
What Have We Learned in 30 Years of Investigations on Bari Transposons?


Abstract
:1. Introduction
The Discovery of the Bari1 Transposon: An Historical Overview
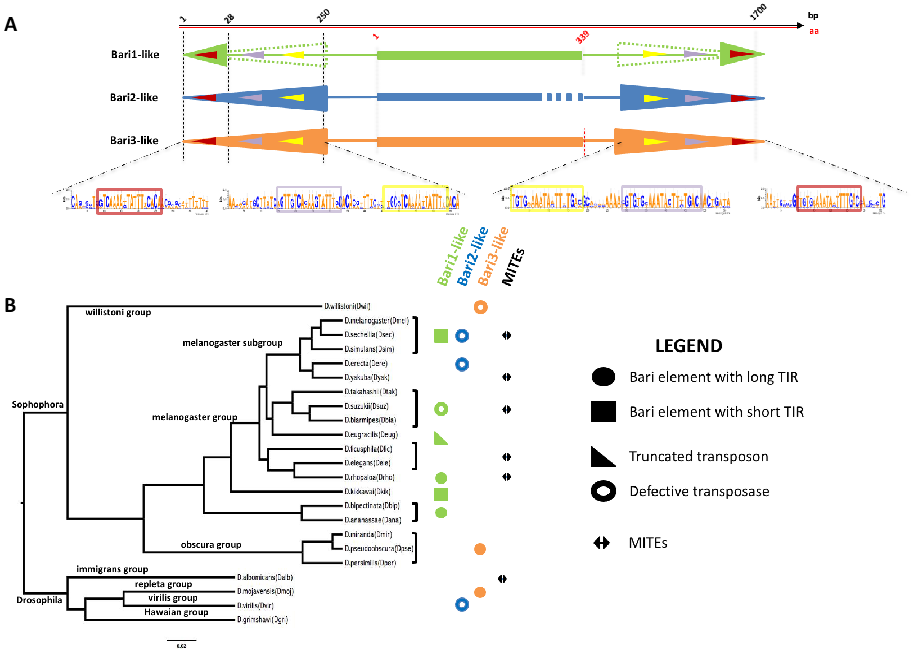
2. The Bari Family Grows Up
2.1. Introducing Bari2 and Bari3
2.2. New Cognate Elements in New Species: The Crew Grows up in the Post-Genomic Era
3. Horizontal Transfer Events Involving Bari Elements
4. The Missing Jump to the Next Level
4.1. Why We Cannot Use Bari Transposons as Tools for Chromosomal Integration
4.2. Active or Non-Active? That Is the Question
5. What Do We Know about the Regulation Bari Transposons?
Bari Transposons Regulation Relies on the piRNA Pathways
6. Do Bari Transposons Have a Role in The Genome?
6.1. Contribution in Creating Somatic Mosaicism and Adaptive Insertions
6.2. Possible Role in Chromatin Assembly
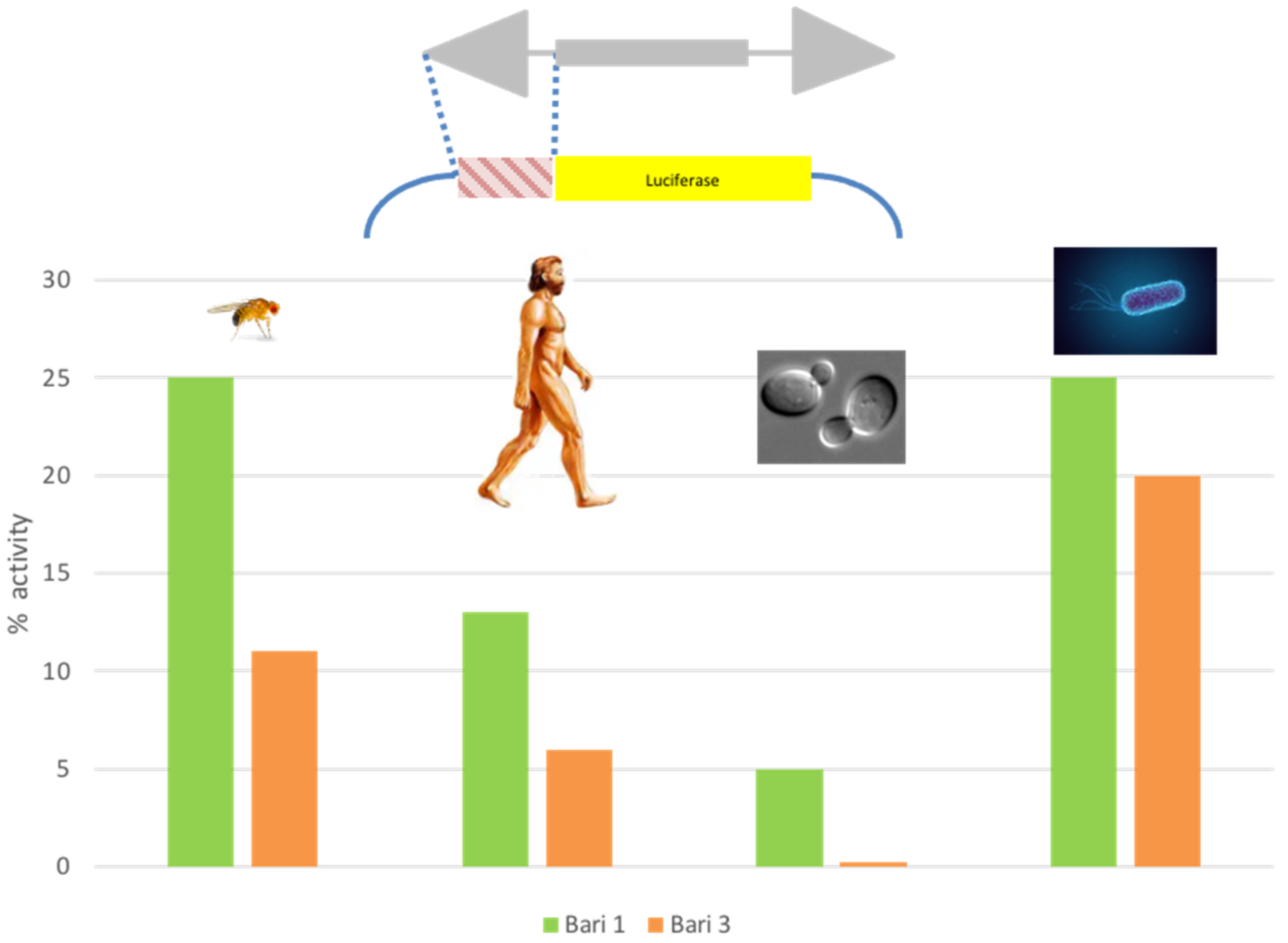
7. The Blurry Promoter: Who Wants to Live Forever?
8. Concluding Remarks
What We Have Learned and What We Still Have to Learn from Bari Transposons
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sandoval-Villegas, N.; Nurieva, W.; Amberger, M.; Ivics, Z. Contemporary Transposon Tools: A Review and Guide through Mechanisms and Applications of Sleeping Beauty, piggyBac and Tol2 for Genome Engineering. Int. J. Mol. Sci. 2021, 22, 5084. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Burnight, E.R.; Cooney, A.L.; Malani, N.; Brady, T.; Sander, J.D.; Staber, J.; Wheelan, S.J.; Joung, J.K.; McCray, P.B., Jr.; et al. piggyBac transposase tools for genome engineering. Proc. Natl. Acad. Sci. USA 2013, 110, E2279–E2287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kesselring, L.; Miskey, C.; Zuliani, C.; Querques, I.; Kapitonov, V.; Laukó, A.; Fehér, A.; Palazzo, A.; Diem, T.; Lustig, J.; et al. A single amino acid switch converts the Sleeping Beauty transposase into an efficient unidirectional excisionase with utility in stem cell reprogramming. Nucleic Acids Res. 2020, 48, 316–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palazzo, A.; Marsano, R.M. Transposable elements: A jump toward the future of expression vectors. Crit. Rev. Biotechnol. 2021, 41, 1–27. [Google Scholar] [CrossRef]
- Rubin, G.M.; Spradling, A.C. Genetic transformation of Drosophila with transposable element vectors. Science 1982, 218, 348–353. [Google Scholar] [CrossRef] [Green Version]
- Spradling, A.C.; Rubin, G.M. Transposition of cloned P elements into Drosophila germ line chromosomes. Science 1982, 218, 341–347. [Google Scholar] [CrossRef] [Green Version]
- O’Brochta, D.A.; Handler, A.M. Mobility of P elements in drosophilids and nondrosophilids. Proc. Natl. Acad. Sci. USA 1988, 85, 6052–6056. [Google Scholar] [CrossRef] [Green Version]
- Tenzen, T.; Matsutani, S.; Ohtsubo, E. Site-specific transposition of insertion sequence IS630. J. Bacteriol. 1990, 172, 3830–3836. [Google Scholar] [CrossRef] [Green Version]
- Plasterk, R.H.A. The Tc1/mariner Transposon Family. In Transposable Elements; Saedler, H., Gierl, A., Eds.; Springer: Berlin/Heidelberg, Germany, 1996; pp. 125–143. [Google Scholar]
- Miskey, C.; Izsvak, Z.; Kawakami, K.; Ivics, Z. DNA transposons in vertebrate functional genomics. Cell Mol. Life Sci. 2005, 62, 629–641. [Google Scholar] [CrossRef]
- Prommersberger, S.; Reiser, M.; Beckmann, J.; Danhof, S.; Amberger, M.; Quade-Lyssy, P.; Einsele, H.; Hudecek, M.; Bonig, H.; Ivics, Z. CARAMBA: A first-in-human clinical trial with SLAMF7 CAR-T cells prepared by virus-free Sleeping Beauty gene transfer to treat multiple myeloma. Gene Ther. 2021, 28, 560–571. [Google Scholar] [CrossRef]
