
Morphological and Immunopathological Aspects of Lingual Tissues in COVID-19



 , , ,
, , ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Specimen Processing
2.2.1. Molecular Processing
2.2.2. Histological Processing and Morphological Analysis
3. Results
3.1. Study Population
3.2. COVID-19+
3.2.1. Molecular Analysis
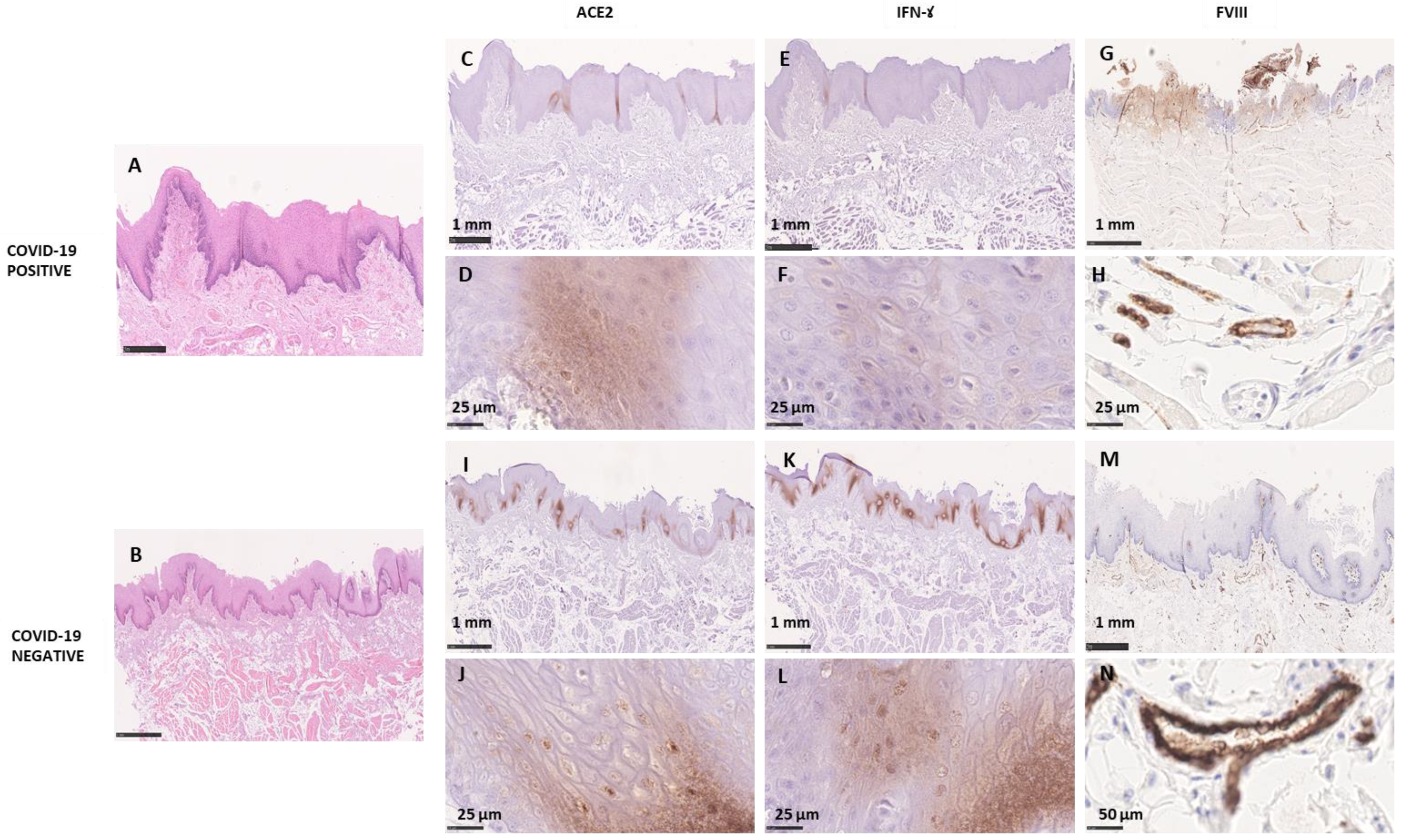
3.2.2. Morphological Analysis
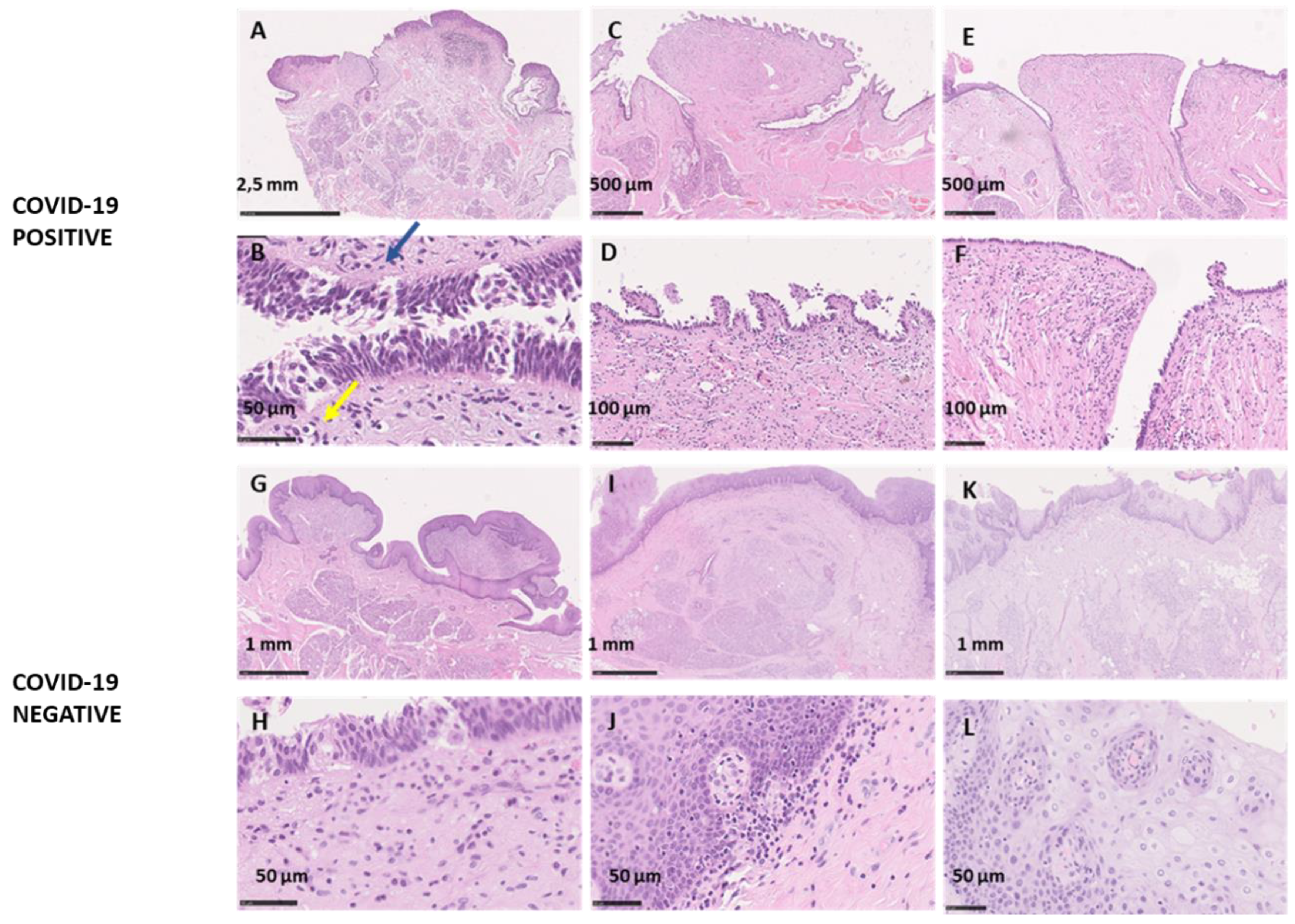
Macroscopic Analysis
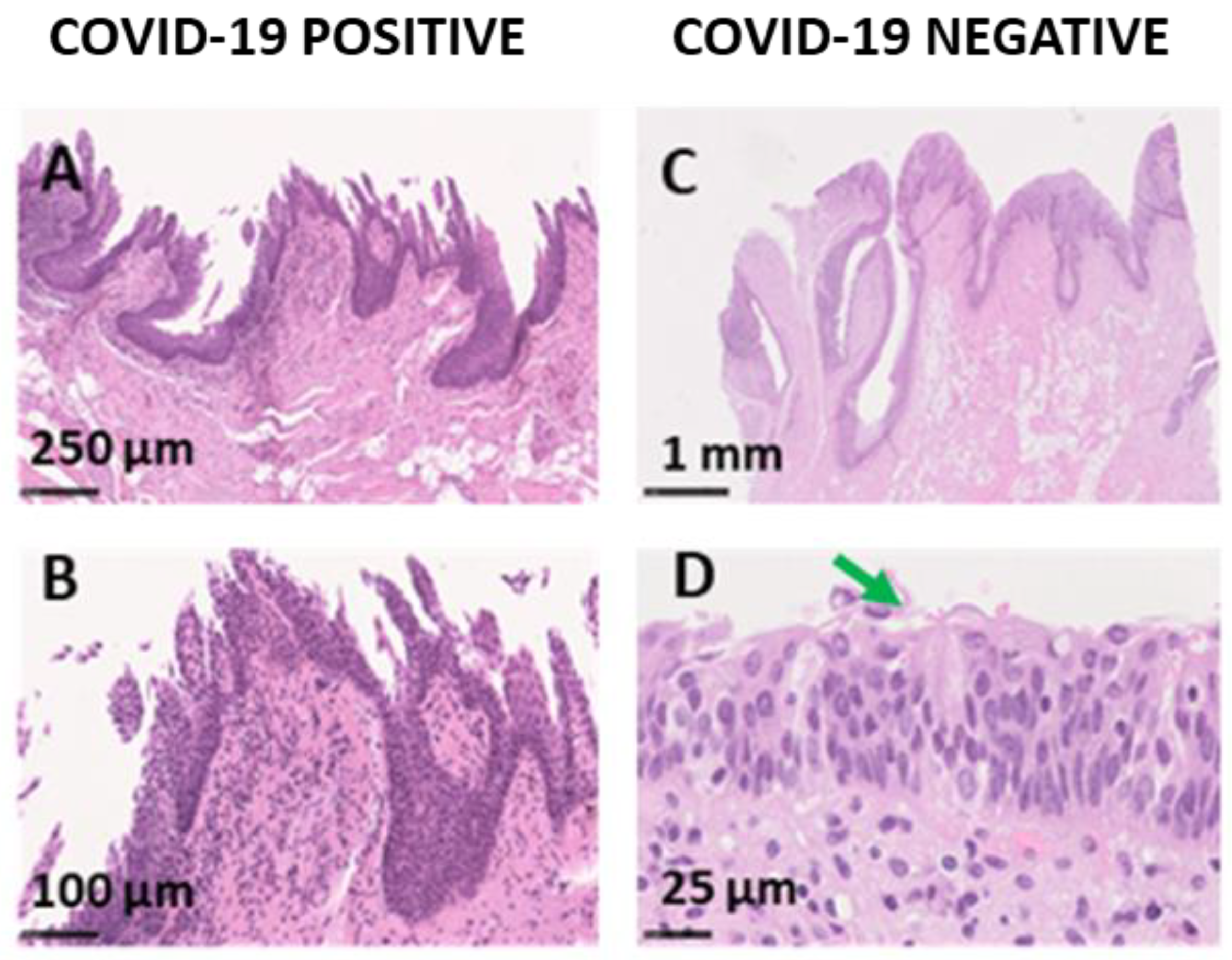
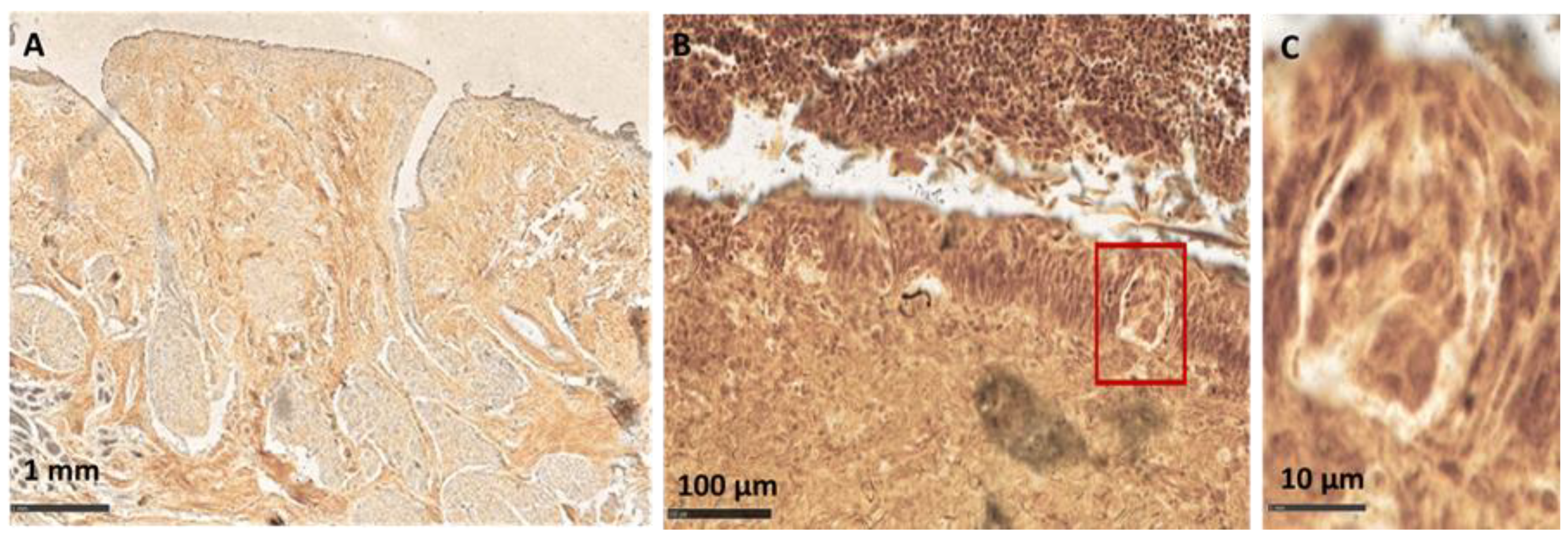
Microscopic Analysis
3.3. COVID-19−
3.3.1. Molecular Analysis
3.3.2. Morphological Analysis
Macroscopic Analysis
Microscopic Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bax, F.; Tascini, C.; Valente, M.; Marini, A.; Surcinelli, A.; Pellitteri, G.; De Carlo, C.; Gerussi, V.; Gigli, G.L. Hyposmia and Dysgeusia in COVID-19. Neurol. Clin. Pract. 2020, 11, e92–e96. [Google Scholar] [CrossRef] [PubMed]
- Mutiawati, E.; Fahriani, M.; Mamada, S.S.; Fajar, J.K.; Frediansyah, A.; Maliga, H.A.; Ilmawan, M.; Bin Emran, T.; Ophinni, Y.; Ichsan, I.; et al. Anosmia and dysgeusia in SARS-CoV-2 infection: Incidence and effects on COVID-19 severity and mortality, and the possible pathobiology mechanisms—A systematic review and meta-analysis. F1000Research 2021, 10, 40. [Google Scholar] [CrossRef]
- Nouchi, A.; Chastang, J.; Miyara, M.; Lejeune, J.; Soares, A.; Ibanez, G.; Saadoun, D.; Morélot-Panzini, C.; Similowski, T.; Amoura, Z.; et al. Prevalence of hyposmia and hypogeusia in 390 COVID-19 hospitalized patients and outpatients: A cross-sectional study. Eur. J. Clin. Microbiol. 2021, 40, 691–697. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Zhao, S.; Yu, B.; Chen, Y.-M.; Wang, W.; Song, Z.-G.; Hu, Y.; Tao, Z.-W.; Tian, J.-H.; Pei, Y.-Y.; et al. A new coronavirus associated with human respiratory disease in China. Nature 2020, 579, 265–269. [Google Scholar] [CrossRef] [Green Version]
- Zahra, S.A.; Iddawela, S.; Pillai, K.; Choudhury, R.Y.; Harky, A. Can symptoms of anosmia and dysgeusia be diagnostic for COVID-19? Brain Behav. 2020, 10, 10. [Google Scholar] [CrossRef]
