
Propranolol Modulates Cerebellar Circuit Activity and Reduces Tremor

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Maintenance
2.2. Genetically Engineered Mouse Lines
2.3. Propranolol Administration
2.4. Tremor Recording and Analysis
2.5. Non-Tremor Movement Events Detection
2.6. Open Field Assay
2.7. Footprinting Assay
2.8. Surgery for Awake Head-Fixed Neural Recordings
2.9. In Vivo Electrophysiology
2.10. Immunohistochemistry
2.11. Imaging of Immunostained Tissue Sections
2.12. Data Analyses
2.13. Data Visualization
3. Results
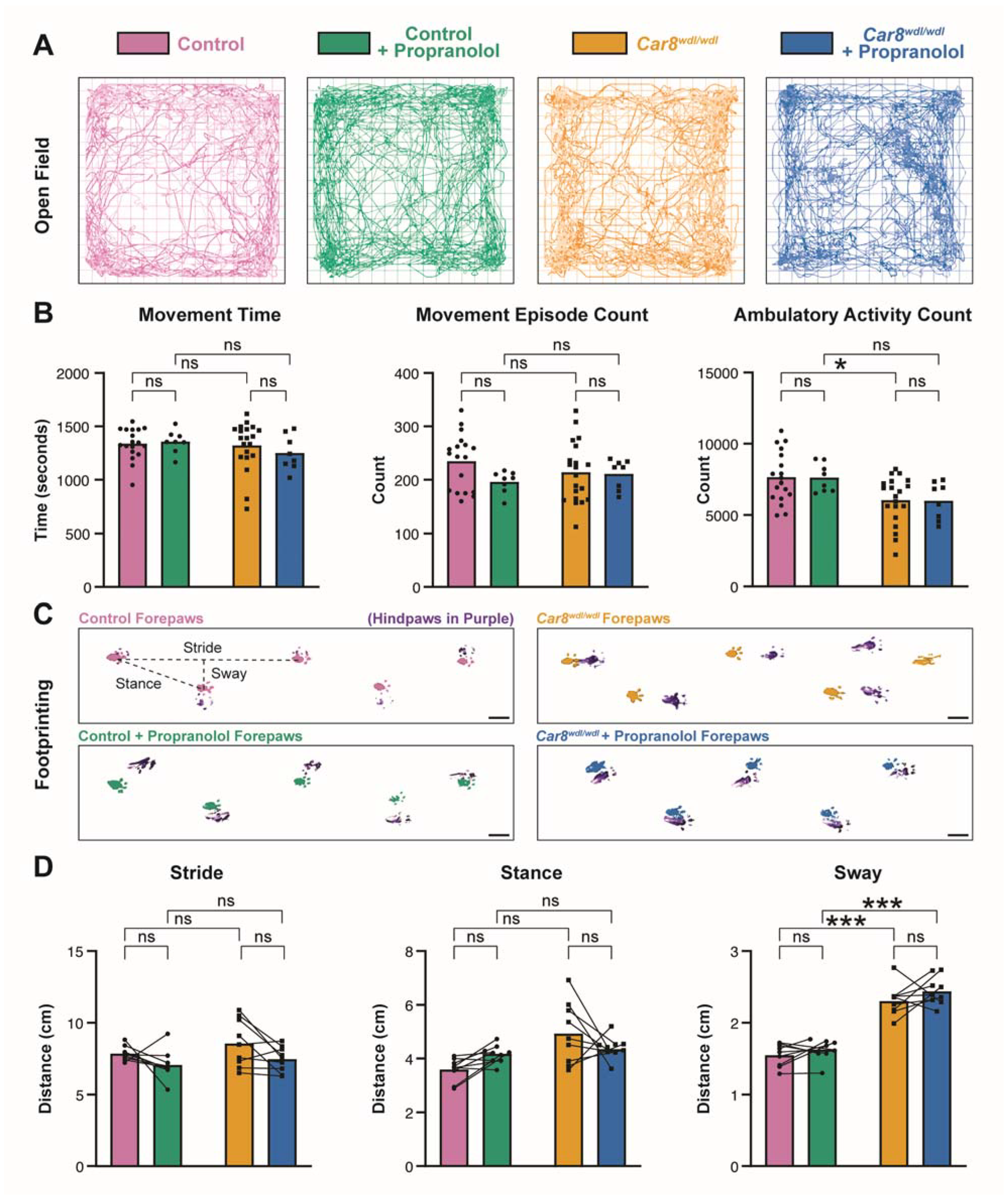
3.1. Propranolol Reduces Tremor in Control and Car8wdl/wdl Mice
3.2. Propranolol Does Not Impact General Locomotor Behaviors
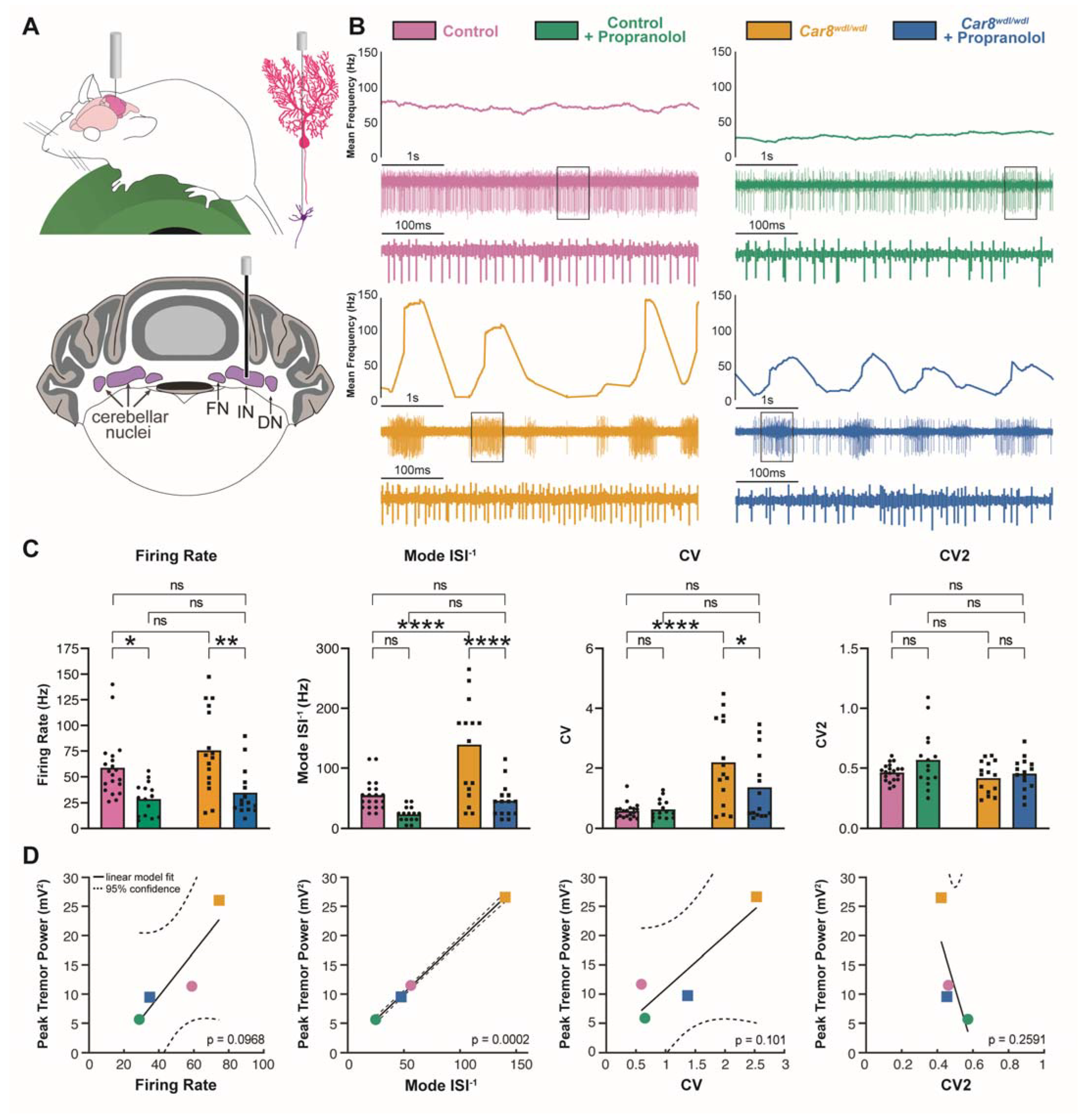
3.3. Propranolol Modulates Purkinje Cell and Cerebellar Nuclei Neuron Firing Activity
3.4. β1 and β2 Adrenergic Receptors Are Expressed throughout the Cerebellar Cortex
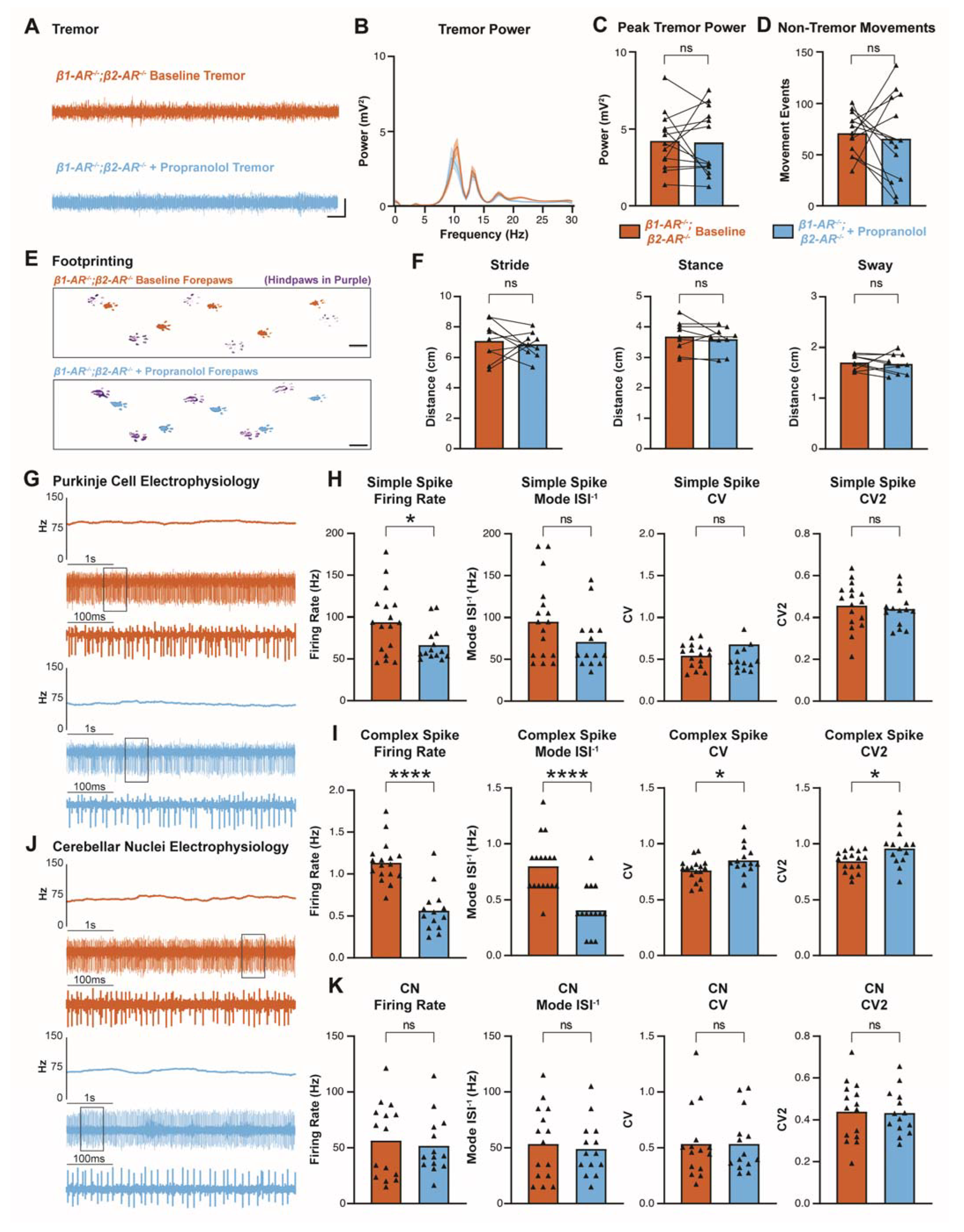
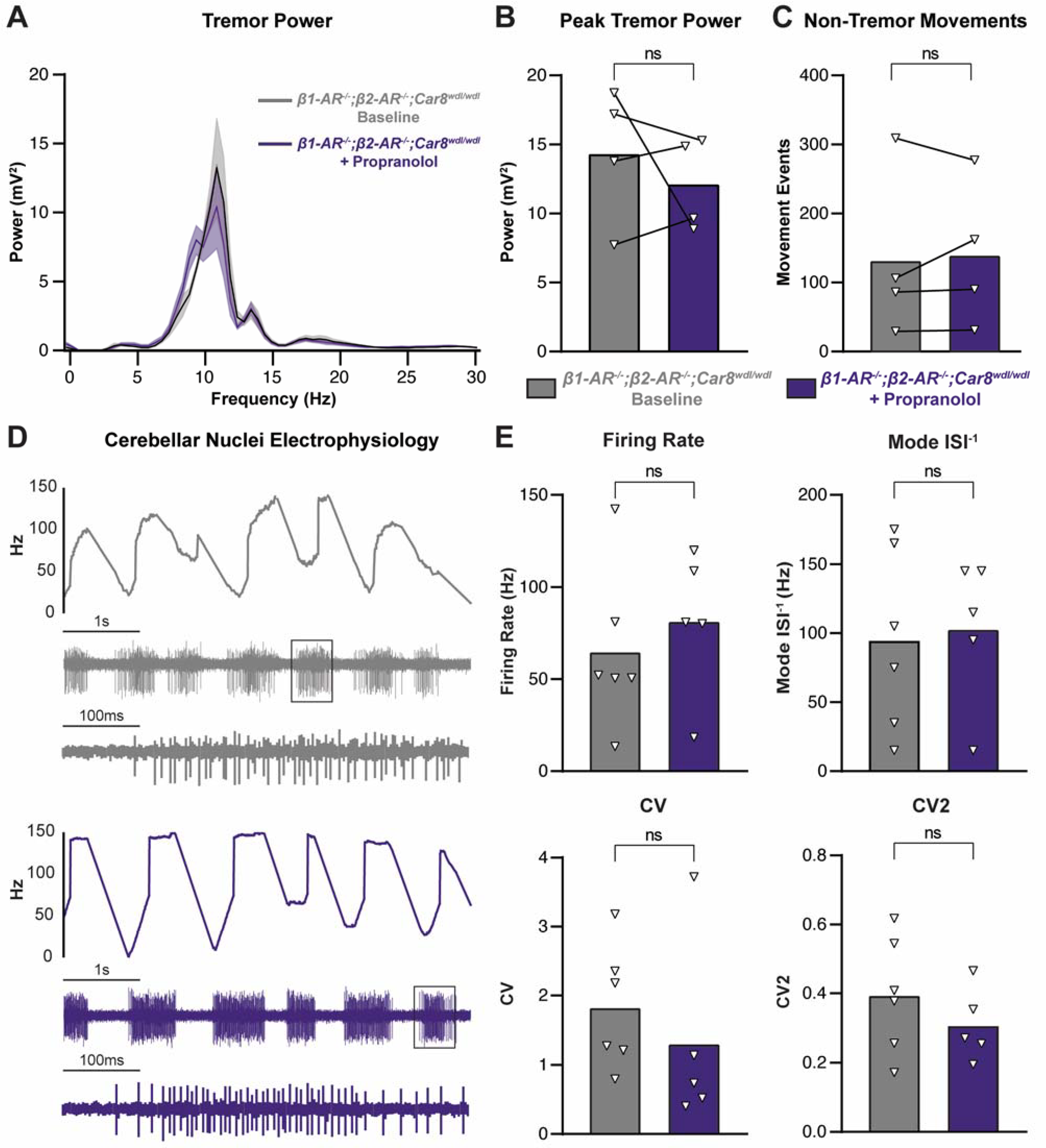
3.5. Propranolol Acts through β1 and β2 Adrenergic Receptors to Reduce Tremor
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Louis, E.D.; McCreary, M. How Common is Essential Tremor? Update on the Worldwide Prevalence of Essential Tremor. Tremor Other Hyperkinetic Mov. 2021, 11, 28. [Google Scholar] [CrossRef]
- Louis, E.D.; Machado, D.G. Tremor-related quality of life: A comparison of essential tremor vs. Parkinson’s disease patients. Park. Relat. Disord. 2015, 21, 729–735. [Google Scholar] [CrossRef] [Green Version]
- Jiménez-Jiménez, F.; Alonso-Navarro, H.; García-Martín, E.; Álvarez, I.; Pastor, P.; Agúndez, J. Genomic Markers for Essential Tremor. Pharmaceuticals 2021, 14, 516. [Google Scholar] [CrossRef] [PubMed]
- Graham, J.D.P. STATIC TREMOR IN ANXIETY STATES. J. Neurol. Neurosurg. Psychiatry 1945, 8, 57–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tyrer, P.J.; Lader, M.H. Tremor in Acute and Chronic Anxiety. Arch. Gen. Psychiatry 1974, 31, 506–509. [Google Scholar] [CrossRef]
- Koller, W.; O’Hara, R.; Dorus, W.; Bauer, J. Tremor in chronic alcoholism. Neurology 1985, 35, 1660. [Google Scholar] [CrossRef] [PubMed]
- Koch, M.; Mostert, J.; Heersema, D.; De Keyser, J. Tremor in multiple sclerosis. J. Neurol. 2007, 254, 133–145. [Google Scholar] [CrossRef] [Green Version]
- Berlin, I.; Grimaldi, A.; Payan, C.; Sachon, C.; Bosquet, F.; Thervet, F.; Puech, A.J. Hypoglycemic Symptoms and Decreased β-Adrenergic Sensitivity in Insulin-Dependent Diabetic Patients. Diabetes Care 1987, 10, 742–747. [Google Scholar] [CrossRef]
- Haddad, P.M.; Dursun, S.M. Neurological complications of psychiatric drugs: Clinical features and management. Hum. Psychopharmacol. Clin. Exp. 2007, 23, S15–S26. [Google Scholar] [CrossRef]
- Brennan, K.C.; Jurewicz, E.C.; Ford, B.; Pullman, S.L.; Louis, E.D. Is essential tremor predominantly a kinetic or a postural tremor? A clinical and electrophysiological study. Mov. Disord. 2002, 17, 313–316. [Google Scholar] [CrossRef] [PubMed]
- Thanvi, B.; Lo, N.; Robinson, T. Essential tremor—the most common movement disorder in older people. Age Ageing 2006, 35, 344–349. [Google Scholar] [CrossRef] [Green Version]
- Elble, R.J. Chapter 22 Physiologic and enhanced physiologic tremor. Handb. Clin. Neurophysiol. 2003, 1, 357–364. [Google Scholar] [CrossRef]
- Growdon, J.H.; Shahani, B.T.; Young, R.R. The effect of alcohol on essential tremor. Neurology 1975, 25, 259. [Google Scholar] [CrossRef]
- Defazio, G.; Conte, A.; Gigante, A.F.; Fabbrini, G.; Berardelli, A. Is tremor in dystonia a phenotypic feature of dystonia? Neurology 2015, 84, 1053–1059. [Google Scholar] [CrossRef] [PubMed]
- Pandey, S.; Sarma, N. Tremor in dystonia. Park. Relat. Disord. 2016, 29, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Hagerman, P.J.; Hagerman, R.J. Fragile X-associated tremor/ataxia syndrome. Ann. N. Y. Acad. Sci. 2015, 1338, 58–70. [Google Scholar] [CrossRef] [Green Version]
