
Role of lncRNA Has2os in Skeletal Muscle Differentiation and Regeneration


Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture and Differentiation Induction
2.2. RNA Extraction and qRT-PCR Assays
2.3. Western Blot
2.4. Gene Knockdown
2.5. Immunofluorescence
2.6. Lucierase Reporter Assays
2.7. Skeletal Muscle Injury Regeneration Model Was Established
2.8. Statistical Analysis and Processing
3. Results
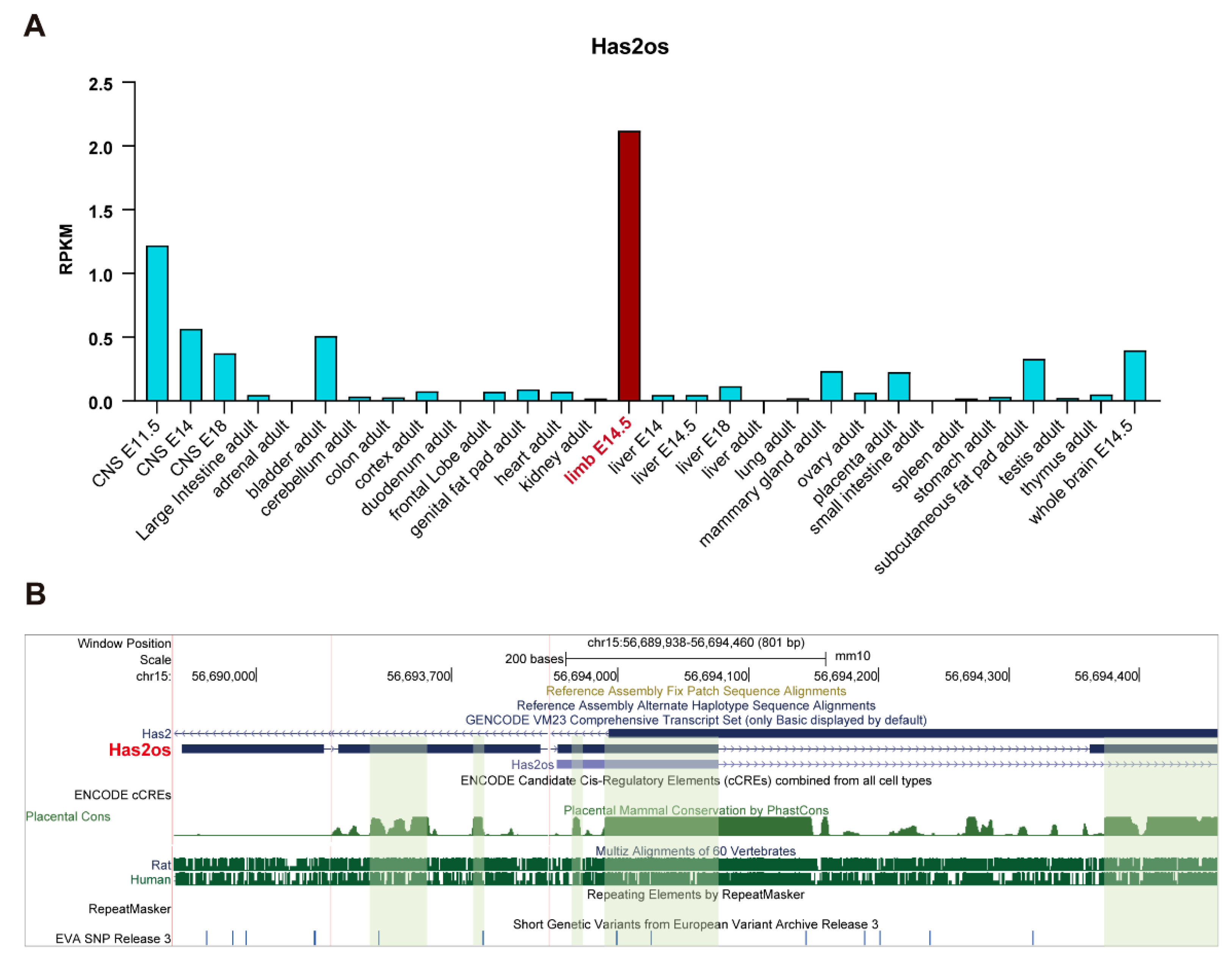
3.1. lncRNA Has2os Is Highly Expressed in Skeletal Muscle
3.2. Expression Pattern of Has2os in Skeletal Muscle Differentiation
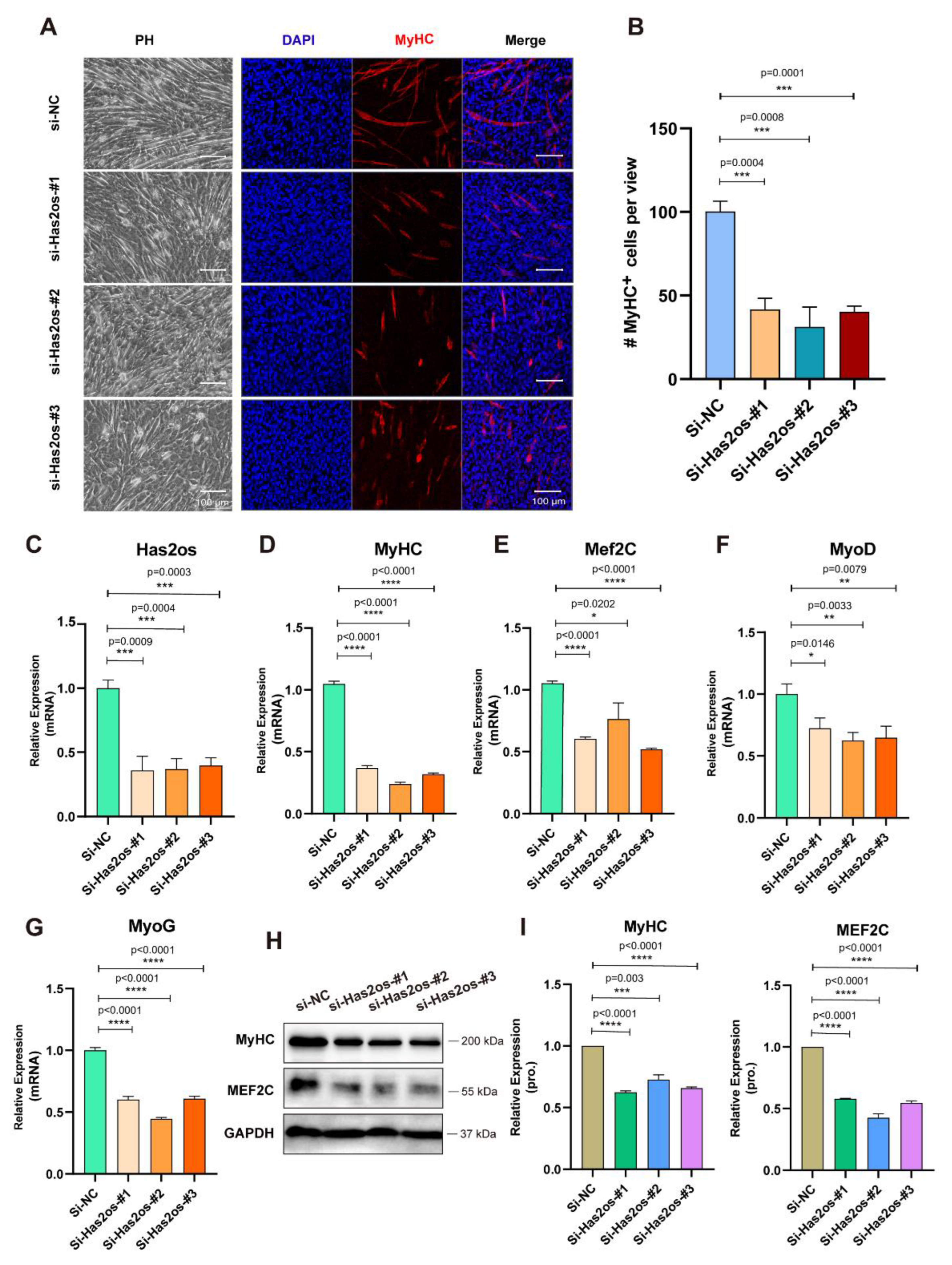
3.3. Knockdown of Has2os Significantly Blocked Muscle Differentiation
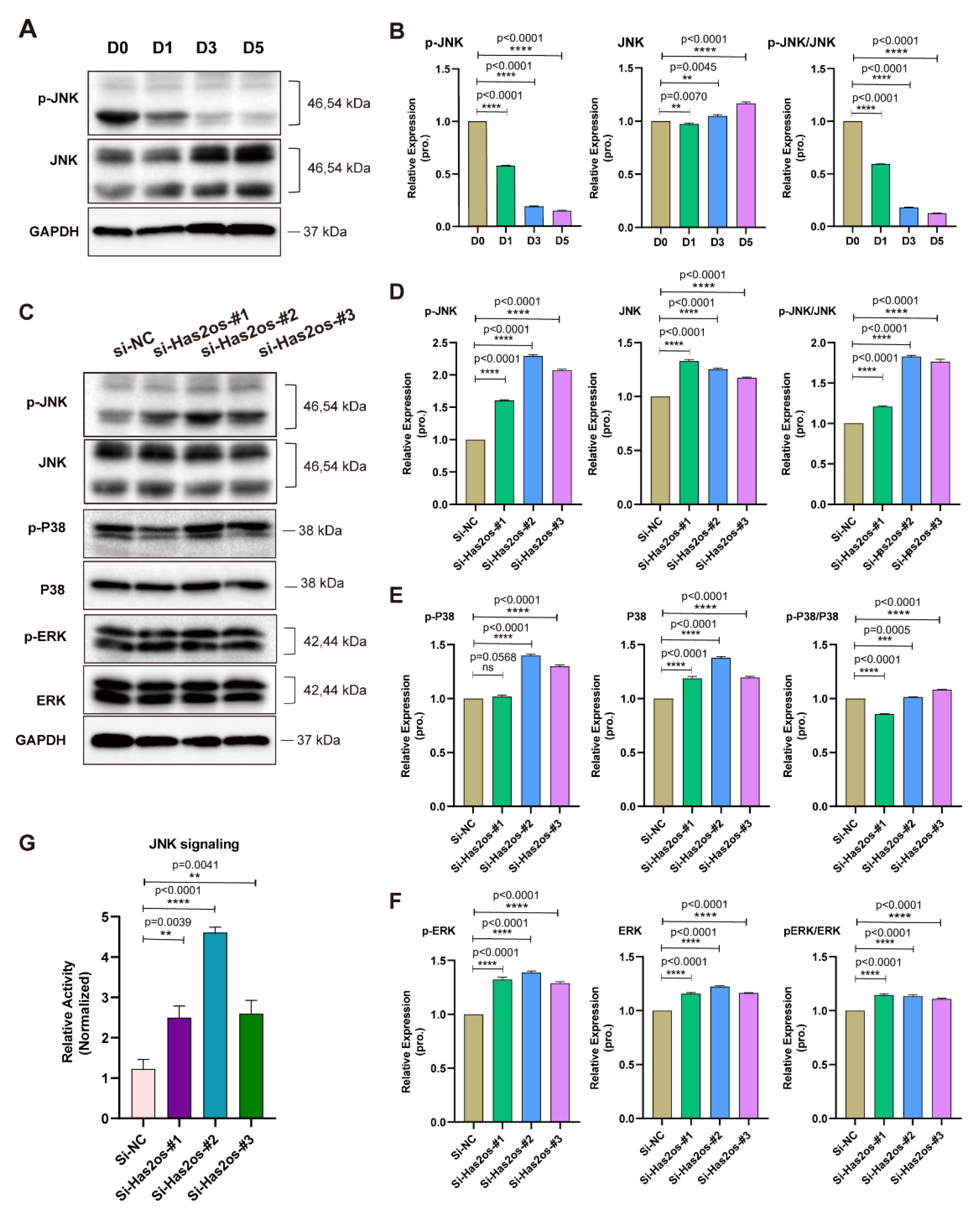
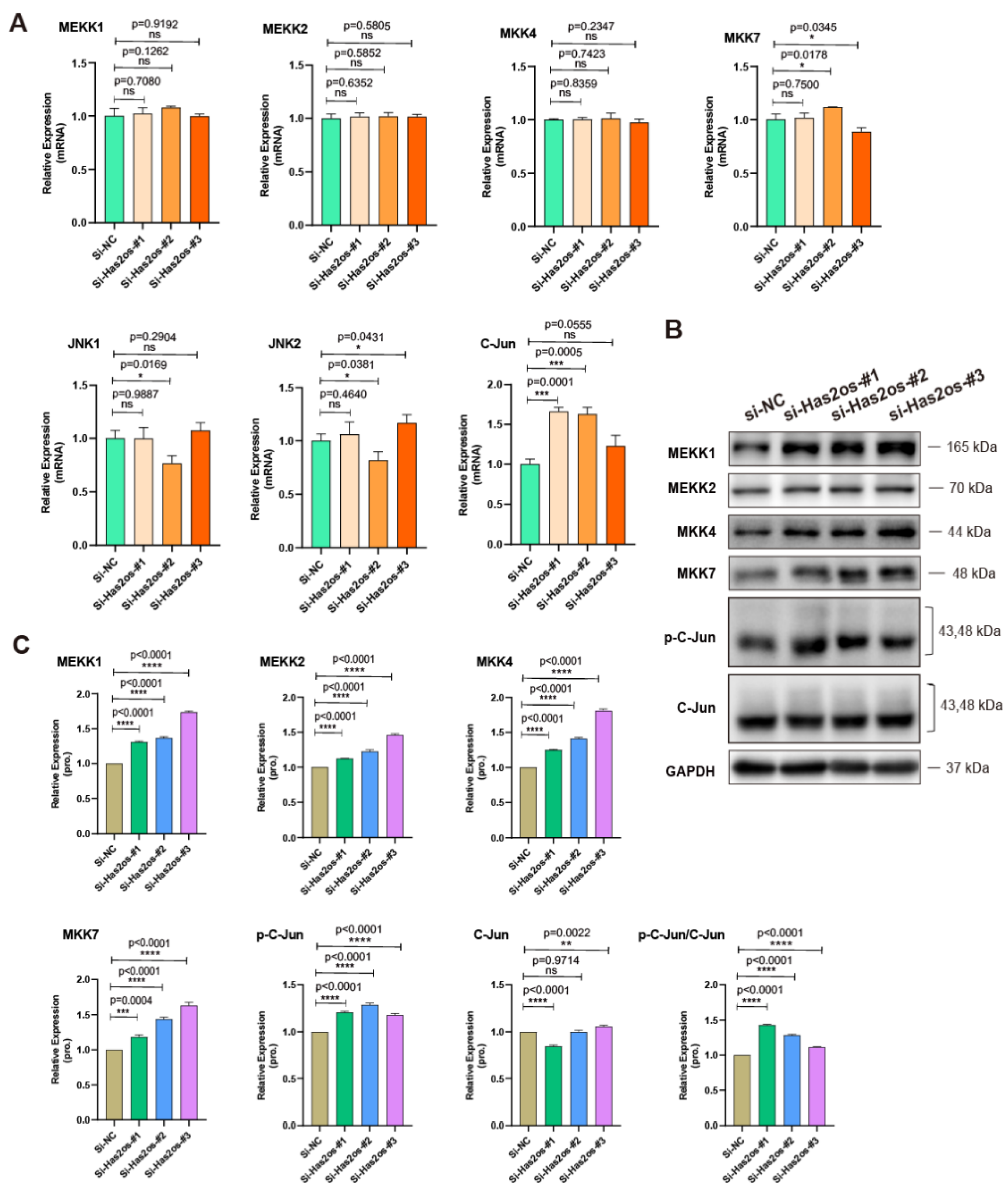
3.4. Has2os Regulates Skeletal Muscle Differentiation through the JNK/MAPK Signaling Pathway
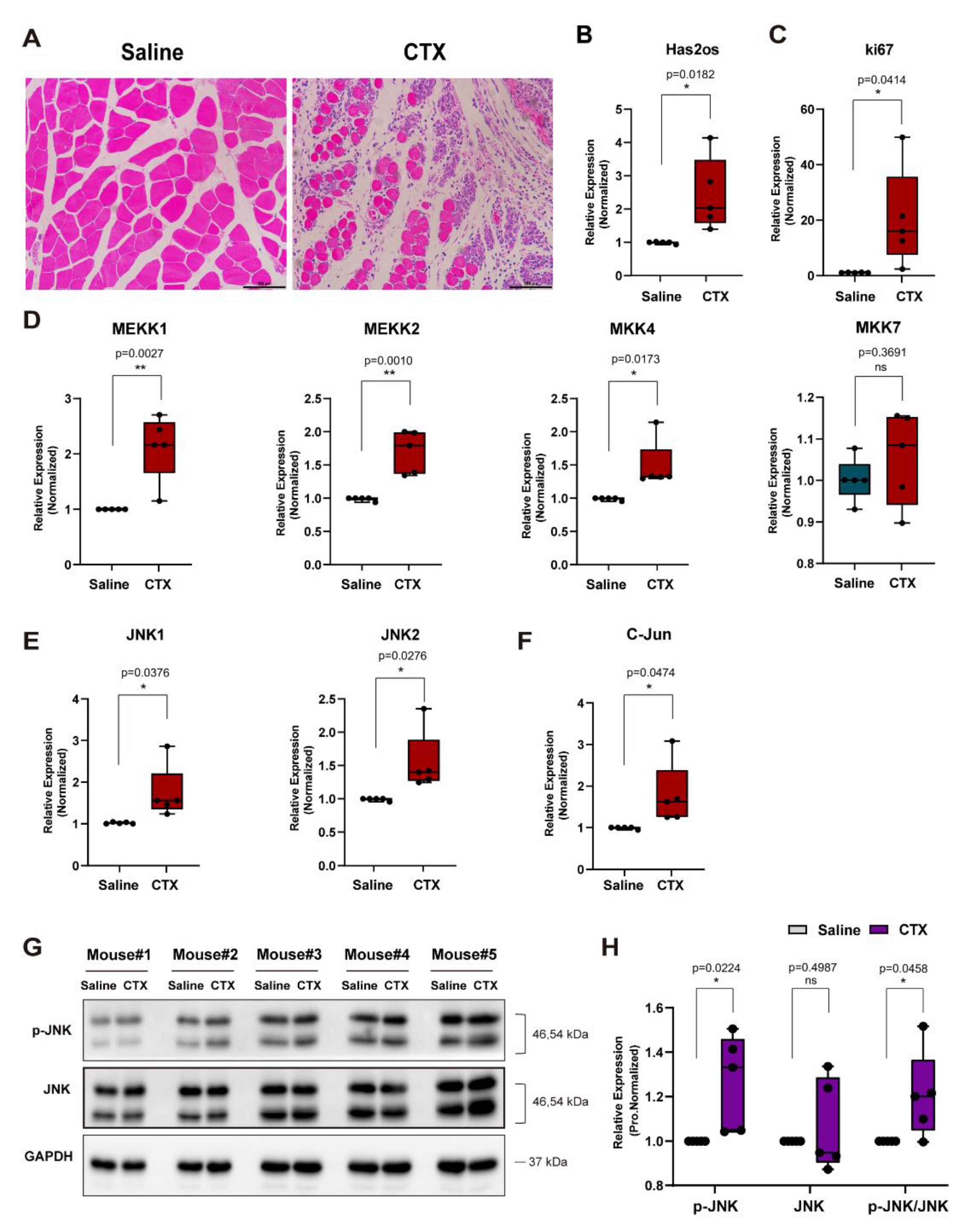
3.5. The lncRNA Has2os Is Involved in the Early Repair of the Muscle Injury
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Christ, B.; Brand-Saberi, B. Limb muscle development. Int. J. Dev. Biol. 2002, 46, 905–914. [Google Scholar] [PubMed]
- Cong, X.X.; Gao, X.K.; Rao, X.S.; Wen, J.; Liu, X.C.; Shi, Y.P.; He, M.Y.; Shen, W.L.; Shen, Y.; Ouyang, H.; et al. Rab5a activates IRS1 to coordinate IGF-AKT-mTOR signaling and myoblast differentiation during muscle regeneration. Cell Death Differ. 2020, 27, 2344–2362. [Google Scholar] [CrossRef] [PubMed]
- Bentzinger, C.F.; Wang, Y.X.; Rudnicki, M.A. Building Muscle: Molecular Regulation of Myogenesis. Cold Spring Harb. Perspect. Biol. 2012, 4, a008342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bryson-Richardson, R.J.; Currie, P. The genetics of vertebrate myogenesis. Nat. Rev. Genet. 2008, 9, 632–646. [Google Scholar] [CrossRef]