- Afshar, Z.M.; Barary, M.; Ebrahimpour, S.; Janbakhsh, A.; Afsharian, M.; Hasanpour, A.; Babazadeh, A. Pathophysiology and Management of Tongue Involvement in COVID-19 Patients. Indian J. Otolaryngol. Head Neck Surg. 2022, 1–4. [Google Scholar] [CrossRef]
- Chi, A.C.; Neville, B.W.; Krayer, J.W.; Gonsalves, W.C. Oral Manifestations of Systemic Disease. Am. Fam. Physician 2010, 82, 1381–1388. [Google Scholar] [CrossRef]
- Stone, L.M.; Tan, S.-S.; Tam, P.P.L.; Finger, T. Analysis of Cell Lineage Relationships in Taste Buds. J. Neurosci. 2002, 22, 4522–4529. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T. Cellular Mechanisms in Taste Buds. Bull. Tokyo Dent. Coll. 2007, 48, 151–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okada, Y.; Yoshimura, K.; Toya, S.; Tsuchimochi, M. Pathogenesis of taste impairment and salivary dysfunction in COVID-19 patients. Jpn. Dent. Sci. Rev. 2021, 57, 111–122. [Google Scholar] [CrossRef]
- Vaira, L.A.; Salzano, G.; Fois, A.G.; Piombino, P.; De Riu, G. Potential pathogenesis of ageusia and anosmia in COVID-19 patients. Int. Forum Allergy Rhinol. 2020, 10, 1103–1104. [Google Scholar] [CrossRef] [PubMed]
- Kirschenbaum, D.; Imbach, L.L.; Ulrich, S.; Rushing, E.J.; Keller, E.; Reimann, R.R.; Frauenknecht, K.; Lichtblau, M.; Witt, M.; Hummel, T.; et al. Inflammatory olfactory neuropathy in two patients with COVID-19. Lancet 2020, 396, 166. [Google Scholar] [CrossRef]
- Vaira, L.A.; Hopkins, C.; Sandison, A.; Manca, A.; Machouchas, N.; Turilli, D.; Lechien, J.R.; Barillari, M.R.; Salzano, G.; Cossu, A.; et al. Olfactory epithelium histopathological findings in long-term coronavirus disease 2019 related anosmia. J. Laryngol. Otol. 2020, 134, 1123–1127. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.; Yoo, S.-J.; Clijsters, M.; Backaert, W.; Vanstapel, A.; Speleman, K.; Lietaer, C.; Choi, S.; Hether, T.D.; Marcelis, L.; et al. Visualizing in deceased COVID-19 patients how SARS-CoV-2 attacks the respiratory and olfactory mucosae but spares the olfactory bulb. Cell 2021, 184, 5932–5949.e15. [Google Scholar] [CrossRef]