- Axelrad, J.; Louis, E.D.; Honig, L.S.; Flores, I.; Ross, G.W.; Pahwa, R.; Lyons, K.E.; Faust, P.L.; Vonsattel, J.P.G. Reduced Purkinje Cell Number in Essential Tremor. Arch. Neurol. 2008, 65, 101–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erickson-Davis, C.R.; Faust, P.L.; Vonsattel, J.-P.G.; Gupta, S.; Honig, L.S.; Louis, E.D. "Hairy Baskets” Associated With Degenerative Purkinje Cell Changes in Essential Tremor. J. Neuropathol. Exp. Neurol. 2010, 69, 262–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Louis, E.D.; Frucht, S.; Rios, E. Intention tremor in essential tremor: Prevalence and association with disease duration. Mov. Disord. 2009, 24, 626–627. [Google Scholar] [CrossRef] [Green Version]
- Louis, E.D.; Vonsattel, J.P.G.; Honig, L.S.; Lawton, A.; Moskowitz, C.; Ford, B.; Frucht, S. Essential Tremor Associated With Pathologic Changes in the Cerebellum. Arch. Neurol. 2006, 63, 1189–1193. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Ma, K.; Faust, P.L.; Honig, L.S.; Cortés, E.; Vonsattel, J.-P.G.; Louis, E.D. Increased number of Purkinje cell dendritic swellings in essential tremor. Eur. J. Neurol. 2011, 19, 625–630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benabid, A.; Pollak, P.; Louveau, A.; Henry, S.; De Rougemont, J. Combined (Thalamotomy and Stimulation) Stereotactic Surgery of the VIM Thalamic Nucleus for Bilateral Parkinson Disease. Ster. Funct. Neurosurg. 1987, 50, 344–346. [Google Scholar] [CrossRef]
- Cury, R.G.; Fraix, V.; Castrioto, A.; Fernández, M.A.P.; Krack, P.; Chabardes, S.; Seigneuret, E.; Alho, E.J.L.; Benabid, A.-L.; Moro, E. Thalamic deep brain stimulation for tremor in Parkinson disease, essential tremor, and dystonia. Neurology 2017, 89, 1416–1423. [Google Scholar] [CrossRef]
- Nazzaro, J.M.; Lyons, K.E.; Pahwa, R. Deep Brain Stimulation for Essential Tremor. Handb. Clin. Neurol. 2013, 116, 155–166. [Google Scholar]
- Ondo, W.; Jankovic, J.; Schwartz, K.; Almaguer, M.; Simpson, R.K. Unilateral thalamic deep brain stimulation for refractory essential tremor and Parkinson’s disease tremor. Neurology 1998, 51, 1063–1069. [Google Scholar] [CrossRef]
- Schuurman, P.R.; Bosch, D.A.; Bossuyt, P.M.M.; Bonsel, G.J.; van Someren, E.J.W.; de Bie, R.M.A.; Merkus, M.P.; Speelman, J.D. A Comparison of Continuous Thalamic Stimulation and Thalamotomy for Suppression of Severe Tremor. New Engl. J. Med. 2000, 342, 461–468. [Google Scholar] [CrossRef] [Green Version]
- Elias, W.J.; Huss, D.; Voss, T.; Loomba, J.; Khaled, M.; Zadicario, E.; Frysinger, R.C.; Sperling, S.A.; Wylie, S.; Monteith, S.J.; et al. A Pilot Study of Focused Ultrasound Thalamotomy for Essential Tremor. New Engl. J. Med. 2013, 369, 640–648. [Google Scholar] [CrossRef] [PubMed]
- Lipsman, N.; Schwartz, M.L.; Huang, Y.; Lee, L.; Sankar, T.; Chapman, M.; Hynynen, K.; Lozano, A.M. MR-Guided Focused Ultrasound Thalamotomy for Essential Tremor: A Proof-of-Concept Study. Lancet Neurol. 2013, 12, 462–468. [Google Scholar] [CrossRef] [PubMed]
- Rohani, M.; Fasano, A. Focused Ultrasound for Essential Tremor: Review of the Evidence and Discussion of Current Hurdles. Tremor Other Hyperkinetic Mov. 2017, 7, 462. [Google Scholar] [CrossRef] [PubMed]
- Zesiewicz, T.A.; Elble, R.J.; Louis, E.D.; Gronseth, G.S.; Ondo, W.G.; Dewey, R.B.; Okun, M.; Sullivan, K.L.; Weiner, W.J. Evidence-based guideline update: Treatment of essential tremor: Report of the Quality Standards Subcommittee of the American Academy of Neurology. Neurology 2011, 77, 1752–1755. [Google Scholar] [CrossRef] [Green Version]
- Deuschl, G.; Raethjen, J.; Hellriegel, H.; Elble, R. Treatment of patients with essential tremor. Lancet Neurol. 2011, 10, 148–161. [Google Scholar] [CrossRef]
- Haubenberger, D.; Hallett, M. Essential Tremor. New Engl. J. Med. 2018, 378, 1802–1810. [Google Scholar] [CrossRef]
- Shanker, V. Essential tremor: Diagnosis and management. Bmj 2019, 366, l4485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Black, J.; Stephenson, J. Pharmacology of a new adrenergic beta-receptor-blocking compound (nethalide). Lancet 1962, 280, 311–314. [Google Scholar] [CrossRef]