- Moncaut, N.; Rigby, P.W.J.; Carvajal, J.J. Dial M(RF) for myogenesis. FEBS J. 2013, 280, 3980–3990. [Google Scholar] [CrossRef] [Green Version]
- Le Grand, F.; Rudnicki, M.A. Skeletal muscle satellite cells and adult myogenesis. Curr. Opin. Cell Biol. 2007, 19, 628–633. [Google Scholar] [CrossRef] [Green Version]
- Xie, S.-J.; Li, J.-H.; Chen, H.-F.; Tan, Y.-Y.; Liu, S.-R.; Zhang, Y.; Xu, H.; Yang, J.-H.; Liu, S.; Zheng, L.-L.; et al. Inhibition of the JNK/MAPK signaling pathway by myogenesis-associated miRNAs is required for skeletal muscle development. Cell Death Differ. 2018, 25, 1581–1597. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Xie, S.; Chen, H.; Li, B.; Chen, Z.; Tan, Y.; Yang, J.; Zheng, L.; Xiao, Z.; Zhang, Q.; et al. The functional analysis of transiently upregulated miR-101 suggests a “braking” regulatory mechanism during myogenesis. Sci. China Life Sci. 2021, 64, 1612–1623. [Google Scholar] [CrossRef]
- Xie, S.J.; Lei, H.; Yang, B.; Diao, L.T.; Liao, J.Y.; He, J.H.; Xiao, Z.D. Dynamic m(6)A mRNA Methylation Reveals the Role of METTL3/14-m(6)A-MNK2-ERK Signaling Axis in Skeletal Muscle Differentiation and Regeneration. Front Cell Dev. Biol. 2021, 9, 744171. [Google Scholar] [CrossRef]
- Diao, L.-T.; Xie, S.-J.; Yu, P.-J.; Sun, Y.-J.; Yang, F.; Tan, Y.-Y.; Tao, S.; Hou, Y.-R.; Zheng, L.-L.; Xiao, Z.-D.; et al. N6-methyladenine demethylase ALKBH1 inhibits the differentiation of skeletal muscle. Exp. Cell Res. 2021, 400, 112492. [Google Scholar] [CrossRef]
