
Revisiting Two Decades of Research Focused on Targeting APE1 for Cancer Therapy: The Pros and Cons




Abstract
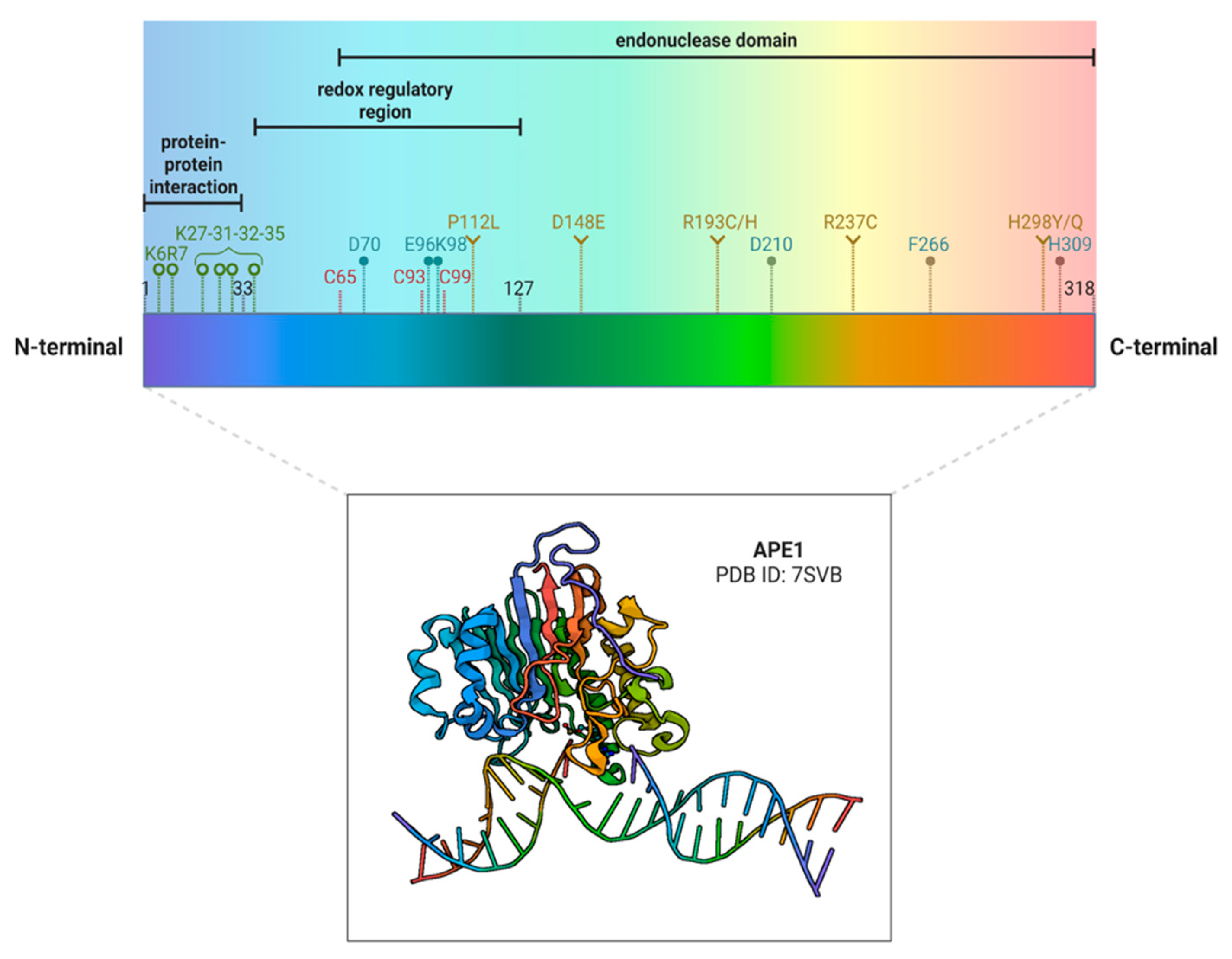
:1. A Brief Introduction to APE1 Biology and Different Functions
2. APE1 and Cancer: A Focus on Polymorphisms and Tissue Expression
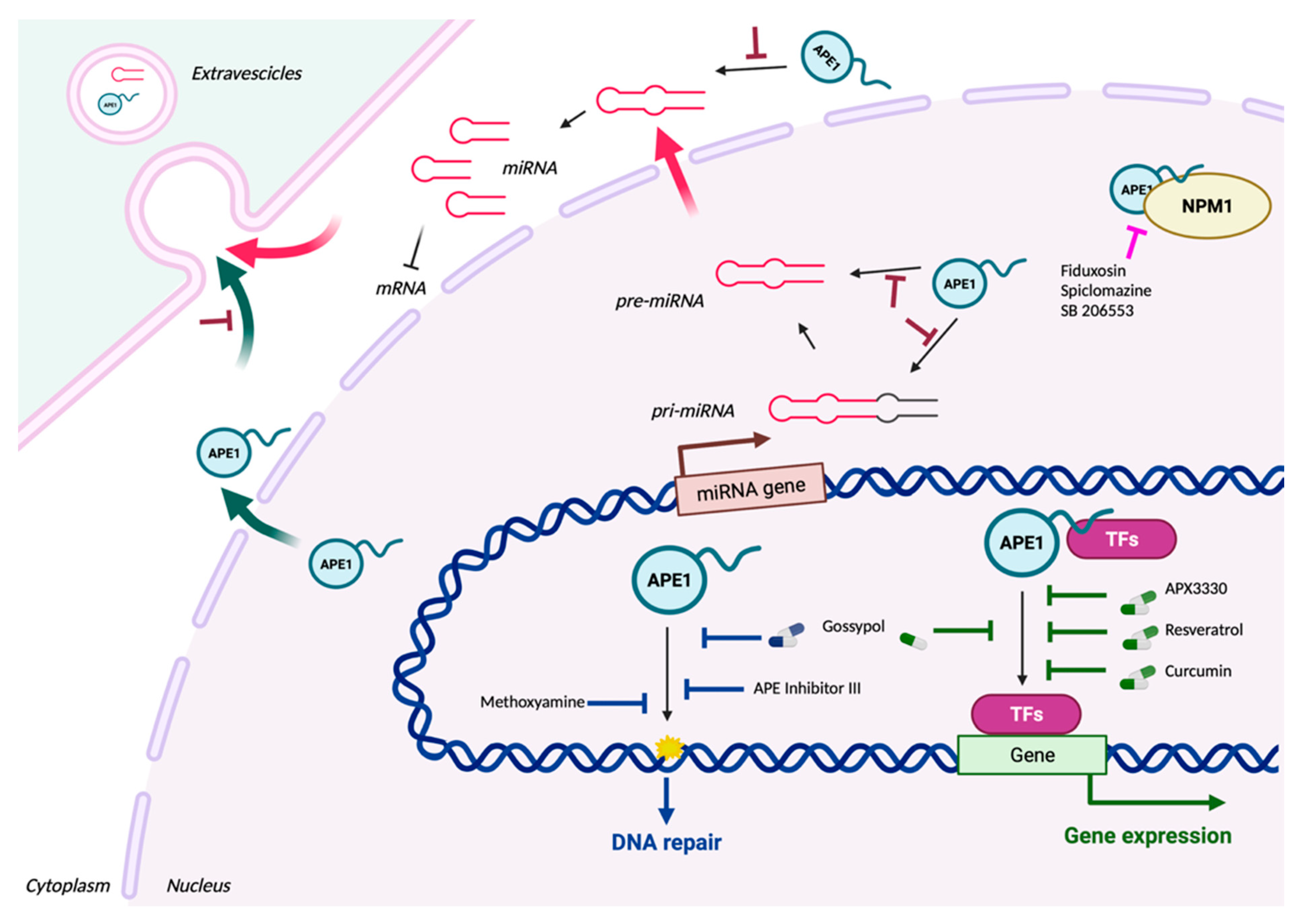
3. APE1 as a Still Promising Therapeutic Target after 20 Years of Research
3.1. Targeting the APE1 Endonuclease Activity
3.2. Targeting the APE1 Redox Activity
3.3. Targeting Both the APE1 Endonuclease and Redox Activities
4. Future Perspectives from Targeting the Non-Canonical Roles of APE1 in miRNA Processing
5. Secreted APE1 as a Novel Prognostic Non-Invasive Biomarker of Cancer Development
6. Conclusive Remarks and Future Perspectives

{kind=link}
{kind=link}
| APE1 Function- Inhibited | Name | IUPAC Name | PubChem CID | Molecular Formula | MW (g/mol) | Structure | Refs |
|---|---|---|---|---|---|---|---|
| Endonuclease | Methoxyamine | O-methylhydroxylamine | 4113 | CH5NO | 47.057 |  | [139,140,141] |
| Lucanthone | 1-[2-(diethylamino)ethylamino]-4-methylthioxanthen-9-one | 10180 | C20H24N2OS | 340.5 |  | [143,144,145] | |
| CRT0044867 | 7-Nitroindole-2-carboxylic acid | 81409 | C9H6N2O4 | 206.15 |  | [23,146,147,148] | |
| Myricetin | 3,5,7-trihydroxy-2-(3,4,5-trihydroxyphenyl)chromen-4-one | 5281672 | C15H10O8 | 318.23 |  | [147] | |
| AR03 | 2,4,9-Trimethylbenzo[b][1,8]naphthyridin-5-amine | 698490 | C15H15N3 | 237.30 |  | [151] | |
| APE Inhibitor III | N-[3-(1,3-benzothiazol-2-yl)-6-isopropyl-4,5,6,7-tetrahydrothieno[2,3-c]pyridin-2-yl]acetamide | 3581333 | C19H21N3OS2 | 371.5 |  | [155,156] | |
| Redox | Curcumin | (1E,6E)-1,7-bis(4-hydroxy-3-methoxyphenyl)hepta-1,6-diene-3,5-dione | 969516 | C21H20O6 | 368.4 |  | [175,176] |
| Resveratrol | 5-[(E)-2-(4-hydroxyphenyl)ethenyl]benzene-1,3-diol | 445154 | C14H12O3 | 228.24 |  | [99,177] | |
| APX3330 | (2E)-2-[(4,5-dimethoxy-2-methyl-3,6-dioxocyclohexa-1,4-dien-1-yl)methylidene]undecanoic acid | 6439397 | C21H30O6 | 378.5 |  | [24,179,180,181,182,183,184,185,186,187,188,189] | |