- Maiorano, E.; Calastri, A.; Robotti, C.; Cassaniti, I.; Baldanti, F.; Zuccaro, V.; Stellin, E.; Ferretti, V.V.; Klersy, C.; Benazzo, M. Clinical, virological and immunological evolution of the olfactory and gustatory dysfunction in COVID-19. Am. J. Otolaryngol. 2021, 43, 103170. [Google Scholar] [CrossRef]
- Hannum, M.E.; Koch, R.J.; Ramirez, V.A.; Marks, S.S.; Toskala, A.K.; Herriman, R.D.; Lin, C.; Joseph, P.V.; Reed, D.R. Taste loss as a distinct symptom of COVID-19: A systematic review and meta-analysis. Chem. Senses 2022, 47. [Google Scholar] [CrossRef]
- Vaira, L.A.; Lechien, J.R.; Salzano, G.; Salzano, F.A.; Maglitto, F.; Saussez, S.; De Riu, G. Gustatory Dysfunction: A Highly Specific and Smell-Independent Symptom of COVID-19. Indian J. Otolaryngol. Head Neck Surg. 2020, 30, 1–3. [Google Scholar] [CrossRef]
- Borghi, E.; Massa, V.; Carmagnola, D.; Dellavia, C.; Parodi, C.; Ottaviano, E.; Sangiorgio, A.; Barcellini, L.; Gambacorta, G.; Forlanini, F.; et al. Saliva Sampling for Chasing SARS-CoV-2: A Game-Changing Strategy. Pharmacol. Res. 2021, 165, 105380. [Google Scholar] [CrossRef] [PubMed]
- Carmagnola, D.; Pellegrini, G.; Canciani, E.; Henin, D.; Perrotta, M.; Forlanini, F.; Barcellini, L.; Dellavia, C. Saliva Molecular Testing for SARS-CoV-2 Surveillance in Two Italian Primary Schools. Children 2021, 8, 544. [Google Scholar] [CrossRef] [PubMed]
- Varga, Z.; Flammer, A.J.; Steiger, P.; Haberecker, M.; Andermatt, R.; Zinkernagel, A.S.; Mehra, M.R.; Schuepbach, R.A.; Ruschitzka, F.; Moch, H. Endothelial cell infection and endotheliitis in COVID-19. Lancet 2020, 395, 1417–1418. [Google Scholar] [CrossRef]
- Sato, T.; Ueha, R.; Goto, T.; Yamauchi, A.; Kondo, K.; Yamasoba, T. Expression of ACE2 and TMPRSS2 Proteins in the Upper and Lower Aerodigestive Tracts of Rats: Implications on COVID 19 Infections. Laryngoscope 2021, 131, E932–E939. [Google Scholar] [CrossRef] [PubMed]
- Doyle, M.E.; Appleton, A.; Liu, Q.-R.; Yao, Q.; Mazucanti, C.H.; Egan, J.M. Human Type II Taste Cells Express Angiotensin-Converting Enzyme 2 and Are Infected by Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2). Am. J. Pathol. 2021, 191, 1511–1519. [Google Scholar] [CrossRef]
- Costela-Ruiz, V.J.; Illescas-Montes, R.; Puerta-Puerta, J.M.; Ruiz, C.; Melguizo-Rodríguez, L. SARS-CoV-2 infection: The role of cytokines in COVID-19 disease. Cytokine Growth Factor Rev. 2020, 54, 62–75. [Google Scholar] [CrossRef] [PubMed]