- Wicker, T.; Gundlach, H.; Spannagl, M.; Uauy, C.; Borrill, P.; Ramírez-González, R.H.; De Oliveira, R.; Mayer, K.F.X.; Paux, E.; Choulet, F.; et al. Impact of transposable elements on genome structure and evolution in bread wheat. Genome Biol. 2018, 19, 103. [Google Scholar] [CrossRef]
- Bourque, G.; Burns, K.H.; Gehring, M.; Gorbunova, V.; Seluanov, A.; Hammell, M.; Imbeault, M.; Izsvák, Z.; Levin, H.L.; Macfarlan, T.S.; et al. Ten things you should know about transposable elements. Genome Biol. 2018, 19, 199. [Google Scholar] [CrossRef]
- Cain, A.K.; Barquist, L.; Goodman, A.L.; Paulsen, I.T.; Parkhill, J.; Van Opijnen, T. A decade of advances in transposon-insertion sequencing. Nat. Rev. Genet. 2020, 21, 526–540. [Google Scholar] [CrossRef]
- Kaya-Okur, H.S.; Wu, S.J.; Codomo, C.A.; Pledger, E.S.; Bryson, T.D.; Henikoff, J.G.; Ahmad, K.; Henikoff, S. CUT&Tag for efficient epigenomic profiling of small samples and single cells. Nat. Commun. 2019, 10, 1930. [Google Scholar] [CrossRef] [Green Version]
- Caggese, C.; Caizzi, R.; Bozzetti, M.P.; Barsanti, P.; Ritossa, F. Genetic determinants of glutamine synthetase in Drosophila melanogaster: A gene for glutamine synthetase I resides in the 21B3-6 region. Biochem. Genet. 1988, 26, 571–584. [Google Scholar] [CrossRef]
- Caggese, C.; Caizzi, R.; Barsanti, P.; Bozzetti, M.P. Mutations in the glutamine synthetase I (gsI) gene produce embryo-lethal female sterility in Drosophila melanogaster. Dev. Genet. 1992, 13, 359–366. [Google Scholar] [CrossRef]
- Caizzi, R.; Bozzetti, M.P.; Caggese, C.; Ritossa, F. Homologous nuclear genes encode cytoplasmic and mitochondrial glutamine synthetase in Drosophila melanogaster. J. Mol. Biol. 1990, 212, 17–26. [Google Scholar] [CrossRef]
- Pimpinelli, S.; Dimitri, P. Cytogenetic analysis of segregation distortion in Drosophila melanogaster: The cytological organization of the Responder (Rsp) locus. Genetics 1989, 121, 765–772. [Google Scholar] [CrossRef]
- Wu, C.I.; True, J.R.; Johnson, N. Fitness reduction associated with the deletion of a satellite DNA array. Nature 1989, 341, 248–251. [Google Scholar] [CrossRef]
- Wu, C.-I.; Lyttle, T.W.; Wu, M.-L.; Lin, G.-F. Association between a satellite DNA sequence and the responder of segregation distorter in D. melanogaster. Cell 1988, 54, 179–189. [Google Scholar] [CrossRef]
- Brittnacher, J.G.; Ganetzky, B. On the components of segregation distortion in Drosophila melanogaster. IV. Construction and analysis of free duplications for the Responder locus. Genetics 1989, 121, 739–750. [Google Scholar] [CrossRef] [PubMed]
- Ganetzky, B. On the components of segregation distortion in Drosophila melanogaster. Genetics 1977, 86, 321–355. [Google Scholar] [CrossRef] [PubMed]
- Moschetti, R.; Chlamydas, S.; Marsano, R.M.; Caizzi, R. Conserved motifs and dynamic aspects of the terminal inverted repeat organization within Bari-like transposons. Mol. Genet. Genom. 2008, 279, 451–461. [Google Scholar] [CrossRef] [PubMed]
- Plasterk, R.H.; Izsvák, Z.; Ivics, Z. Resident aliens: The Tc1/mariner superfamily of transposable elements. Trends Genet. 1999, 15, 326–332. [Google Scholar] [CrossRef]