| APX2009 | (2E)-N,N-diethyl-2-[(3-methoxy-1,4-dioxonaphthalen-2-yl)methylidene]pentanamide | 71618575 | C21H25NO4 | 355.4 |  | [179] | |
| Endonuclease Redox | Gossypol | 7-(8-formyl-1,6,7-trihydroxy-3-methyl-5-propan-2-ylnaphthalen-2-yl)-2,3,8-trihydroxy-6-methyl-4-propan-2-ylnaphthalene-1-carbaldehyde | 3503 | C30H30O8 | 518.6 |  | [192,193] |
| Protein-protein interaction | Fiduxosin | 5-[4-[(3aR,9bR)-9-methoxy-3,3a,4,9b-tetrahydro-1H-chromeno[3,4-c]pyrrol-2-yl]butyl]-12-phenyl-8-thia-3,5,10,13-tetrazatricyclo[7.4.0.02,7]trideca-1(13),2(7),9,11-tetraene-4,6-dione | 172307 | C30H29N5O4S | 555.6 |  | [49,118,202] |
| Spiclomazine | 8-[3-(2-chlorophenothiazin-10-yl)propyl]-1-thia-4,8-diazaspiro[4.5]decan-3-one | 65714 | C22H24ClN3OS2 | 446.0 |  | [49,118,202] | |
| SB 206553 | 1-methyl-N-pyridin-3-yl-6,7-dihydropyrrolo[2,3-f]indole-5-carboxamide | 5163 | C17H16N4O | 292.33 |  | [49,118,202] |
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Antoniali, G.; Malfatti, M.C.; Tell, G. Unveiling the Non-Repair Face of the Base Excision Repair Pathway in RNA Processing: A Missing Link between DNA Repair and Gene Expression? DNA Repair 2017, 56, 65–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malfatti, M.C.; Antoniali, G.; Codrich, M.; Burra, S.; Mangiapane, G.; Dalla, E.; Tell, G. New Perspectives in Cancer Biology from a Study of Canonical and Non-Canonical Functions of Base Excision Repair Proteins with a Focus on Early Steps. Mutagenesis 2020, 35, 129–149. [Google Scholar] [CrossRef] [PubMed]
- Malfatti, M.C.; Antoniali, G.; Codrich, M.; Tell, G. Coping with RNA Damage with a Focus on APE1, a BER Enzyme at the Crossroad between DNA Damage Repair and RNA Processing/Decay. DNA Repair 2021, 104, 103133. [Google Scholar] [CrossRef] [PubMed]