- Black, J.; Crowther, A.; Shanks, R.; Smith, L.; Dornhorst, A. A new adrenergic: Beta-receptor antagonist. Lancet 1964, 283, 1080–1081. [Google Scholar] [CrossRef]
- Owen, D.; Marsden, C. Effect of adrenergic β-blockade on parkinsonian tremor. Lancet 1965, 286, 1259–1262. [Google Scholar] [CrossRef] [PubMed]
- Barrett, A.M. The pharmacology of "inderal" (propranolol hydrochloride). Brux Med 1970, 50. [Google Scholar]
- Shand, D.G. Pharmacokinetics of propranolol: A review. Postgrad. Med J. 1976, 52. [Google Scholar]
- Nies, A.S.; Shand, D.G. Clinical pharmacology of propranolol. Circulation 1975, 52, 6–15. [Google Scholar] [CrossRef] [Green Version]
- Drayer, D.E. Lipophilicity, Hydrophilicity, and the Central Nervous System Side Effects of Beta Blockers. Pharmacother. J. Hum. Pharmacol. Drug Ther. 1987, 7, 87–91. [Google Scholar] [CrossRef]
- Hedera, P.; Cibulčík, F.; Davis, T.L. Pharmacotherapy of Essential Tremor. J. Central Nerv. Syst. Dis. 2013, 5, 43–55. [Google Scholar] [CrossRef] [PubMed]
- Lader, M.H.; Tyrer, P.J. Central and peripheral effects of propranolol and sotalol in normal human subjects. Br. J. Pharmacol. 1972, 45, 557–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morgan, M.H.; Hewer, R.L.; Cooper, R. Effect of the beta adrenergic blocking agent propranolol on essential tremor. J. Neurol. Neurosurg. Psychiatry 1973, 36, 618–624. [Google Scholar] [CrossRef]
- Sweet, R.D.; Blumberg, J.; Lee, J.E.; McDowell, F.H. Propranolol treatment of essential tremor. Neurology 1974, 24, 64. [Google Scholar] [CrossRef] [PubMed]
- Tolosa, E.S.; Loewenson, R.B. Essential tremor: Treatment with propranolol. Neurology 1975, 25, 1041. [Google Scholar] [CrossRef] [PubMed]
- Calzetti, S.; Findley, L.J.; A Gresty, M.; Perucca, E.; Richens, A. Metoprolol and propranolol in essential tremor: A double-blind, controlled study. J. Neurol. Neurosurg. Psychiatry 1981, 44, 814–819. [Google Scholar] [CrossRef] [Green Version]
- Abila, B.; Wilson, J.; Marshall, R.; Richens, A. The tremorolytic action of beta-adrenoceptor blockers in essential, physiological and isoprenaline-induced tremor is mediated by beta- adrenoceptors located in a deep peripheral compartment. Br. J. Clin. Pharmacol. 1985, 20, 369–376. [Google Scholar] [CrossRef] [Green Version]
- Leszkovszky, G.; Tardos, L. Some effects of propranolol on the central nervous system. J. Pharm. Pharmacol. 1965, 17, 518–520. [Google Scholar] [CrossRef]
- Murmann, W.; Almirante, L.; Saccani-Guelfi, M. Effects of hexobarbitone, ether, morphine, and urethane upon the acute toxicity of propranolol and D-(—)-INPEA. J. Pharm. Pharmacol. 1966, 18, 692–694. [Google Scholar] [CrossRef]
- Hinshelwood, R.D. Hallucinations and propranolol. BMJ 1969, 2, 445. [Google Scholar] [CrossRef] [Green Version]
- Bainbridge, J.; Greenwood, D. Tranquillizing effects of propranolol demonstrated in rats. Neuropharmacology 1971, 10, 453–458. [Google Scholar] [CrossRef]
- Pan, M.-K.; Kuo, S.-H. Tracking the central and peripheral origin of tremor. Clin. Neurophysiol. 2018, 129, 1451–1452. [Google Scholar] [CrossRef] [PubMed]
- Nicoletti, V.; Cecchi, P.; Pesaresi, I.; Frosini, D.; Cosottini, M.; Ceravolo, R. Cerebello-thalamo-cortical network is intrinsically altered in essential tremor: Evidence from a resting state functional MRI study. Sci. Rep. 2020, 10, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Pinto, A.D.; Lang, A.E.; Chen, R. The cerebellothalamocortical pathway in essential tremor. Neurology 2003, 60, 1985–1987. [Google Scholar] [CrossRef] [PubMed]
- Ni, Z.; Pinto, A.D.; Lang, A.E.; Chen, R. Involvement of the Cerebellothalamocortical Pathway in Parkinson Disease. Ann. Neurol. 2010, 68, 816–824. [Google Scholar] [CrossRef] [PubMed]