- Petrov, D.A.; Schutzman, J.L.; Hartl, D.L.; Lozovskaya, E.R. Diverse transposable elements are mobilized in hybrid dysgenesis in Drosophila virilis. Proc. Natl. Acad. Sci. USA 1995, 92, 8050–8054. [Google Scholar] [CrossRef] [Green Version]
- Merriman, P.J.; Grimes, C.D.; Ambroziak, J.; Hackett, D.A.; Skinner, P.; Simmons, M.J. S elements: A family of Tc1-like transposons in the genome of Drosophila melanogaster. Genetics 1995, 141, 1425–1438. [Google Scholar] [CrossRef]
- Arcà, B.; Savakis, C. Distribution of the Transposable Element Minos in the Genus Drosophila. Genetica 2000, 108, 263–267. [Google Scholar] [CrossRef] [Green Version]
- Izsvak, Z.; Khare, D.; Behlke, J.; Heinemann, U.; Plasterk, R.H.; Ivics, Z. Involvement of a bifunctional, paired-like DNA-binding domain and a transpositional enhancer in Sleeping Beauty transposition. J. Biol. Chem. 2002, 277, 34581–34588. [Google Scholar] [CrossRef] [Green Version]
- Palazzo, A.; Lovero, D.; D’Addabbo, P.; Caizzi, R.; Marsano, R.M. Identification of Bari Transposons in 23 Sequenced Drosophila Genomes Reveals Novel Structural Variants, MITEs and Horizontal Transfer. PLoS ONE 2016, 11, e0156014. [Google Scholar] [CrossRef] [Green Version]
- McGurk, M.P.; Barbash, D.A. Double insertion of transposable elements provides a substrate for the evolution of satellite DNA. Genome Res. 2018, 28, 714–725. [Google Scholar] [CrossRef] [Green Version]
- Caizzi, R.; Caggese, C.; Pimpinelli, S. Bari-1, a new transposon-like family in Drosophila melanogaster with a unique heterochromatic organization. Genetics 1993, 133, 335–345. [Google Scholar] [CrossRef]
- Caggese, C.; Pimpinelli, S.; Barsanti, P.; Caizzi, R. The distribution of the transposable element Bari-1 in the Drosophila melanogaster and Drosophila simulans genomes. Genetica 1995, 96, 269–283. [Google Scholar] [CrossRef]
- Marsano, R.M.; Milano, R.; Minervini, C.; Moschetti, R.; Caggese, C.; Barsanti, P.; Caizzi, R. Organization and possible origin of the Bari-1 cluster in the heterochromatic h39 region of Drosophila melanogaster. Genetica 2003, 117, 281–289. [Google Scholar] [CrossRef]
- Berloco, M.F.; Minervini, C.F.; Moschetti, R.; Palazzo, A.; Viggiano, L.; Marsano, R.M. Evidence of the Physical Interaction between Rpl22 and the Transposable Element Doc5, a Heterochromatic Transposon of Drosophila melanogaster. Genes 2021, 12, 1997. [Google Scholar] [CrossRef]
- Marsano, R.M.; Marconi, S.; Moschetti, R.; Barsanti, P.; Caggese, C.; Caizzi, R. MAX, a novel retrotransposon of the BEL-Pao family, is nested within the Bari1 cluster at the heterochromatic h39 region of chromosome 2 in Drosophila melanogaster. Mol. Genet. Genom. 2004, 270, 477–484. [Google Scholar] [CrossRef]
- Marsano, R.M.; Moschetti, R.; Barsanti, P.; Caggese, C.; Caizzi, R. A survey of the DNA sequences surrounding the Bari1 repeats in the pericentromeric h39 region of Drosophila melanogaster. Gene 2003, 307, 167–174. [Google Scholar] [CrossRef]
- Crooks, G.E.; Hon, G.; Chandonia, J.M.; Brenner, S.E. WebLogo: A sequence logo generator. Genome Res. 2004, 14, 1188–1190. [Google Scholar] [CrossRef] [Green Version]
- Moschetti, R.; Caggese, C.; Barsanti, P.; Caizzi, R. Intra- and interspecies variation among Bari-1 elements of the melanogaster species group. Genetics 1998, 150, 239–250. [Google Scholar] [CrossRef]
- Hartl, D.L.; Lozovskaya, E.R.; Nurminsky, D.I.; Lohe, A.R. What restricts the activity of mariner-like transposable elements. Trends Genet. 1997, 13, 197–201. [Google Scholar] [CrossRef]