- Allinson, S.L.; Sleeth, K.M.; Matthewman, G.E.; Dianov, G.L. Orchestration of Base Excision Repair by Controlling the Rates of Enzymatic Activities. DNA Repair 2004, 3, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Raj, J.; Li, J.; Ha, A.; Hossain, M.A.; Richardson, C.; Mukherjee, P.; Yan, S. APE1 Senses DNA Single-Strand Breaks for Repair and Signaling. Nucleic Acids Res. 2020, 48, 1925–1940. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.; Li, J.; Zhao, H.; McMahon, A.; McGhee, K.; Yan, S. APE1 Recruits ATRIP to SsDNA in an RPA-Dependent and -Independent Manner to Promote the ATR DNA Damage Response. eLife 2023, 12, e82324. [Google Scholar] [CrossRef]
- Hurley, L.H.; Wheelhouse, R.T.; Sun, D.; Kerwin, S.M.; Salazar, M.; Fedoroff, O.Y.; Han, F.X.; Han, H.; Izbicka, E.; Von Hoff, D.D. G-Quadruplexes as Targets for Drug Design. Pharmacol. Ther. 2000, 85, 141–158. [Google Scholar] [CrossRef]
- Fleming, A.M.; Burrows, C.J. G-Quadruplex Folds of the Human Telomere Sequence Alter the Site Reactivity and Reaction Pathway of Guanine Oxidation Compared to Duplex DNA. Chem. Res. Toxicol. 2013, 26, 593–607. [Google Scholar] [CrossRef] [Green Version]
- Fleming, A.M.; Zhu, J.; Ding, Y.; Visser, J.A.; Zhu, J.; Burrows, C.J. Human DNA Repair Genes Possess Potential G-Quadruplex Sequences in Their Promoters and 5′-Untranslated Regions. Biochemistry 2018, 57, 991–1002. [Google Scholar] [CrossRef]
- Fleming, A.M.; Burrows, C.J. 8-Oxo-7,8-Dihydroguanine, Friend and Foe: Epigenetic-like Regulator versus Initiator of Mutagenesis. DNA Repair 2017, 56, 75–83. [Google Scholar] [CrossRef]
- Fleming, A.M.; Zhu, J.; Ding, Y.; Burrows, C.J. 8-Oxo-7,8-dihydroguanine in the Context of a Gene Promoter G-Quadruplex Is an On–Off Switch for Transcription. ACS Chem. Biol. 2017, 12, 2417–2426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burra, S.; Marasco, D.; Malfatti, M.C.; Antoniali, G.; Virgilio, A.; Esposito, V.; Demple, B.; Galeone, A.; Tell, G. Human AP-Endonuclease (Ape1) Activity on Telomeric G4 Structures Is Modulated by Acetylatable Lysine Residues in the N-Terminal Sequence. DNA Repair 2019, 73, 129–143. [Google Scholar] [CrossRef]