- Karki, R.; Sharma, B.R.; Tuladhar, S.; Williams, E.P.; Zalduondo, L.; Samir, P.; Zheng, M.; Sundaram, B.; Banoth, B.; Malireddi, R.K.S.; et al. Synergism of TNF-α and IFN-γ Triggers Inflammatory Cell Death, Tissue Damage, and Mortality in SARS-CoV-2 Infection and Cytokine Shock Syndromes. Cell 2021, 184, 149–168.e17. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, H.; Nakagawa, K.; Tago, M.; Nakamura, N.; Shiraishi, K.; Eda, M.; Nakata, H.; Ms, N.N.; Yokoyama, R.; Bs, M.O.; et al. Taste dysfunction in patients receiving radiotherapy. Head Neck 2006, 28, 508–516. [Google Scholar] [CrossRef]
- Henkin, R.I.; Larson, A.L.; Powell, R.D. Hypogeusia, Dysgeusia, Hyposmia, and Dysosmia following Influenza-Like Infection. Ann. Otol. Rhinol. Laryngol. 1975, 84, 672–682. [Google Scholar] [CrossRef]
- Gu, S.X.; Tyagi, T.; Jain, K.; Gu, V.W.; Lee, S.H.; Hwa, J.M.; Kwan, J.M.; Krause, D.S.; Lee, A.I.; Halene, S.; et al. Thrombocytopathy and Endotheliopathy: Crucial Contributors to COVID-19 Thromboinflammation. Nat. Rev. Cardiol. 2021, 18, 194–209. [Google Scholar] [CrossRef]
- Arey, L.B.; Tremaine, M.J.; Monzingo, F.L. The numerical and topographical relations of taste buds to human circumv allate papillae throughout the life span. Anat. Rec. 1935, 64, 9–25. [Google Scholar] [CrossRef]
- Frati, E.; Bianchi, S.; Amendola, A.; Colzani, D.; Petrelli, F.; Zehender, G.; Tanzi, E. Genetic characterization of variants of HPV-16, HPV-18 and HPV-52 circulating in Italy among general and high-risk populations. Mol. Med. Rep. 2019, 21, 894–902. [Google Scholar] [CrossRef]
- Croci, G.; Vaira, V.; Trabattoni, D.; Biasin, M.; Valenti, L.; Baselli, G.; Barberis, M.; Rocco, E.G.; Gregato, G.; Scandroglio, M.; et al. Emergency Lung Transplantation after COVID-19: Immunopathological Insights on Two Affected Patients. Cells 2021, 10, 611. [Google Scholar] [CrossRef]
- Dellavia, C.; Canullo, L.; Allievi, C.; Lang, N.P.; Pellegrini, G. Soft tissue surrounding switched platform implants: An immunohistochemical evaluation. Clin. Oral Implant. Res. 2013, 24, 63–70. [Google Scholar] [CrossRef]
