz, K.; Klausz, K.; Reiding, K.; et al. The selection of variable regions affects effector mechanisms of IgA antibodies against CD20. Blood Adv. 2021, 5, 3807–3820. [Google Scholar] [CrossRef] [PubMed]
- Chernyavska, M.; Hermans, C.K.; Chan, C.; Baumann, N.; Rösner, T.; Leusen, J.H.; Valerius, T.; Verdurmen, W.P. Evaluation of immunotherapies improving macrophage anti-tumor response using a microfluidic model. Organs-on-a-Chip 2022, 4, 100019. [Google Scholar] [CrossRef]
- Bosma, G.C.; Custer, R.P.; Bosma, M.J. A severe combined immunodeficiency mutation in the mouse. Nature 1983, 301, 527–530. [Google Scholar] [CrossRef]
- Shultz, L.D.; Schweitzer, P.A.; Christianson, S.W.; Gott, B.; Schweitzer, I.B.; Tennent, B.; McKenna, S.; Mobraaten, L.; Rajan, T.V.; Greiner, D.L.; et al. Multiple defects in innate and adaptive immunologic function in NOD/LtSz-scid mice. J. Immunol. 1995, 154, 180–191. [Google Scholar]
- Deyhle, M.R.; Callaway, C.S.; Neyroud, D.; D’Lugos, A.C.; Judge, S.M.; Judge, A.R. Depleting Ly6G Positive Myeloid Cells Reduces Pancreatic Cancer-Induced Skeletal Muscle Atrophy. Cells 2022, 11, 1893. [Google Scholar] [CrossRef]
- Chan, C.; Lustig, M.; Baumann, N.; Valerius, T.; van Tetering, G.; Leusen, J.H.W. Targeting Myeloid Checkpoint Molecules in Combination With Antibody Therapy: A Novel Anti-Cancer Strategy With IgA Antibodies? Front. Immunol. 2022, 13, 932155. [Google Scholar] [CrossRef]
- Sun, D.; Sun, P.; He, S.; Shi, M. Rat IgG mediated circulatory cell depletion in mice requires mononuclear phagocyte system and is facilitated by complement. J. Leukoc. Biol. 2020, 107, 529–539. [Google Scholar] [CrossRef]
- Uchida, J.; Hamaguchi, Y.; Oliver, J.A.; Ravetch, J.V.; Poe, J.C.; Haas, K.M.; Tedder, T.F. The Innate Mononuclear Phagocyte Network Depletes B Lymphocytes through Fc Receptor–dependent Mechanisms during Anti-CD20 Antibody Immunotherapy. J. Exp. Med. 2004, 199, 1659–1669. [Google Scholar] [CrossRef] [PubMed]
- Otten, M.A.; Van Der Bij, G.J.; Verbeek, S.J.; Nimmerjahn, F.; Ravetch, J.V.; Beelen, R.H.J.; Van De Winkel, J.G.J.; Van Egmond, M. Experimental Antibody Therapy of Liver Metastases Reveals Functional Redundancy between Fc gammaRI and Fc gammaRIV. J. Immunol. 2008, 181, 6829–6836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gül, N.; Babes, L.; Bellaire-Siegmund, K.; Korthouwer, R.; Bögels, M.; Braster, R.; Vidarsson, G.; ten Hagen, T.L.M.; Kubes, P.; van Egmond, M. Macrophages eliminate circulating tumor cells after monoclonal antibody therapy. J. Clin. Investig. 2014, 124, 812–823. [Google Scholar] [CrossRef] [PubMed]
- Daëron, M. Fc Receptor Biology. Annu. Rev. Immunol. 1997, 15, 203–234. [Google Scholar] [CrossRef]
- Bugl, S.; Wirths, S.; Radsak, M.P.; Schild, H.; Stein, P.; André, M.C.; Müller, M.R.; Malenke, E.; Wiesner, T.; Märklin, M.; et al. Steady-state neutrophil homeostasis is dependent on TLR4/TRIF signaling. Blood 2013, 121, 723–733. [Google Scholar] [CrossRef] [Green Version]
- Kaplan, M.J. Role of neutrophils in systemic autoimmune diseases. Arthritis Res. Ther. 2013, 15, 219. [Google Scholar] [CrossRef] [Green Version]
- Leusen, J.H. IgA as therapeutic antibody. Mol. Immunol. 2015, 68, 35–39. [Google Scholar] [CrossRef]