- Claeys Bouuaert, C.; Chalmers, R.M. Gene therapy vectors: The prospects and potentials of the cut-and-paste transposons. Genetica 2010, 138, 473–484. [Google Scholar] [CrossRef]
- Palazzo, A.; Moschetti, R.; Caizzi, R.; Marsano, R.M. The Drosophila mojavensis Bari3 transposon: Distribution and functional characterization. Mob. DNA 2014, 5, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bureau, T.E.; Wessler, S.R. Tourist: A large family of small inverted repeat elements frequently associated with maize genes. Plant Cell 1992, 4, 1283–1294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, N.; Feschotte, C.; Zhang, X.; Wessler, S.R. Using rice to understand the origin and amplification of miniature inverted repeat transposable elements (MITEs). Curr. Opin. Plant Biol. 2004, 7, 115–119. [Google Scholar] [CrossRef] [PubMed]
- Quesneville, H.; Nouaud, D.; Anxolabéhère, D. P elements and MITE relatives in the whole genome sequence of Anopheles gambiae. BMC Genom. 2006, 7, 214. [Google Scholar] [CrossRef] [Green Version]
- De Freitas Ortiz, M.; Silva Loreto, E.L. The hobo-related elements in the melanogaster species group. Genet. Res. 2008, 90, 243–252. [Google Scholar] [CrossRef]
- Dias, E.S.; Carareto, C.M. msechBari, a new MITE-like element in Drosophila sechellia related to the Bari transposon. Genet. Res. 2011, 93, 381–385. [Google Scholar] [CrossRef] [Green Version]
- Keeling, P.J. Functional and ecological impacts of horizontal gene transfer in eukaryotes. Curr. Opin. Genet. Dev. 2009, 19, 613–619. [Google Scholar] [CrossRef]
- Boto, L. Horizontal gene transfer in the acquisition of novel traits by metazoans. Proc. Biol. Sci. 2014, 281, 20132450. [Google Scholar] [CrossRef] [Green Version]
- Schaack, S.; Gilbert, C.; Feschotte, C. Promiscuous DNA: Horizontal transfer of transposable elements and why it matters for eukaryotic evolution. Trends Evol. 2010, 25, 537–546. [Google Scholar] [CrossRef] [Green Version]
- Wallau, G.L.; Vieira, C.; Loreto, É.L.S. Genetic exchange in eukaryotes through horizontal transfer: Connected by the mobilome. Mob. DNA 2018, 9, 6. [Google Scholar] [CrossRef]
- Wallau, G.L.; Capy, P.; Loreto, E.; Le Rouzic, A.; Hua-Van, A. VHICA, a New Method to Discriminate between Vertical and Horizontal Transposon Transfer: Application to the Mariner Family within Drosophila. Mol. Biol. Evol. 2016, 33, 1094–1109. [Google Scholar] [CrossRef] [Green Version]
- Dias, E.S.; Carareto, C.M. Ancestral polymorphism and recent invasion of transposable elements in Drosophila species. BMC Evol. Biol. 2012, 12, 119. [Google Scholar] [CrossRef] [Green Version]
- Bartolome, C.; Bello, X.; Maside, X. Widespread evidence for horizontal transfer of transposable elements across Drosophila genomes. Genome Biol. 2009, 10, R22. [Google Scholar] [CrossRef] [Green Version]
- Ometto, L.; Cestaro, A.; Ramasamy, S.; Grassi, A.; Revadi, S.; Siozios, S.; Moretto, M.; Fontana, P.; Varotto, C.; Pisani, D.; et al. Linking genomics and ecology to investigate the complex evolution of an invasive Drosophila pest. Genome Biol. Evol. 2013, 5, 745–757. [Google Scholar] [CrossRef] [Green Version]
- Moschetti, R.; Marsano, R.M.; Barsanti, P.; Caggese, C.; Caizzi, R. FB elements can promote exon shuffling: A promoter-less white allele can be reactivated by FB mediated transposition in Drosophila melanogaster. Mol. Genet. Genom. 2004, 271, 394–401. [Google Scholar] [CrossRef]