- Pedrosa, D.J.; Quatuor, E.-L.; Reck, C.; Pauls, K.A.M.; Huber, C.A.; Visser-Vandewalle, V.; Timmermann, L. Thalamomuscular Coherence in Essential Tremor: Hen or Egg in the Emergence of Tremor? J. Neurosci. 2014, 34, 14475–14483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paris-Robidas, S.; Brochu, E.; Sintes, M.; Emond, V.; Bousquet, M.; Vandal, M.; Pilote, M.; Tremblay, C.; Di Paolo, T.; Rajput, A.H.; et al. Defective dentate nucleus GABA receptors in essential tremor. Brain 2011, 135, 105–116. [Google Scholar] [CrossRef] [Green Version]
- Louis, E.D.; Rabinowitz, D.; Choe, M.; Tate, W.J.; Kelly, G.C.; Kuo, S.-H.; Faust, P.L. Mapping Purkinje Cell Placement Along the Purkinje Cell Layer: An Analysis of Postmortem Tissue from Essential Tremor Patients vs. Controls. Cerebellum 2015, 15, 726–731. [Google Scholar] [CrossRef] [Green Version]
- Pan, M.-K.; Li, Y.-S.; Wong, S.-B.; Ni, C.-L.; Wang, Y.-M.; Liu, W.-C.; Lu, L.-Y.; Lee, J.-C.; Cortes, E.P.; Vonsattel, J.-P.G.; et al. Cerebellar oscillations driven by synaptic pruning deficits of cerebellar climbing fibers contribute to tremor pathophysiology. Sci. Transl. Med. 2020, 12, eaay1769. [Google Scholar] [CrossRef]
- Brown, A.M.; White, J.J.; E van der Heijden, M.; Zhou, J.; Lin, T.; Sillitoe, R.V. Purkinje cell misfiring generates high-amplitude action tremors that are corrected by cerebellar deep brain stimulation. eLife 2020, 9, e51928. [Google Scholar] [CrossRef]
- Paraguay, I.B.; França, C.; Duarte, K.P.; Diniz, J.M.; Galhardoni, R.; Silva, V.; Iglesio, R.; Bissoli, A.B.; Menezes, J.R.; Carra, R.B.; et al. Dentate nucleus stimulation for essential tremor. Park. Relat. Disord. 2020, 82, 121–122. [Google Scholar] [CrossRef] [PubMed]
- Cury, R.G.; França, C.; Duarte, K.P.; Paraguay, I.; Diniz, J.M.; Cunha, P.; Galhardoni, R.; Silva, V.; Iglesio, R.; Bissoli, A.B.; et al. Safety and Outcomes of Dentate Nucleus Deep Brain Stimulation for Cerebellar Ataxia. Cerebellum 2021, 21, 861–865. [Google Scholar] [CrossRef] [PubMed]
- White, J.J.; Arancillo, M.; King, A.; Lin, T.; Miterko, L.N.; Gebre, S.A.; Sillitoe, R.V. Pathogenesis of severe ataxia and tremor without the typical signs of neurodegeneration. Neurobiol. Dis. 2016, 86, 86–98. [Google Scholar] [CrossRef]
- Kuo, S.-H.; Louis, E.D.; Faust, P.L.; Handforth, A.; Chang, S.-Y.; Avlar, B.; Lang, E.J.; Pan, M.-K.; Miterko, L.N.; Brown, A.M.; et al. Current Opinions and Consensus for Studying Tremor in Animal Models. Cerebellum 2019, 18, 1036–1063. [Google Scholar] [CrossRef] [PubMed]
- Jiao, Y.; Yan, J.; Zhao, Y.; Donahue, L.R.; Beamer, W.G.; Li, X.; Roe, B.A.; LeDoux, M.S.; Gu, W. Carbonic Anhydrase-Related Protein VIII Deficiency Is Associated With a Distinctive Lifelong Gait Disorder in Waddles Mice. Genetics 2005, 171, 1239–1246. [Google Scholar] [CrossRef] [Green Version]
- Miterko, L.N.; White, J.J.; Lin, T.; Brown, A.M.; O’Donovan, K.J.; Sillitoe, R.V. Persistent motor dysfunction despite homeostatic rescue of cerebellar morphogenesis in the Car8 waddles mutant mouse. Neural Dev. 2019, 14, 6. [Google Scholar] [CrossRef] [Green Version]
- Rohrer, D.K.; Chruscinski, A.; Schauble, E.H.; Bernstein, D.; Kobilka, B.K. Cardiovascular and Metabolic Alterations in Mice Lacking Both β1- and β2-Adrenergic Receptors. J. Biol. Chem. 1999, 274, 16701–16708. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Brown, A.M.; Lackey, E.P.; Arancillo, M.; Lin, T.; Sillitoe, R.V. Purkinje cell neurotransmission patterns cerebellar basket cells into zonal modules defined by distinct pinceau sizes. eLife 2020, 9, e55569. [Google Scholar] [CrossRef]
- Miterko, L.N.; Lin, T.; Zhou, J.; van der Heijden, M.E.; Beckinghausen, J.; White, J.J.; Sillitoe, R.V. Neuromodulation of the cerebellum rescues movement in a mouse model of ataxia. Nat. Commun. 2021, 12, 1295. [Google Scholar] [CrossRef]
- Carter, R.J.; Lione, L.A.; Humby, T.; Mangiarini, L.; Mahal, A.; Bates, G.; Dunnett, S.; Morton, A.J. Characterization of Progressive Motor Deficits in Mice Transgenic for the Human Huntington’s Disease Mutation. J. Neurosci. 1999, 19, 3248–3257. [Google Scholar] [CrossRef] [Green Version]