- Guerini-Rocco, E.; Taormina, S.V.; Vacirca, D.; Ranghiero, A.; Rappa, A.; Fumagalli, C.; Maffini, F.; Rampinelli, C.; Galetta, D.; Tagliabue, M.; et al. SARS-CoV-2 detection in formalin-fixed paraffin-embedded tissue specimens from surgical resection of tongue squamous cell carcinoma. J. Clin. Pathol. 2020, 73, 754–757. [Google Scholar] [CrossRef] [PubMed]
- Bradley, B.T.; Maioli, H.; Johnston, R.; Chaudhry, I.; Fink, S.L.; Xu, H.; Najafian, B.; Deutsch, G.; Lacy, J.M.; Williams, T.; et al. Histopathology and ultrastructural findings of fatal COVID-19 infections in Washington State: A case series. Lancet 2020, 396, 320–332, Erratum in Lancet 2020, 396, 312. [Google Scholar] [CrossRef]
- Zhu, N.; Zhang, D.; Wang, W.; Li, X.; Yang, B.; Song, J.; Zhao, X.; Huang, B.; Shi, W.; Lu, R.; et al. A Novel Coronavirus from Patients with Pneumonia in China, 2019. N. Engl. J. Med. 2020, 382, 727–733. [Google Scholar] [CrossRef] [PubMed]
- Borczuk, A.C.; Salvatore, S.P.; Seshan, S.V.; Patel, S.S.; Bussel, J.B.; Mostyka, M.; Elsoukkary, S.; He, B.; DEL Vecchio, C.; Fortarezza, F.; et al. COVID-19 pulmonary pathology: A multi-institutional autopsy cohort from Italy and New York City. Mod. Pathol. 2020, 33, 2156–2168. [Google Scholar] [CrossRef] [PubMed]
- Witt, M. Anatomy and development of the human taste system. In Neurogenetics, Part II; Elsevier BV: Amsterdam, The Netherlands, 2019; Volume 164, pp. 147–171. [Google Scholar]
- Meinhardt, J.; Radke, J.; Dittmayer, C.; Franz, J.; Thomas, C.; Mothes, R.; Laue, M.; Schneider, J.; Brünink, S.; Greuel, S.; et al. Olfactory transmucosal SARS-CoV-2 invasion as a port of central nervous system entry in individuals with COVID-19. Nat. Neurosci. 2021, 24, 168–175. [Google Scholar] [CrossRef] [PubMed]
- Sakaguchi, W.; Kubota, N.; Shimizu, T.; Saruta, J.; Fuchida, S.; Kawata, A.; Yamamoto, Y.; Sugimoto, M.; Yakeishi, M.; Tsukinoki, K. Existence of SARS-CoV-2 Entry Molecules in the Oral Cavity. Int. J. Mol. Sci. 2020, 21, 6000. [Google Scholar] [CrossRef] [PubMed]
- Sawa, Y.; Ibaragi, S.; Okui, T.; Yamashita, J.; Ikebe, T.; Harada, H. Expression of SARS-CoV-2 entry factors in human oral tissue. J. Anat. 2021, 238, 1341–1354. [Google Scholar] [CrossRef]