- Roychoudhury, S.; Pramanik, S.; Harris, H.L.; Tarpley, M.; Sarkar, A.; Spagnol, G.; Sorgen, P.L.; Chowdhury, D.; Band, V.; Klinkebiel, D.; et al. Endogenous Oxidized DNA Bases and APE1 Regulate the Formation of G-Quadruplex Structures in the Genome. Proc. Natl. Acad. Sci. USA 2020, 117, 11409–11420. [Google Scholar] [CrossRef] [PubMed]
- Antoniali, G.; Serra, F.; Lirussi, L.; Tanaka, M.; D’Ambrosio, C.; Zhang, S.; Radovic, S.; Dalla, E.; Ciani, Y.; Scaloni, A.; et al. Mammalian APE1 Controls MiRNA Processing and Its Interactome Is Linked to Cancer RNA Metabolism. Nat. Commun. 2017, 8, 797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barnes, T.; Kim, W.-C.; Mantha, A.K.; Kim, S.-E.; Izumi, T.; Mitra, S.; Lee, C.H. Identification of Apurinic/Apyrimidinic Endonuclease 1 (APE1) as the Endoribonuclease That Cleaves c-Myc MRNA. Nucleic Acids Res. 2009, 37, 3946–3958. [Google Scholar] [CrossRef] [Green Version]
- Chohan, M.; Mackedenski, S.; Li, W.-M.; Lee, C.H. Human Apurinic/Apyrimidinic Endonuclease 1 (APE1) Has 3′ RNA Phosphatase and 3′ Exoribonuclease Activities. J. Mol. Biol. 2015, 427, 298–311. [Google Scholar] [CrossRef]
- Malfatti, M.C.; Balachander, S.; Antoniali, G.; Koh, K.D.; Saint-Pierre, C.; Gasparutto, D.; Chon, H.; Crouch, R.J.; Storici, F.; Tell, G. Abasic and Oxidized Ribonucleotides Embedded in DNA Are Processed by Human APE1 and Not by RNase H2. Nucleic Acids Res. 2017, 45, 11193–11212. [Google Scholar] [CrossRef] [Green Version]
- Malfatti, M.C.; Codrich, M.; Dalla, E.; D’Ambrosio, C.; Storici, F.; Scaloni, A.; Tell, G. AUF1 Recognizes 8-Oxo-Guanosine Embedded in DNA and Stimulates APE1 Endoribonuclease Activity. Antioxid. Redox Signal. 2023; ahead of print. [Google Scholar] [CrossRef]
- Tosolini, D.; Antoniali, G.; Dalla, E.; Tell, G. Role of Phase Partitioning in Coordinating DNA Damage Response: Focus on the Apurinic Apyrimidinic Endonuclease 1 Interactome. Biomol. Concepts 2020, 11, 209–220. [Google Scholar] [CrossRef]
- Li, J.; Zhao, H.; McMahon, A.; Yan, S. APE1 Assembles Biomolecular Condensates to Promote the ATR–Chk1 DNA Damage Response in Nucleolus. Nucleic Acids Res. 2022, 50, 10503–10525. [Google Scholar] [CrossRef]