- White, J.J.; Lin, T.; Brown, A.M.; Arancillo, M.; Lackey, E.P.; Stay, T.L.; Sillitoe, R.V. An optimized surgical approach for obtaining stable extracellular single-unit recordings from the cerebellum of head-fixed behaving mice. J. Neurosci. Methods 2016, 262, 21–31. [Google Scholar] [CrossRef] [Green Version]
- White, J.J.; Sillitoe, R.V. Genetic silencing of olivocerebellar synapses causes dystonia-like behaviour in mice. Nat. Commun. 2017, 8, 14912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arancillo, M.; White, J.J.; Lin, T.; Stay, T.L.; Sillitoe, R.V. In vivo analysis of Purkinje cell firing properties during postnatal mouse development. J. Neurophysiol. 2015, 113, 578–591. [Google Scholar] [CrossRef]
- E van der Heijden, M.; Lackey, E.P.; Perez, R.; Ișleyen, F.S.; Brown, A.M.; Donofrio, S.G.; Lin, T.; Zoghbi, H.Y.; Sillitoe, R.V. Maturation of Purkinje cell firing properties relies on neurogenesis of excitatory neurons. eLife 2021, 10, e68045. [Google Scholar] [CrossRef]
- White, J.J.; Arancillo, M.; Stay, T.; George-Jones, N.; Levy, S.; Heck, D.; Sillitoe, R.V. Cerebellar Zonal Patterning Relies on Purkinje Cell Neurotransmission. J. Neurosci. 2014, 34, 8231–8245. [Google Scholar] [CrossRef] [Green Version]
- van der Heijden, M.E.; Brown, A.M.; Sillitoe, R.V. Influence of data sampling methods on the representation of neural spiking activity in vivo. iScience 2022, 25. [Google Scholar] [CrossRef]
- Heijden, M.E.; Kizek, D.J.; Perez, R.; Ruff, E.K.; Ehrlich, M.E.; Sillitoe, R.V. Abnormal cerebellar function and tremor in a mouse model for non-manifesting partially penetrant dystonia type 6. J. Physiol. 2021, 599, 2037–2054. [Google Scholar] [CrossRef] [PubMed]
- Holt, G.R.; Softky, W.R.; Koch, C.; Douglas, R.J. Comparison of discharge variability in vitro and in vivo in cat visual cortex neurons. J. Neurophysiol. 1996, 75, 1806–1814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sillitoe, R.V.; Vogel, M.; Joyner, A.L. Engrailed Homeobox Genes Regulate Establishment of the Cerebellar Afferent Circuit Map. J. Neurosci. 2010, 30, 10015–10024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sillitoe, R.V.; Kunzle, H.; Hawkes, R. Zebrin II compartmentation of the cerebellum in a basal insectivore, the Madagascan hedgehog tenrec Echinops telfairi. J. Anat. 2003, 203, 283–296. [Google Scholar] [CrossRef]
- White, J.J.; Sillitoe, R.V. Postnatal development of cerebellar zones revealed by neurofilament heavy chain protein expression. Front. Neuroanat. 2013, 7, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnold, D.B.; Heintz, N. A calcium responsive element that regulates expression of two calcium binding proteins in Purkinje cells. Proc. Natl. Acad. Sci. USA 1997, 94, 8842–8847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, B. Points of View: Color Blindness. Nat. Methods 2011, 8, 441. [Google Scholar] [CrossRef] [PubMed]
- Timotius, I.K.; Canneva, F.; Minakaki, G.; Pasluosta, C.; Moceri, S.; Casadei, N.; Riess, O.; Winkler, J.; Klucken, J.; von Hörsten, S.; et al. Dynamic footprint based locomotion sway assessment in α-synucleinopathic mice using Fast Fourier Transform and Low Pass Filter. J. Neurosci. Methods 2017, 296, 1–11. [Google Scholar] [CrossRef]
- Reznikoff, G.A.; Manaker, S.; Rhodes, C.H.; Winokur, A.; Rainbow, T.C. Localization and quantification of beta-adrenergic receptors in human brain. Neurology 1986, 36, 1067. [Google Scholar] [CrossRef] [PubMed]
- Yeh, H.; Woodward, D. Beta-1 adrenergic receptors mediate noradrenergic facilitation of purkinje cell responses to gamma-aminobutyric acid in cerebellum of rat. Neuropharmacology 1983, 22, 629–639. [Google Scholar] [CrossRef] [PubMed]
- Saitow, F.; Konishi, S. Excitability Increase Induced by β-Adrenergic Receptor-Mediated Activation of Hyperpolarization-Activated Cation Channels in Rat Cerebellar Basket Cells. J. Neurophysiol. 2000, 84, 2026–2034. [Google Scholar] [CrossRef]
- Palacios, J.; Kuhar, M.J. Beta adrenergic receptor localization in rat brain by light microscopic autoradiography. Neurochem. Int. 1982, 4, 473–490. [Google Scholar] [CrossRef]