- Chen, L.; Carmichael, G.G. Long noncoding RNAs in mammalian cells: What, where, and why? Wiley Interdiscip. Rev. RNA 2010, 1, 2–21. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.C.; Chang, H.Y. Molecular Mechanisms of Long Noncoding RNAs. Mol. Cell 2011, 43, 904–914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Jin, J.; Xu, Z.; Zuo, B. Functions and Regulatory Mechanisms of lncRNAs in Skeletal Myogenesis, Muscle Disease and Meat Production. Cells 2019, 8, 1107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Chen, X.; Sun, H.; Wang, H. Long non-coding RNAs in the regulation of skeletal myogenesis and muscle diseases. Cancer Lett. 2018, 417, 58–64. [Google Scholar] [CrossRef]
- Milligan, L.; Antoine, E.; Bisbal, C.; Weber, M.; Brunel, C.; Forné, T.; Cathala, G. H19 gene expression is up-regulated exclusively by stabilization of the RNA during muscle cell diferentiation. Oncogene 2000, 19, 5810–5816. [Google Scholar] [CrossRef] [Green Version]
- Cai, B.; Ma, M.; Zhang, J.; Kong, S.; Zhou, Z.; Li, Z.; Abdalla, B.A.; Xu, H.; Zhang, X.; Lawal, R.A.; et al. Long noncoding RNA ZFP36L2-AS functions as a metabolic modulator to regulate muscle development. Cell Death Dis. 2022, 13, 389. [Google Scholar] [CrossRef]
- Lv, W.; Jin, J.; Xu, Z.; Luo, H.; Guo, Y.; Wang, X.; Wang, S.; Zhang, J.; Zuo, H.; Bai, W.; et al. lncMGPF is a novel positive regulator of muscle growth and regeneration. J. Cachex-Sarcopenia Muscle 2020, 11, 1723–1746. [Google Scholar] [CrossRef]
- Martone, J.; Mariani, D.; Desideri, F.; Ballarino, M. Non-coding RNAs Shaping Muscle. Front. Cell Dev. Biol. 2020, 7, 394. [Google Scholar] [CrossRef]