- Brandsma, A.M.; Bondza, S.; Evers, M.; Koutstaal, R.; Nederend, M.; Jansen, J.H.M.; Rösner, T.; Valerius, T.; Leusen, J.H.W.; Broeke, T.T. Potent Fc Receptor Signaling by IgA Leads to Superior Killing of Cancer Cells by Neutrophils Compared to IgG. Front. Immunol. 2019, 10, 704. [Google Scholar] [CrossRef] [Green Version]
- Evers, M.; Broeke, T.T.; Jansen, J.M.; Nederend, M.; Hamdan, F.; Reiding, K.R.; Meyer, S.; Moerer, P.; Brinkman, I.; Rösner, T.; et al. Novel chimerized IgA CD20 antibodies: Improving neutrophil activation against CD20-positive malignancies. mAbs 2020, 12, 1795505. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antigen | Species | Isotype | Company | Catalog Number |
|---|---|---|---|---|
| Ly-6G (clone 1A8) | Mouse | IgG2a Kappa | Absolute Antibody | Ab00295-2.0 |
| Ly-6G (clone 1A8) | Rat | IgG2a Kappa | Bio X Cell | BP0075-1_5 mg |
| Antigen | Fluorophore | Company | Catalog Number | Dilution |
|---|---|---|---|---|
| CD11b | Alexa488 | BD | 557672 | 100× |
| GR-1 (clone RB6-8C5) | PE-Cy7 | eBioscience | 25-5931-82 | 200× |
| CD115 | APC | Sony Biotechnology | 1277550 | 200× |
| Ly-6C | APC-Cy7 | Biolegend | 128025 | 200× |
| Siglec-F/CD170 | BV421 | BD | 562681 | 100× |
| CD45 | BV510 | Biolegend | 103137 | 200× |
| Antigen | Fluorophore | Company | Catalog Number | Dilution |
|---|---|---|---|---|
| CD3e | PE | Biolegend | 100308 | 200× |
| NK1.1 | PerCP-Cy5.5 | Biolegend | 108727 | 100× |
| B220/CD45R | PE-Cy7 | BD | 561881 | 100× |
| CD11b | APC | eBioscience | 17-0112-80 | 100× |
| Ly-6C | APC-Cy7 | Biolegend | 128025 | 200× |
| CD45 | Pacific Blue | Biolegend | 103126 | 200× |
| GR-1 (clone RB6-8C5) | Pacific Orange | Invitrogen | RM3030 | 100× |
| Antigen | Fluorophore | Company | Catalog Number | Dilution |
|---|---|---|---|---|
| CD11b | FITC | Pharmingen | 553310 | 50× |
| Ly-6C | PerCP-Cy5.5 | Biolegend | 128012 | 100× |
| Ly-6G | PE-Cy7 | Biolegend | 127618 | 300× |
| Siglec-F/CD170 | BV421 | BD | 562681 | 100× |
| CD45 | BV510 | Biolegend | 103137 | 200× |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Olofsen, P.A.; Stip, M.C.; Jansen, J.H.M.; Chan, C.; Nederend, M.; Tieland, R.G.; Tsioumpekou, M.; Leusen, J.H.W. Effective, Long-Term, Neutrophil Depletion Using a Murinized Anti-Ly-6G 1A8 Antibody. Cells 2022, 11, 3406. https://doi.org/10.3390/cells11213406
Olofsen PA, Stip MC, Jansen JHM, Chan C, Nederend M, Tieland RG, Tsioumpekou M, Leusen JHW. Effective, Long-Term, Neutrophil Depletion Using a Murinized Anti-Ly-6G 1A8 Antibody. Cells. 2022; 11(21):3406. https://doi.org/10.3390/cells11213406
Chicago/Turabian StyleOlofsen, Patricia A., Marjolein C. Stip, J. H. Marco Jansen, Chilam Chan, Maaike Nederend, Ralph G. Tieland, Maria Tsioumpekou, and Jeanette H. W. Leusen. 2022. "Effective, Long-Term, Neutrophil Depletion Using a Murinized Anti-Ly-6G 1A8 Antibody" Cells 11, no. 21: 3406. https://doi.org/10.3390/cells11213406
APA StyleOlofsen, P. A., Stip, M. C., Jansen, J. H. M., Chan, C., Nederend, M., Tieland, R. G., Tsioumpekou, M., & Leusen, J. H. W. (2022). Effective, Long-Term, Neutrophil Depletion Using a Murinized Anti-Ly-6G 1A8 Antibody. Cells, 11(21), 3406. https://doi.org/10.3390/cells11213406