- Rainbow, T.C.; Parsons, B.; Wolfe, B.B. Quantitative autoradiography of beta 1- and beta 2-adrenergic receptors in rat brain. Proc. Natl. Acad. Sci. USA 1984, 81, 1585–1589. [Google Scholar] [CrossRef] [Green Version]
- Chruscinski, A.J.; Rohrer, D.K.; Schauble, E.; Desai, K.H.; Bernstein, D.; Kobilka, B.K. Targeted Disruption of the Β2 Adrenergic Receptor Gene. J. Biol. Chem. 1999, 274, 16694–16700. [Google Scholar] [CrossRef] [Green Version]
- Rohrer, D.K.; Desai, K.H.; Jasper, J.R.; E Stevens, M.; Regula, D.P.; Barsh, G.S.; Bernstein, D.; Kobilka, B.K. Targeted disruption of the mouse beta1-adrenergic receptor gene: Developmental and cardiovascular effects. Proc. Natl. Acad. Sci. USA 1996, 93, 7375–7380. [Google Scholar] [CrossRef] [Green Version]
- Zilm, D.H.; Sellers, E.M. The effect of propranolol on normal physiologic tremor. Electroencephalogr. Clin. Neurophysiol. 1976, 41, 310–313. [Google Scholar] [CrossRef] [PubMed]
- Su, D.; Yang, S.; Hu, W.; Wang, D.; Kou, W.; Liu, Z.; Wang, X.; Wang, Y.; Ma, H.; Sui, Y.; et al. The Characteristics of Tremor Motion Help Identify Parkinson’s Disease and Multiple System Atrophy. Front. Neurol. 2020, 11, 540. [Google Scholar] [CrossRef]
- Benito-León, J.; Serrano, J.I.; Louis, E.D.; Holobar, A.; Romero, J.P.; Povalej-Bržan, P.; Kranjec, J.; Bermejo-Pareja, F.; Del Castillo, M.D.; Posada, I.J.; et al. Essential tremor severity and anatomical changes in brain areas controlling movement sequencing. Ann. Clin. Transl. Neurol. 2018, 6, 83–97. [Google Scholar] [CrossRef]
- Wang, S.; Aziz, T.Z.; Stein, J.F.; Bain, P.G.; Liu, X. Physiological and harmonic components in neural and muscular coherence in Parkinsonian tremor. Clin. Neurophysiol. 2006, 117, 1487–1498. [Google Scholar] [CrossRef] [PubMed]
- Farmer, S.; Bremner, F.D.; Halliday, D.; Rosenberg, J.R.; A Stephens, J. The frequency content of common synaptic inputs to motoneurones studied during voluntary isometric contraction in man. J. Physiol. 1993, 470, 127–155. [Google Scholar] [CrossRef] [PubMed]
- Reitsma, W. The Relation between Muscle Oscillations and Alternating Movements—Der Zusammenhang zwischen den Muskelschwingungen und den alternierenden Bewegungen. Biomed. Eng./Biomed. Tech. 1995, 40, 137–149. [Google Scholar] [CrossRef]
- Morgan, J.C.; Kurek, J.A.; Davis, J.L.; Sethi, K.D. Insights into Pathophysiology from Medication-induced Tremor. Tremor Other Hyperkinetic Movements 2017, 7. [Google Scholar] [CrossRef]
- Jefferson, D.; Jenner, P.; Marsden, C. Relationship between plasma propranolol levels and the clinical suppression of essential tremor [proceedings]. Br. J. Clin. Pharmacol. 1979, 7, 419P–420P. [Google Scholar] [CrossRef] [PubMed]
- Martin, F.C.; Le, A.T.; Handforth, A. Harmaline-induced tremor as a potential preclinical screening method for essential tremor medications. Mov. Disord. 2005, 20, 298–305. [Google Scholar] [CrossRef]
- Kralic, J.E.; Homanics, G.E.; Morrow, A.L. Genetic Essential Tremor in g g -Aminobutyric Acid A Receptor a a 1 Subunit Knockout Mice. J. Clin. Investig. 2005, 115. [Google Scholar] [CrossRef] [Green Version]
- Paterson, N.E.; Malekiani, S.A.; Foreman, M.M.; Olivier, B.; Hanania, T. Pharmacological Characterization of Harmaline-Induced Tremor Activity in Mice. Eur. J. Pharmacol. 2009, 616, 73–80. [Google Scholar] [CrossRef]
- Braham, J.; Sadeh, M.; Turgman, J.; Sarova-Pinchas, I. Beneficial effect of propranolol in familial ataxia. Ann. Neurol. 1979, 5, 207. [Google Scholar] [CrossRef]
- Duhigg, W.J. Effects of Propranolol on Ataxic Syndromes. Arch. Neurol. 1985, 42, 15. [Google Scholar] [CrossRef] [PubMed]
- Paparella, G.; Ferrazzano, G.; Cannavacciuolo, A.; Dezza, F.C.; Fabbrini, G.; Bologna, M.; Berardelli, A. Differential effects of propranolol on head and upper limb tremor in patients with essential tremor and dystonia. J. Neurol. 2018, 265, 2695–2703. [Google Scholar] [CrossRef] [PubMed]
- Volkind, R.A.; Llinás, R. The olivo-cerebellar system: Functional properties as revealed by harmaline-induced tremor. Exp. Brain Res. 1973, 18, 69–87. [Google Scholar] [CrossRef]