- Cesana, M.; Cacchiarelli, D.; Legnini, I.; Santini, T.; Sthandier, O.; Chinappi, M.; Tramontano, A.; Bozzoni, I. A long noncoding RNA controls muscle differentiation by functioning as a competing endogenous RNA. Cell 2011, 147, 358–369. [Google Scholar] [CrossRef] [Green Version]
- Gong, C.; Li, Z.; Ramanujan, K.; Clay, I.; Zhang, Y.; Lemire-Brachat, S.; Glass, D.J. A Long Non-coding RNA, LncMyoD, Regulates Skeletal Muscle Differentiation by Blocking IMP2-Mediated mRNA Translation. Dev. Cell 2015, 34, 181–191. [Google Scholar] [CrossRef]
- Dong, A.; Preusch, C.B.; So, W.K.; Lin, K.; Luan, S.; Yi, R.; Cheung, T.H. A long noncoding RNA, LncMyoD, modulates chromatin accessibility to regulate muscle stem cell myo-genic lineage progression. Proc. Natl. Acad. Sci. USA 2020, 117, 32464–32475. [Google Scholar] [CrossRef] [PubMed]
- Hitachi, K.; Nakatani, M.; Takasaki, A.; Ouchi, Y.; Uezumi, A.; Ageta, H.; Inagaki, H.; Kurahashi, H.; Tsuchida, K. Myogenin promoter-associated lncRNA Myoparr is essential for myogenic differentiation. EMBO Rep. 2019, 20, e47468. [Google Scholar] [CrossRef] [PubMed]
- Sui, Y.; Han, Y.; Zhao, X.; Li, D.; Li, G. Long non-coding RNA Irm enhances myogenic differentiation by interacting with MEF2D. Cell Death Dis. 2019, 10, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, L.; Sun, K.; Chen, X.; Zhao, Y.; Wang, L.; Zhou, L.; Sun, H.; Wang, H. Genome-wide survey by ChIP-seq reveals YY1 regulation of lincRNAs in skeletal myogenesis. EMBO J. 2013, 32, 2575–2588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, L.; Sun, K.; Zhao, Y.; Zhang, S.; Wang, X.; Li, Y.; Lu, L.; Chen, X.; Chen, F.; Bao, X.; et al. Linc-YY1 promotes myogenic differentiation and muscle regeneration through an interaction with the transcription factor YY1. Nat. Commun. 