- Capy, P.; Gasperi, G.; Biémont, C.; Bazin, C. Stress and transposable elements: Co-evolution or useful parasites? Heredity 2000, 85, 101–106. [Google Scholar] [CrossRef]
- Feng, G.; Leem, Y.-E.; Levin, H.L. Transposon integration enhances expression of stress response genes. Nucleic Acids Res. 2013, 41, 775–789. [Google Scholar] [CrossRef] [Green Version]
- Rech, G.E.; Bogaerts-Márquez, M.; Barrón, M.G.; Merenciano, M.; Villanueva-Cañas, J.L.; Horváth, V.; Fiston-Lavier, A.-S.; Luyten, I.; Venkataram, S.; Quesneville, H.; et al. Stress response, behavior, and development are shaped by transposable element-induced mutations in Drosophila. PLoS Genet. 2019, 15, e1007900. [Google Scholar] [CrossRef] [Green Version]
- Horváth, V.; Merenciano, M.; González, J. Revisiting the Relationship between Transposable Elements and the Eukaryotic Stress Response. Trends Genet. 2017, 33, 832–841. [Google Scholar] [CrossRef]
- Palazzo, A.; Marconi, S.; Specchia, V.; Bozzetti, M.P.; Ivics, Z.; Caizzi, R.; Marsano, R.M. Functional Characterization of the Bari1 Transposition System. PLoS ONE 2013, 8, e79385. [Google Scholar] [CrossRef] [Green Version]
- Junakovic, N.; Di Franco, C.; Terrinoni, A. Evidence for a host role in regulating the activity of transposable elements in Drosophila melanogaster: The case of the persistent instability of Bari 1 elements in Charolles stock. Genetica 1997, 100, 149–154. [Google Scholar] [CrossRef] [PubMed]
- Soriano, S.; Fortunati, D.; Junakovic, N. Evidence for the Host Contribution in the Definition of Preferential Insertion Sites of the Elements of Bari 1 Transposon Family in Drosophila melanogaster. J. Mol. Evol. 2002, 55, 606–615. [Google Scholar] [CrossRef] [PubMed]
- Marsano, R.M.; Caizzi, R.; Moschetti, R.; Junakovic, N. Evidence for a functional interaction between the Bari1 transposable element and the cytochrome P450 cyp12a4 gene in Drosophila melanogaster. Gene 2005, 357, 122–128. [Google Scholar] [CrossRef]
- Specchia, V.; Piacentini, L.; Tritto, P.; Fanti, L.; D’Alessandro, R.; Palumbo, G.; Pimpinelli, S.; Bozzetti, M.P. Hsp90 prevents phenotypic variation by suppressing the mutagenic activity of transposons. Nature 2010, 463, 662–665. [Google Scholar] [CrossRef] [PubMed]
- Specchia, V.; Bozzetti, M.P. The Role of HSP90 in Preserving the Integrity of Genomes Against Transposons Is Evolutionarily Conserved. Cells 2021, 10, 1096. [Google Scholar] [CrossRef]
- Pimpinelli, S.; Piacentini, L. Environmental change and the evolution of genomes: Transposable elements as translators of phenotypic plasticity into genotypic variability. Funct. Ecol. 2020, 34, 428–441. [Google Scholar] [CrossRef]
- Malone, C.D.; Hannon, G.J. Small RNAs as guardians of the genome. Cell 2009, 136, 656–668. [Google Scholar] [CrossRef] [Green Version]
- Brennecke, J.; Aravin, A.A.; Stark, A.; Dus, M.; Kellis, M.; Sachidanandam, R.; Hannon, G.J. Discrete small RNA-generating loci as master regulators of transposon activity in Drosophila. Cell 2007, 128, 1089–1103. [Google Scholar] [CrossRef] [Green Version]
- Grewal, S.I.S.; Elgin, S.C.R. Transcription and RNA interference in the formation of heterochromatin. Nature 2007, 447, 399–406. [Google Scholar] [CrossRef]
- Wang, S.H.; Elgin, S.C.R. Drosophila Piwi functions downstream of piRNA production mediating a chromatin-based transposon silencing mechanism in female germ line. Proc. Natl. Acad. Sci. USA 2011, 108, 21164–21169. [Google Scholar] [CrossRef] [Green Version]