- Diener, H.-C.; Dichgans, J. Pathophysiology of cerebellar ataxia. Mov. Disord. 1992, 7, 95–109. [Google Scholar] [CrossRef] [PubMed]
- Deuschl, G.; Wenzelburger, R.; Löffler, K.; Raethjen, J.; Stolze, H. Essential tremor and cerebellar dysfunction Clinical and kinematic analysis of intention tremor. Brain 2000, 123, 1568–1580. [Google Scholar] [CrossRef] [Green Version]
- Schmahmann, J.D. Disorders of the Cerebellum: Ataxia, Dysmetria of Thought, and the Cerebellar Cognitive Affective Syndrome. J. Neuropsychiatry 2004, 16, 367–378. [Google Scholar] [CrossRef]
- Prudente, C.; Hess, E.; Jinnah, H. Dystonia as a network disorder: What is the role of the cerebellum? Neuroscience 2013, 260, 23–35. [Google Scholar] [CrossRef] [Green Version]
- Louis, E.D. Linking Essential Tremor to the Cerebellum: Neuropathological Evidence. Cerebellum 2015, 15, 235–242. [Google Scholar] [CrossRef]
- Shakkottai, V.G.; Batla, A.; Bhatia, K.; Dauer, W.T.; Dresel, C.; Niethammer, M.; Eidelberg, D.; Raike, R.S.; Smith, Y.; Jinnah, H.A.; et al. Current Opinions and Areas of Consensus on the Role of the Cerebellum in Dystonia. Cerebellum 2016, 16, 577–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laurens, C.; Abot, A.; Delarue, A.; Knauf, C. Central Effects of Beta-Blockers May Be Due to Nitric Oxide and Hydrogen Peroxide Release Independently of Their Ability to Cross the Blood-Brain Barrier. Front. Neurosci. 2019, 13, 33. [Google Scholar] [CrossRef]
- Cahill, L.; Pham, C.A.; Setlow, B. Impaired Memory Consolidation in Rats Produced with β-Adrenergic Blockade. Neurobiol. Learn. Mem. 2000, 74, 259–266. [Google Scholar] [CrossRef] [Green Version]
- Lonergan, M.; Olivera-Figueroa, L.; Pitman, R.; Brunet, A. Propranolol’s effects on the consolidation and reconsolidation of long-term emotional memory in healthy participants: A meta-analysis. J. Psychiatry Neurosci. 2013, 38, 222–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castonguay, C.-E.; Liao, C.; Khayachi, A.; Liu, Y.; Medeiros, M.; Houle, G.; Ross, J.P.; Dion, P.A.; Rouleau, G.A. Transcriptomic effects of propranolol and primidone converge on molecular pathways relevant to essential tremor. npj Genom. Med. 2022, 7, 46. [Google Scholar] [CrossRef]
- Rice, M.E. H2O2. Neurosci. 2011, 17, 389–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Picón-Pagès, P.; Garcia-Buendia, J.; Muñoz, F.J. Functions and dysfunctions of nitric oxide in brain. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2018, 1865, 1949–1967. [Google Scholar] [CrossRef]
- Handforth, A.; Lang, E.J. Increased Purkinje Cell Complex Spike and Deep Cerebellar Nucleus Synchrony as a Potential Basis for Syndromic Essential Tremor. A Review and Synthesis of the Literature. Cerebellum 2020, 20, 266–281. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Santaniello, S. Role of cerebellar GABAergic dysfunctions in the origins of essential tremor. Proc. Natl. Acad. Sci. USA 2019, 116, 13592–13601. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.-Y.; Louis, E.D.; Faust, P.L.; Koeppen, A.H.; Vonsattel, J.-P.G.; Kuo, S.-H. Abnormal climbing fibre-Purkinje cell synaptic connections in the essential tremor cerebellum. Brain 2014, 137, 3149–3159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Person, A.L.; Raman, I.M. Synchrony and neural coding in cerebellar circuits. Front. Neural Circuits 2012, 6, 97. [Google Scholar] [CrossRef] [Green Version]
- Armstrong, D.M.; A Edgley, S. Discharges of nucleus interpositus neurones during locomotion in the cat. J. Physiol. 1984, 351, 411–432. [Google Scholar] [CrossRef]
- Thach, W.T. Discharge of cerebellar neurons related to two maintained postures and two prompt movements. I. Nuclear cell output. J. Neurophysiol. 1970, 33, 527–536. [Google Scholar] [CrossRef] [PubMed]
- Hoebeek, F.E.; Witter, L.; Ruigrok, T.J.H.; De Zeeuw, C.I. Differential olivo-cerebellar cortical control of rebound activity in the cerebellar nuclei. Proc. Natl. Acad. Sci. USA 2010, 107, 8410–8415. [Google Scholar] [CrossRef] [Green Version]