- Dall’Agnese, G.; Dall’Agnese, A.; Banani, S.F.; Codrich, M.; Malfatti, M.C.; Antoniali, G.; Tell, G. Role of Condensates in Modulating DNA Repair Pathways and Its Implication for Chemoresistance. J. Biol. Chem. 2023, 299, 104800. [Google Scholar] [CrossRef] [PubMed]
- Mangiapane, G.; Parolini, I.; Conte, K.; Malfatti, M.C.; Corsi, J.; Sanchez, M.; Pietrantoni, A.; D’Agostino, V.G.; Tell, G. Enzymatically Active Apurinic/Apyrimidinic Endodeoxyribonuclease 1 Is Released by Mammalian Cells through Exosomes. J. Biol. Chem. 2021, 296, 100569. [Google Scholar] [CrossRef]
- Fishel, M.L.; Kelley, M.R. The DNA Base Excision Repair Protein Ape1/Ref-1 as a Therapeutic and Chemopreventive Target. Mol. Asp. Med. 2007, 28, 375–395. [Google Scholar] [CrossRef]
- Shah, F.; Logsdon, D.; Messmann, R.A.; Fehrenbacher, J.C.; Fishel, M.L.; Kelley, M.R. Exploiting the Ref-1-APE1 Node in Cancer Signaling and Other Diseases: From Bench to Clinic. NPJ Precis. Oncol. 2017, 1, 19. [Google Scholar] [CrossRef] [Green Version]
- Lirussi, L.; Antoniali, G.; D’Ambrosio, C.; Scaloni, A.; Nilsen, H.; Tell, G. APE1 Polymorphic Variants Cause Persistent Genomic Stress and Affect Cancer Cell Proliferation. Oncotarget 2016, 7, 26293–26306. [Google Scholar] [CrossRef] [PubMed]
- Wallace, S.S.; Murphy, D.L.; Sweasy, J.B. Base Excision Repair and Cancer. Cancer Lett. 2012, 327, 73–89. [Google Scholar] [CrossRef] [Green Version]
- Whitaker, A.M.; Stark, W.J.; Flynn, T.S.; Freudenthal, B.D. Molecular and Structural Characterization of Disease-Associated APE1 Polymorphisms. DNA Repair 2020, 91–92, 102867. [Google Scholar] [CrossRef]
- Kim, W.C.; Ma, C.; Li, W.-M.; Chohan, M.; Wilson, D.M., III; Lee, C.H. Altered Endoribonuclease Activity of Apurinic/Apyrimidinic Endonuclease 1 Variants Identified in the Human Population. PLoS ONE 2014, 9, e90837. [Google Scholar] [CrossRef] [PubMed]
- Wilson, D.M.; Kim, D.; Berquist, B.R.; Sigurdson, A.J. Variation in Base Excision Repair Capacity. Mutat. Res. 2011, 711, 100–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pieretti, M.; Khattar, N.H.; Smith, S.A. Common Polymorphisms and Somatic Mutations in Human Base Excision Repair Genes in Ovarian and Endometrial Cancers. Mutat. Res. 2001, 432, 53–59. [Google Scholar] [CrossRef]