- Le Thomas, A.; Rogers, A.K.; Webster, A.; Marinov, G.K.; Liao, S.E.; Perkins, E.M.; Hur, J.K.; Aravin, A.A.; Tóth, K.F. Piwi induces piRNA-guided transcriptional silencing and establishment of a repressive chromatin state. Genes Dev. 2013, 27, 390–399. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Ma, Z.; Niu, K.; Xiao, Y.; Wu, X.; Pan, C.; Zhao, Y.; Wang, K.; Zhang, Y.; Liu, N. Antagonistic roles of Nibbler and Hen1 in modulating piRNA 3′ ends in Drosophila. Development 2016, 143, 530–539. [Google Scholar] [CrossRef] [Green Version]
- Pritykin, Y.; Brito, T.; Schupbach, T.; Singh, M.; Pane, A. Integrative analysis unveils new functions for the Drosophila Cutoff protein in noncoding RNA biogenesis and gene regulation. RNA 2017, 23, 1097–1109. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Zhang, P.; Lu, Y.; Li, Y.; Zheng, Y.; Kan, Y.; Chen, R.; He, S. piRBase: A comprehensive database of piRNA sequences. Nucleic Acids Res. 2019, 47, D175–D180. [Google Scholar] [CrossRef] [Green Version]
- Chlamydas, S.; Heun, P.; Dimitri, P.; Moschetti, R.; Barsanti, P.; Caizzi, R. The paracentric inversion In(2Rh)PL alters the centromeric organization of chromosome 2 in Drosophila melanogaster. Chromosome Res. 2009, 17, 1–9. [Google Scholar] [CrossRef]
- Zhang, F.; Wang, J.; Xu, J.; Zhang, Z.; Koppetsch, B.S.; Schultz, N.; Vreven, T.; Meignin, C.; Davis, I.; Zamore, P.D.; et al. UAP56 couples piRNA clusters to the perinuclear transposon silencing machinery. Cell 2012, 151, 871–884. [Google Scholar] [CrossRef] [Green Version]
- Grentzinger, T.; Armenise, C.; Brun, C.; Mugat, B.; Serrano, V.; Pelisson, A.; Chambeyron, S. piRNA-mediated transgenerational inheritance of an acquired trait. Genome Res. 2012, 22, 1877–1888. [Google Scholar] [CrossRef] [Green Version]
- Mugat, B.; Akkouche, A.; Serrano, V.; Armenise, C.; Li, B.; Brun, C.; Fulga, T.A.; Van Vactor, D.; Pélisson, A.; Chambeyron, S. MicroRNA-Dependent Transcriptional Silencing of Transposable Elements in Drosophila Follicle Cells. PLoS Genet. 2015, 11, e1005194. [Google Scholar] [CrossRef] [Green Version]
- Olovnikov, I.; Ryazansky, S.; Shpiz, S.; Lavrov, S.; Abramov, Y.; Vaury, C.; Jensen, S.; Kalmykova, A. De novo piRNA cluster formation in the Drosophila germ line triggered by transgenes containing a transcribed transposon fragment. Nucleic Acids Res. 2013, 41, 5757–5768. [Google Scholar] [CrossRef] [Green Version]
- Feltzin, V.L.; Khaladkar, M.; Abe, M.; Parisi, M.; Hendriks, G.J.; Kim, J.; Bonini, N.M. The exonuclease Nibbler regulates age-associated traits and modulates piRNA length in Drosophila. Aging Cell 2015, 14, 443–452. [Google Scholar] [CrossRef]
- Chirn, G.W.; Rahman, R.; Sytnikova, Y.A.; Matts, J.A.; Zeng, M.; Gerlach, D.; Yu, M.; Berger, B.; Naramura, M.; Kile, B.T.; et al. Conserved piRNA Expression from a Distinct Set of piRNA Cluster Loci in Eutherian Mammals. PLoS Genet. 2015, 11, e1005652. [Google Scholar] [CrossRef] [PubMed]
- Shpiz, S.; Ryazansky, S.; Olovnikov, I.; Abramov, Y.; Kalmykova, A. Euchromatic transposon insertions trigger production of novel Pi- and endo-siRNAs at the target sites in the drosophila germline. PLoS Genet. 2014, 10, e1004138. [Google Scholar] [CrossRef] [PubMed]