- Haga, S.; Yamamoto, N.; Nakai-Murakami, C.; Osawa, Y.; Tokunaga, K.; Sata, T.; Sasazuki, T.; Ishizaka, Y. Modulation of TNF- -converting enzyme by the spike protein of SARS-CoV and ACE2 induces TNF- production and facilitates viral entry. Proc. Natl. Acad. Sci. USA 2008, 105, 7809–7814. [Google Scholar] [CrossRef] [Green Version]
- Glowacka, I.; Bertram, S.; Herzog, P.; Pfefferle, S.; Steffen, I.; Muench, M.O.; Simmons, G.; Hofmann, H.; Kuri, T.; Weber, F.; et al. Differential Downregulation of ACE2 by the Spike Proteins of Severe Acute Respiratory Syndrome Coronavirus and Human Coronavirus NL63. J. Virol. 2010, 84, 1198–1205. [Google Scholar] [CrossRef] [Green Version]
- Kuba, K.; Imai, Y.; Rao, S.; Gao, H.; Guo, F.; Guan, B.; Huan, Y.; Yang, P.; Zhang, Y.; Deng, W.; et al. A crucial role of angiotensin converting enzyme 2 (ACE2) in SARS coronavirus–induced lung injury. Nat. Med. 2005, 11, 875–879. [Google Scholar] [CrossRef] [PubMed]
- Oudit, G.Y.; Kassiri, Z.; Jiang, C.; Liu, P.P.; Poutanen, S.; Penninger, J.; Butany, J. SARS-coronavirus modulation of myocardial ACE2 expression and inflammation in patients with SARS. Eur. J. Clin. Investig. 2009, 39, 618–625. [Google Scholar] [CrossRef]
- Ziegler, C.G.K.; Allon, S.J.; Nyquist, S.K.; Mbano, I.M.; Miao, V.N.; Tzouanas, C.N.; Cao, Y.; Yousif, A.S.; Bals, J.; Hauser, B.M.; et al. SARS-CoV-2 Receptor ACE2 Is an Interferon-Stimulated Gene in Human Airway Epithelial Cells and Is Detected in Specific Cell Subsets across Tissues. Cell 2020, 181, 1016–1035.e19. [Google Scholar] [CrossRef]
- Heuberger, J.; Trimpert, J.; Vladimirova, D.; Goosmann, C.; Lin, M.; Schmuck, R.; Mollenkopf, H.; Brinkmann, V.; Tacke, F.; Osterrieder, N.; et al. Epithelial response to IFN-γ promotes SARS-CoV-2 infection. EMBO Mol. Med. 2021, 13, e13191. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Zhou, M.; Brand, J.; Huang, L. Inflammation Activates the Interferon Signaling Pathways in Taste Bud Cells. J. Neurosci. 2007, 27, 10703–10713. [Google Scholar] [CrossRef] [PubMed]