- Illuzzi, J.L.; Harris, N.A.; Manvilla, B.A.; Kim, D.; Li, M.; Drohat, A.C.; Iii, D.M.W. Functional Assessment of Population and Tumor-Associated APE1 Protein Variants. PLoS ONE 2013, 8, e65922. [Google Scholar] [CrossRef] [PubMed]
- Abyzov, A.; Uzun, A.; Strauss, P.R.; Ilyin, V.A. An AP Endonuclease 1-DNA Polymerase Beta Complex: Theoretical Prediction of Interacting Surfaces. PLoS Comput. Biol. 2008, 4, e1000066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hinz, J.M.; Mao, P.; McNeill, D.R.; Wilson, D.M. Reduced Nuclease Activity of Apurinic/Apyrimidinic Endonuclease (APE1) Variants on Nucleosomes: Identification of Access Residues. J. Biol. Chem. 2015, 290, 21067–21075. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Aksoy, B.A.; Dogrusoz, U.; Dresdner, G.; Gross, B.; Sumer, S.O.; Sun, Y.; Jacobsen, A.; Sinha, R.; Larsson, E.; et al. Integrative Analysis of Complex Cancer Genomics and Clinical Profiles Using the CBioPortal. Sci. Signal. 2013, 6, pl1. [Google Scholar] [CrossRef] [Green Version]
- Cerami, E.; Gao, J.; Dogrusoz, U.; Gross, B.E.; Sumer, S.O.; Aksoy, B.A.; Jacobsen, A.; Byrne, C.J.; Heuer, M.L.; Larsson, E.; et al. The CBio Cancer Genomics Portal: An Open Platform for Exploring Multidimensional Cancer Genomics Data. Cancer Discov. 2012, 2, 401–404. [Google Scholar] [CrossRef] [Green Version]
- Reva, B.; Antipin, Y.; Sander, C. Predicting the Functional Impact of Protein Mutations: Application to Cancer Genomics. Nucleic Acids Res. 2011, 39, e118. [Google Scholar] [CrossRef] [Green Version]
- Reva, B.; Antipin, Y.; Sander, C. Determinants of Protein Function Revealed by Combinatorial Entropy Optimization. Genome Biol. 2007, 8, R232. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.; Shin, J.H.; Lee, Y.R.; Joo, H.K.; Song, K.H.; Na, Y.G.; Chang, S.J.; Lim, J.S.; Jeon, B.H. Urinary APE1/Ref-1: A Potential Bladder Cancer Biomarker. Dis. Markers 2016, 2016, 7276502. [Google Scholar] [CrossRef] [Green Version]