- De Zeeuw, C.I.; Hoebeek, F.E.; Bosman, L.; Schonewille, M.; Witter, L.; Koekkoek, S.K. Spatiotemporal firing patterns in the cerebellum. Nat. Rev. Neurosci. 2011, 12, 327–344. [Google Scholar] [CrossRef]
- Elble, R.J. Central Mechanisms of Tremor. J. Clin. Neurophysiol. 1996, 13, 133–144. [Google Scholar] [CrossRef] [PubMed]
- Hallett, M. Overview of human tremor physiology. Mov. Disord. 2008, 13, 43–48. [Google Scholar] [CrossRef]
- Raethjen, J.; Deuschl, G. The oscillating central network of Essential tremor. Clin. Neurophysiol. 2012, 123, 61–64. [Google Scholar] [CrossRef]
- Louis, E.D. Essential tremor then and now: How views of the most common tremor diathesis have changed over time. Park. Relat. Disord. 2017, 46, S70–S74. [Google Scholar] [CrossRef] [PubMed]
- Stiles, G.L.; Caron, M.G.; Lefkowitz, R.J. Beta-adrenergic receptors: Biochemical mechanisms of physiological regulation. Physiol. Rev. 1984, 64, 661–743. [Google Scholar] [CrossRef] [PubMed]
- Barnes, P.J. Beta-adrenergic receptors and their regulation. Am. J. Respir. Crit. Care Med. 1995, 152, 838–860. [Google Scholar] [CrossRef] [PubMed]
- Collins, S. The -Adrenergic Receptors and the Control of Adipose Tissue Metabolism and Thermogenesis. Recent Prog. Horm. Res. 2001, 56, 309–328. [Google Scholar] [CrossRef] [PubMed]
- Lynch, G.S.; Ryall, J.G. Role of β-Adrenoceptor Signaling in Skeletal Muscle: Implications for Muscle Wasting and Disease. Physiol. Rev. 2008, 88, 729–767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oostland, M.; Buijink, M.R.; van Hooft, J.A. Serotonergic control of Purkinje cell maturation and climbing fibre elimination by 5-HT3receptors in the juvenile mouse cerebellum. J. Physiol. 2013, 591, 1793–1807. [Google Scholar] [CrossRef]
- Mitoma, H.; Konishi, S. Long-lasting facilitation of inhibitory transmission by monoaminergic and cAMP-dependent mechanism in rat cerebellar GABAergic synapses. Neurosci. Lett. 1996, 217, 141–144. [Google Scholar] [CrossRef]
- Tritos, N.; Kitraki, E.; Phillipidis, H.; Stylianopoulou, F. β-Adrenergic Receptors Mediate a Stress-Induced Decrease in IGF-II mRNA in the Rat Cerebellum. Cell. Mol. Neurobiol. 1998, 18, 525–534. [Google Scholar] [CrossRef]
- Cartford, M.C.; Samec, A.; Fister, M.; Bickford, P.C. Cerebellar norepinephrine modulates learning of delay classical eyeblink conditioning: Evidence for post-synaptic signaling via PKA. Learn. Mem. 2004, 11, 732–737. [Google Scholar] [CrossRef] [Green Version]
- Drengler, S.M.; Lorden, J.F.; Billitz, M.S.; A Oltmans, G. Adrenergic agents inhibit rapid increases in cerebellar Purkinje cell glutamic acid decarboxylase (GAD67) mRNA levels after climbing fiber lesions or reserpine treatment. J. Neurosci. 1996, 16, 1844–1851. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, J.; Van der Heijden, M.E.; Salazar Leon, L.E.; Lin, T.; Miterko, L.N.; Kizek, D.J.; Perez, R.M.; Pavešković, M.; Brown, A.M.; Sillitoe, R.V. Propranolol Modulates Cerebellar Circuit Activity and Reduces Tremor. Cells 2022, 11, 3889. https://doi.org/10.3390/cells11233889
Zhou J, Van der Heijden ME, Salazar Leon LE, Lin T, Miterko LN, Kizek DJ, Perez RM, Pavešković M, Brown AM, Sillitoe RV. Propranolol Modulates Cerebellar Circuit Activity and Reduces Tremor. Cells. 2022; 11(23):3889. https://doi.org/10.3390/cells11233889
Chicago/Turabian StyleZhou, Joy, Meike E. Van der Heijden, Luis E. Salazar Leon, Tao Lin, Lauren N. Miterko, Dominic J. Kizek, Ross M. Perez, Matea Pavešković, Amanda M. Brown, and Roy V. Sillitoe. 2022. "Propranolol Modulates Cerebellar Circuit Activity and Reduces Tremor" Cells 11, no. 23: 3889. https://doi.org/10.3390/cells11233889
APA StyleZhou, J., Van der Heijden, M. E., Salazar Leon, L. E., Lin, T., Miterko, L. N., Kizek, D. J., Perez, R. M., Pavešković, M., Brown, A. M., & Sillitoe, R. V. (2022). Propranolol Modulates Cerebellar Circuit Activity and Reduces Tremor. Cells, 11(23), 3889. https://doi.org/10.3390/cells11233889