- Rapkiewicz, A.V.; Mai, X.; Carsons, S.E.; Pittaluga, S.; Kleiner, D.E.; Berger, J.S.; Thomas, S.; Adler, N.; Charytan, D.; Gasmi, B.; et al. Megakaryocytes and platelet-fibrin thrombi characterize multi-organ thrombosis at autopsy in COVID-19: A case series. EClinicalMedicine 2020, 24, 100434. [Google Scholar] [CrossRef] [PubMed]
- Wichmann, D.; Sperhake, J.P.; Lütgehetmann, M.; Steurer, S.; Edler, C.; Heinemann, A.; Heinrich, F.; Mushumba, H.; Kniep, I.; Schröder, A.S.; et al. Autopsy Findings and Venous Thromboembolism in Patients with COVID-19. Ann. Intern. Med. 2020, 173, 268–277. [Google Scholar] [CrossRef] [PubMed]
- Cipolloni, L.; Sessa, F.; Bertozzi, G.; Baldari, B.; Cantatore, S.; Testi, R.; D’Errico, S.; Di Mizio, G.; Asmundo, A.; Castorina, S.; et al. Preliminary Post-Mortem COVID-19 Evidence of Endothelial Injury and Factor VIII Hyperexpression. Diagnostics 2020, 10, 575. [Google Scholar] [CrossRef]
- Jacquemin, M.; Neyrinck, A.; Hermanns, M.I.; Lavend’Homme, R.; Rega, F.; Saint-Remy, J.-M.; Peerlinck, K.; Van Raemdonck, D.; Kirkpatrick, C.J. FVIII production by human lung microvascular endothelial cells. Blood 2006, 108, 515–517. [Google Scholar] [CrossRef]
- Brenner, E. Human body preservation—Old and new techniques. J. Anat. 2014, 224, 316–344. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.K.; Jo, D.H. Current and potential use of fresh frozen cadaver in surgical training and anatomical education. Anat. Sci. Educ. 2021. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subject ID | Age (Years) | Gender | Death-Autopsy Time Lapse (Days) | Pre-Death State | Cause of Death |
|---|---|---|---|---|---|
| COVID-19 positive | |||||
| 1 | 41 | M | 9 | Unknown | Traffic accident |
| 2 | 65 | M | 17 | Influenza-like symptoms | Bilateral pneumonia |
| 3 | 67 | M | 7 | Unknown | Cardiac arrest |
| 4 | 73 | M | 7 | Previous hospitalization for COVID-19 related respiratory symptoms | Cardiac arrest |
| 5 | 57 | M | 57 | Influenza-like symptoms | Pulmonary edema |
| 6 | 47 | M | 10 | Influenza-like symptoms | Pulmonary edema |
| 7 | 25 | M | 11 | Influenza-like symptoms | Cardiac arrest |
| 8 | 47 | M | 16 | Influenza-like symptoms | Cardiac arrest |
| COVID-19 negative | |||||
| 9 | 68 | M | 6 | Influenza-like symptoms in HIV+ | Cardiac arrest |