2015, 6, 10026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Zhao, Y.; Bao, X.; Zhu, X.; Kwok, Y.K.-Y.; Sun, K.; Chen, X.; Huang, Y.; Jauch, R.; Esteban, M.A.; et al. LncRNA Dum interacts with Dnmts to regulate Dppa2 expression during myogenic differentiation and muscle regeneration. Cell Res. 2015, 25, 335–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, X.; Zhang, Y.; Li, T.; Ma, Z.; Jia, H.; Chen, Q.; Zhao, Y.; Zhai, L.; Zhong, R.; Li, C.; et al. Long non-coding RNA Linc-RAM enhances myogenic differentiation by interacting with MyoD. Nat. Commun. 2017, 8, 14016. [Google Scholar] [CrossRef] [Green Version]
- Xie, S.-J.; Tao, S.; Diao, L.-T.; Li, P.-L.; Chen, W.-C.; Zhou, Z.-G.; Hu, Y.-X.; Hou, Y.-R.; Lei, H.; Xu, W.-Y.; et al. Characterization of Long Non-coding RNAs Modified by m6A RNA Methylation in Skeletal Myogenesis. Front. Cell Dev. Biol. 2021, 9, 2766. [Google Scholar] [CrossRef]
- Matsumoto, K.; Li, Y.; Jakuba, C.; Sugiyama, Y.; Sayo, T.; Okuno, M.; Dealy, C.N.; Toole, B.P.; Takeda, J.; Yamaguchi, Y.; et al. Conditional inactivation of Has2 reveals a crucial role for hyaluronan in skeletal growth, pattern-ing, chondrocyte maturation and joint formation in the developing limb. Development 2009, 136, 2825–2835. [Google Scholar] [CrossRef] [Green Version]
- Hunt, L.C.; Gorman, C.; Kintakas, C.; McCulloch, D.R.; Mackie, E.J.; White, J.D. Hyaluronan synthesis and myogenesis: A requirement for hyaluronan synthesis during myogenic dif-ferentiation independent of pericellular matrix formation. J. Biol. Chem. 2013, 288, 13006–13021. [Google Scholar] [CrossRef]