- Loreto, E.L.S.; Pereira, C.M. Somatizing the transposons action. Mob. Genet. Elem. 2017, 7, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Perrat, P.N.; DasGupta, S.; Wang, J.; Theurkauf, W.; Weng, Z.; Rosbash, M.; Waddell, S. Transposition-driven genomic heterogeneity in the Drosophila brain. Science 2013, 340, 91–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Treiber, C.D.; Waddell, S. Resolving the prevalence of somatic transposition in Drosophila. Elife 2017, 6, e28297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Treiber, C.D.; Waddell, S. Transposon expression in the Drosophila brain is driven by neighboring genes and diversifies the neural transcriptome. bioRxiv 2019, 838045. [Google Scholar] [CrossRef] [PubMed]
- Siudeja, K.; Nassari, S.; Gervais, L.; Skorski, P.; Lameiras, S.; Stolfa, D.; Zande, M.; Bernard, V.; Frio, T.R.; Bardin, A.J. Frequent Somatic Mutation in Adult Intestinal Stem Cells Drives Neoplasia and Genetic Mosaicism during Aging. Cell Stem Cell 2015, 17, 663–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siudeja, K.; Van den Beek, M.; Riddiford, N.; Boumard, B.; Wurmser, A.; Stefanutti, M.; Lameiras, S.; Bardin, A.J. Unraveling the features of somatic transposition in the Drosophila intestine. Embo J. 2021, 40, e106388. [Google Scholar] [CrossRef]
- Adams, M.D.; Celniker, S.E.; Holt, R.A.; Evans, C.A.; Gocayne, J.D.; Amanatides, P.G.; Scherer, S.E.; Li, P.W.; Hoskins, R.A.; Galle, R.F.; et al. The genome sequence of Drosophila melanogaster. Science 2000, 287, 2185–2195. [Google Scholar] [CrossRef] [Green Version]
- Guio, L.; Barron, M.G.; Gonzalez, J. The transposable element Bari-Jheh mediates oxidative stress response in Drosophila. Mol. Ecol. 2014, 23, 2020–2030. [Google Scholar] [CrossRef]
- Guio, L.; Vieira, C.; Gonzalez, J. Stress affects the epigenetic marks added by natural transposable element insertions in Drosophila melanogaster. Sci. Rep. 2018, 8, 12197. [Google Scholar] [CrossRef] [Green Version]
- Slotkin, R.K.; Martienssen, R. Transposable elements and the epigenetic regulation of the genome. Nat. Rev. Genet. 2007, 8, 272–285. [Google Scholar] [CrossRef]
- Caizzi, R.; Moschetti, R.; Piacentini, L.; Fanti, L.; Marsano, R.M.; Dimitri, P. Comparative Genomic Analyses Provide New Insights into the Evolutionary Dynamics of Heterochromatin in Drosophila. PLoS Genet. 2016, 12, e1006212. [Google Scholar] [CrossRef]
- Moschetti, R.; Palazzo, A.; Lorusso, P.; Viggiano, L.; Marsano, R.M. “What You Need, Baby, I Got It”: Transposable Elements as Suppliers of Cis-Operating Sequences in Drosophila. Biology 2020, 9, 25. [Google Scholar] [CrossRef] [Green Version]
- Van Steensel, B.; Henikoff, S. Identification of in vivo DNA targets of chromatin proteins using tethered Dam methyltransferase. Nat. Biotechnol. 2000, 18, 424–428. [Google Scholar] [CrossRef]
- Van Steensel, B.; Delrow, J.; Henikoff, S. Chromatin profiling using targeted DNA adenine methyltransferase. Nat. Genet. 2001, 27, 304–308. [Google Scholar] [CrossRef] [Green Version]
- Casale, A.M.; Cappucci, U.; Piacentini, L. Unravelling HP1 functions: Post-transcriptional regulation of stem cell fate. Chromosoma 2021, 130, 103–111. [Google Scholar] [CrossRef]
- Brasier, A.R.; Ron, D. Luciferase reporter gene assay in mammalian cells. In Methods Enzymol; Academic Press: Cambridge, MA, USA, 1992; Volume 216, pp. 386–397. [Google Scholar]
- Palazzo, A.; Caizzi, R.; Viggiano, L.; Marsano, R.M. Does the Promoter Constitute a Barrier in the Horizontal Transposon Transfer Process? Insight from Bari Transposons. Genome Biol. Evol. 2017, 9, 1637–1645. [Google Scholar] [CrossRef] [Green Version]