| 10 | 50 | M | 8 | Influenza-like symptoms | Cardiac arrest |
| 11 | 85 | M | 5 | Unknown | Stroke |
| 12 | 77 | F | 30 | Unknown | Cardiac arrest |
| 13 | 51 | M | 8 | Influenza-like symptoms | Cardiac arrest |
| 14 | 72 | M | 11 | Unknown | Cardiac arrest |
| 15 | 63 | F | 3 | Unknown | Trauma |
| 16 | 78 | M | 7 | Unknown | Cardiac Arrest |
| Subject ID | Real-Time RT-PCR | VP (n) | VP Dimension (mm) |
|---|---|---|---|
| COVID-19 positive | |||
| 1 | Negative | / | / |
| 2 | Positive | 8 | 4–5 |
| 3 | Negative | 7 | 3 |
| 4 | Positive | 7 | 6 |
| 5 | Positive | 6 | 5 |
| 6 | Positive | 8 | 4 |
| 7 | Negative | 9 | 2–3 |
| 8 | Negative | 8 | 4–5 |
| COVID-19 negative | |||
| 9 | Negative | 11 | 2–6 |
| 10 | Negative | 6 | 3 |
| 11 | Negative | 7 | 4 |
| 12 | Negative | 9 | 3–4 |
| 13 | Negative | 8 | 4 |
| 14 | Negative | 7 | 4 |
| 15 | Negative | 9 | 2–3 |
| 16 | Negative | 8 | 4–5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Henin, D.; Pellegrini, G.; Carmagnola, D.; Lanza Attisano, G.C.; Lopez, G.; Ferrero, S.; Amendola, A.; De Angelis, D.; Tanzi, E.; Dellavia, C. Morphological and Immunopathological Aspects of Lingual Tissues in COVID-19. Cells 2022, 11, 1248. https://doi.org/10.3390/cells11071248
Henin D, Pellegrini G, Carmagnola D, Lanza Attisano GC, Lopez G, Ferrero S, Amendola A, De Angelis D, Tanzi E, Dellavia C. Morphological and Immunopathological Aspects of Lingual Tissues in COVID-19. Cells. 2022; 11(7):1248. https://doi.org/10.3390/cells11071248
Chicago/Turabian StyleHenin, Dolaji, Gaia Pellegrini, Daniela Carmagnola, Giuseppe Carlo Lanza Attisano, Gianluca Lopez, Stefano Ferrero, Antonella Amendola, Danilo De Angelis, Elisabetta Tanzi, and Claudia Dellavia. 2022. "Morphological and Immunopathological Aspects of Lingual Tissues in COVID-19" Cells 11, no. 7: 1248. https://doi.org/10.3390/cells11071248
APA StyleHenin, D., Pellegrini, G., Carmagnola, D., Lanza Attisano, G. C., Lopez, G., Ferrero, S., Amendola, A., De Angelis, D., Tanzi, E., & Dellavia, C. (2022). Morphological and Immunopathological Aspects of Lingual Tissues in COVID-19. Cells, 11(7), 1248. https://doi.org/10.3390/cells11071248