- Gesteira, T.F.; Sun, M.; Coulson-Thomas, Y.M.; Yamaguchi, Y.; Yeh, L.K.; Hascall, V.; Coulson-Thomas, V.J. Hyaluronan Rich Microenvironment in the Limbal Stem Cell Niche Regulates Limbal Stem Cell Dif-ferentiation. Investig. Ophthalmol. Vis. Sci. 2017, 58, 4407–4421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simpson, R.M.; Hong, X.; Wong, M.M.; Karamariti, E.; Bhaloo, S.I.; Warren, D.; Kong, W.; Hu, Y.; Xu, Q. Hyaluronan Is Crucial for Stem Cell Differentiation into Smooth Muscle Lineage. Stem Cells 2016, 34, 1225–1238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.T. Epigenetic Regulation by Long Noncoding RNAs. Science 2012, 338, 1435–1439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burattini, S.; Ferri, P.; Battistelli, M.; Curci, R.; Luchetti, F.; Falcieri, E. C2C12 murine myoblasts as a model of skeletal muscle development: Morpho-functional characterization. Eur. J. Histochem. 2004, 48, 223–234. [Google Scholar] [PubMed]
- Shang, M.; Cappellesso, F.; Amorim, R.; Serneels, J.; Virga, F.; Eelen, G.; Carobbio, S.; Rincon, M.Y.; Maechler, P.; De Bock, K.; et al. Macrophage-derived glutamine boosts satellite cells and muscle regeneration. Nature 2020, 587, 626–631. [Google Scholar] [CrossRef]
- Terazawa, S.; Nakano, M.; Yamamoto, A.; Imokawa, G. Mycosporine-like amino acids stimulate hyaluronan secretion by up-regulating hyaluronan synthase 2 via activation of the p38/MSK1/CREB/c-Fos/AP-1 axis. J. Biol. Chem. 2020, 295, 7274–7288. [Google Scholar] [CrossRef] [Green Version]
- Buonaiuto, G.; Desideri, F.; Taliani, V.; Ballarino, M. Muscle Regeneration and RNA: New Perspectives for Ancient Molecules. Cells 2021, 10, 2512. [Google Scholar] [CrossRef]