- Hartl, D.L.; Lohe, A.R.; Lozovskaya, E.R. Regulation of the transposable element mariner. Genetica 1997, 100, 177–184. [Google Scholar] [CrossRef]
- Kanhere, A.; Bansal, M. Structural properties of promoters: Similarities and differences between prokaryotes and eukaryotes. Nucleic Acids 2005, 33, 3165–3175. [Google Scholar] [CrossRef] [Green Version]
- Silva, J.C.; Loreto, E.L.; Clark, J.B. Factors that affect the horizontal transfer of transposable elements. Curr. Issues Mol. Biol. 2004, 6, 57–71. [Google Scholar] [PubMed]
- Peccoud, J.; Loiseau, V.; Cordaux, R.; Gilbert, C. Massive horizontal transfer of transposable elements in insects. Proc. Natl. Acad. Sci. USA 2017, 114, 4721–4726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palazzo, A.; Lorusso, P.; Miskey, C.; Walisko, O.; Gerbino, A.; Marobbio, C.M.T.; Ivics, Z.; Marsano, R.M. Transcriptionally promiscuous “blurry” promoters in Tc1/mariner transposons allow transcription in distantly related genomes. Mob. DNA 2019, 10, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Name | Tissue | Genetic Background | Methods | Reads | Sequence | Length | Reference |
|---|---|---|---|---|---|---|---|
| piR-dme-2858217 | O; E | W; T; M | oxidized small RNA small RNA | 1–3 | TTTGACCACCTCTGGTCATGGTCAAAA | 27 | [77,78,79] |
| piR-dme-3826713 | O | W; T; M | small RNA oxidized small RNA | 1–6 | TCTGGTCATGGTCAAAATTATTTT | 24 | [77,79,80,81] |
| piR-dme-8496440 | O | T | small RNA | 1–3 | TTTGACCACCTCTGGTCATGGTCAA | 25 | [80] |
| piR-dme-13381112 | O | T | small RNA | 1 | CCACCTCTGGTCATGGTCAAAATTAT | 26 | [80] |
| piR-dme-21388569 | F; O | W; T | small RNA | 1–9 | TTTGACCACCTCTGGTCATGGTCAAAAT | 28 | [79,81,82] |
| piR-dme-21631816 | O | W | small RNA | 1 | CACCTCTGGTCATGGTCAAAATTAT | 25 | [83] |
| piR-dme-26496558 | O | T | small RNA | 1 | ACCACCTCTGGTCATGGTCAAAATTA | 26 | [79] |
| piR-dme-26779428 | O | T | small RNA | 1 | CCACCTCTGGTCATGGTCAAAAT | 23 | [79] |
| piR-dme-27814712 | O | T | small RNA | 1 | TCTGGTCATGGTCAAAATTATTT | 23 | [79] |
| piR-dme-29438648 | O | T | small RNA | 1 | TGACCACCTCTGGTCATGGTCAAA | 24 | [79] |
| piR-dme-29670403 | O | T | small RNA | 1 | TTGACCACCTCTGGTCATGGTCAAAA | 26 | [79] |
| piR-dme-30537191 | O | T | small RNA | 2 | TTTGACCACCTCTGGTCATGGTCAAA | 26 | [79] |
| piR-dme-31705044 | O | T | small RNA | 1 | CTCTGGTCATGGTCAAAATTATTT | 24 | [79] |
| piR-dme-32470189 | O | W | small RNA | 1 | TTTGACCACCTCTGGTCATGGTCA | 24 | [81] |
| piR-dme-33774286 | O | W | small RNA | 1 | TGACCACCTCTGGTCATGGTCAAAAT | 26 | [81] |
| piR-dme-38817646 | O | T | small RNA | 1 | CTCTGGTCATGGTCAAAATTATTTT | 25 | [81] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Palazzo, A.; Caizzi, R.; Moschetti, R.; Marsano, R.M. What Have We Learned in 30 Years of Investigations on Bari Transposons? Cells 2022, 11, 583. https://doi.org/10.3390/cells11030583
Palazzo A, Caizzi R, Moschetti R, Marsano RM. What Have We Learned in 30 Years of Investigations on Bari Transposons? Cells. 2022; 11(3):583. https://doi.org/10.3390/cells11030583
Chicago/Turabian StylePalazzo, Antonio, Ruggiero Caizzi, Roberta Moschetti, and René Massimiliano Marsano. 2022. "What Have We Learned in 30 Years of Investigations on Bari Transposons?" Cells 11, no. 3: 583. https://doi.org/10.3390/cells11030583
APA StylePalazzo, A., Caizzi, R., Moschetti, R., & Marsano, R. M. (2022). What Have We Learned in 30 Years of Investigations on Bari Transposons? Cells, 11(3), 583. https://doi.org/10.3390/cells11030583