- Huang, H.; Xing, D.; Zhang, Q.; Li, H.; Lin, J.; He, Z.; Lin, J. LncRNAs as a new regulator of chronic musculoskeletal disorder. Cell Prolif. 2021, 54, e13113. [Google Scholar] [CrossRef]
- Li, Z.; Cai, B.; Abdalla, B.A.; Zhu, X.; Zheng, M.; Han, P.; Nie, Q. LncIRS1 controls muscle atrophy via sponging miR-15 family to activate IGF1-PI3K/AKT pathway. J. Cachexia Sarcopenia Muscle 2019, 10, 391–410. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Primer | Sequence (5′-->3′) |
|---|---|---|
| MyHC | MyHC-RT-qF | CGCAAGAATGTTCTCAGGCT |
| MyHC-RT-qR | GCCAGGTTGACATTGGATTG | |
| Mef2C | Mef2C-RT-qF | ATCCCGATGCAGACGATTCAG |
| Mef2C-RT-qR | AACAGCACACAATCTTTGCCT | |
| MyoD | MyoD-RT-qF | CTACCCAAGGTGGAGATCCTG |
| MyoD-RT-qR | CACTGTAGTAGGCGGTGTCGT | |
| MyoG | MyoG-RT-qF | ATCCAGTACATTGAGCGCCTAC |
| MyoG-RT-qR | GACGTAAGGGAGTGCAGATTGT | |
| Has2os | Has2os-RT-qF | CTCCGGAGCCCCTTTGAC |
| Has2os-RT-qR | ACGCGTTTGGGTGAACTTTG | |
| Has2 | Has2-RT-qF | CATCTGTGGAGATGGTGAAGGTC |
| Has2-RT-qR | AGCCATCCAGTATCTCACGCTG | |
| ki67 | ki67-RT-qF | GAGGAGAAACGCCAACCAAGAG |
| ki67-RT-qR | TTTGTCCTCGGTGGCGTTATCC | |
| Pax7 | Pax7-RT-qF | GTTCGGGAAGAAAGAGGACGAC |
| Pax7-RT-qR | GGTTCTGATTCCACATCTGAGCC | |
| JNK1 | JNK1-RT-qF | CGTTCTCCGCTACGGGCTT |
| JNK1-RT-qR | AGTCTAATGGCTTCATCCAAATA | |
| JNK2 | JNK2-RT-qF | CTGGAGCCCAAGGAATTGT |
| JNK2-RT-qR | GCGTTTGGTTCTGAAAAGGA | |
| MEKK1 | MEKK1-RT-qF | TCCAGAAGTTTGTGTCACGC |
| MEKK1-RT-qR | GGGACACATCTGCTCCTCTT | |
| MEKK2 | MEKK2-RT-qF | CCCAGAGTATGACGACAGTCGA |
| MEKK2-RT-qR | GGTAGACCCTACCAAAAGCTCC | |
| MKK4 | MKK4-RT-qF | TAAACCGTGCTGTCGATTTG |
| MKK4-RT-qR | TCGGACCTCCAGCTTCTG | |
| MKK7 | MKK7-RT-qF | GAAGCAGGAGAACCGTGAGG |
| MKK7-RT-qR | AGGAGCAGGGCTTAGAGTG | |
| C-Jun | C-Jun-RT-qF | GAAAAGTAGCCCCCAACCTC |
| C-Jun-RT-qR | GGGACACAGCTTTCACCCTA | |
| GAPDH | GAPDH-RT-qF | CGTCCCGTAGACAAAATGGT |
| GAPDH-RT-qR | TCAATGAAGGGGTCGTTGAT |
| Gene Name | Sense (5′-->3′) | Antisense (5′-->3′) |
|---|---|---|
| si-NC | UUCUCCGAACGUGUCACGUTT | ACGUGACACGUUCGGAGAATT |
| si-Has2os-#1 | GGAGUAUUGAUGCAGGCAATT | UUGCCUGCAUCAAUACUCCTT |
| si-Has2os-#2 | GGGACAAUCAGCUUUCCUUTT | AAGGAAAGCUGAUUGUCCCTT |
| si-Has2os-#3 | GCGUCUGUAUAACGUACCUTT | AGGUACGUUAUACAGACGCTT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, W.; Chen, W.; Liu, P.; Qian, S.; Tao, S.; Huang, M.; Xu, W.; Li, C.; Chen, X.; Lin, H.; et al. Role of lncRNA Has2os in Skeletal Muscle Differentiation and Regeneration. Cells 2022, 11, 3497. https://doi.org/10.3390/cells11213497
Chen W, Chen W, Liu P, Qian S, Tao S, Huang M, Xu W, Li C, Chen X, Lin H, et al. Role of lncRNA Has2os in Skeletal Muscle Differentiation and Regeneration. Cells. 2022; 11(21):3497. https://doi.org/10.3390/cells11213497
Chicago/Turabian StyleChen, Wanxin, Weicai Chen, Peng Liu, Shiyu Qian, Shuang Tao, Mengchun Huang, Wanyi Xu, Cuiping Li, Xiaoyan Chen, Huizhu Lin, and et al. 2022. "Role of lncRNA Has2os in Skeletal Muscle Differentiation and Regeneration" Cells 11, no. 21: 3497. https://doi.org/10.3390/cells11213497
APA StyleChen, W., Chen, W., Liu, P., Qian, S., Tao, S., Huang, M., Xu, W., Li, C., Chen, X., Lin, H., Qin, Z., Lu, J., & Xie, S. (2022). Role of lncRNA Has2os in Skeletal Muscle Differentiation and Regeneration. Cells, 11(21), 3497. https://doi.org/10.3390/cells11213497