- Fishel, M.L.; Xia, H.; McGeown, J.; McIlwain, D.W.; Elbanna, M.; Craft, A.A.; Kaimakliotis, H.Z.; Sandusky, G.E.; Zhang, C.; Pili, R.; et al. Antitumor Activity and Mechanistic Characterization of APE1/Ref-1 Inhibitors in Bladder Cancer. Mol. Cancer Ther. 2019, 18, 1947–1960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, J.H.; Choi, S.; Lee, Y.R.; Park, M.S.; Na, Y.G.; Irani, K.; Lee, S.D.; Park, J.B.; Kim, J.M.; Lim, J.S.; et al. APE1/Ref-1 as a Serological Biomarker for the Detection of Bladder Cancer. Cancer Res. Treat. 2015, 47, 823–833. [Google Scholar] [CrossRef] [Green Version]
- Song, H.; Zeng, J.; Lele, S.; LaGrange, C.A.; Bhakat, K.K. APE1 and SSRP1 Is Overexpressed in Muscle Invasive Bladder Cancer and Associated with Poor Survival. Heliyon 2021, 7, e06756. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.-A.; Yang, B.; Tang, T.; Yang, Y.; Zhang, D.; Xiao, H.; Xu, J.; Wang, L.; Lin, L.; Jiang, J. Correlation of APE1 with VEGFA and CD163+ Macrophage Infiltration in Bladder Cancer and Their Prognostic Significance. Oncol. Lett. 2020, 20, 2881–2887. [Google Scholar] [CrossRef]
- Güllü Amuran, G.; Tinay, I.; Filinte, D.; Ilgin, C.; Peker Eyüboğlu, I.; Akkiprik, M. Urinary micro-RNA Expressions and Protein Concentrations May Differentiate Bladder Cancer Patients from Healthy Controls. Int. Urol. Nephrol. 2020, 52, 461–468. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Shukla, V.K.; Misra, P.K.; Raman, M.J. Dysregulated Expression and Subcellular Localization of Base Excision Repair (BER) Pathway Enzymes in Gallbladder Cancer. Int. J. Mol. Cell. Med. 2018, 7, 119–132. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Liu, C.; Lu, H.; Yin, M.; Shao, C.; Hu, X.; Wu, J.; Wang, Y. The Expression of APE1 in Triple-Negative Breast Cancer and Its Effect on Drug Sensitivity of Olaparib. Tumour Biol. 2017, 39, 1010428317713390. [Google Scholar] [CrossRef] [Green Version]
- Woo, J.; Park, H.; Sung, S.H.; Moon, B.-I.; Suh, H.; Lim, W. Prognostic Value of Human Apurinic/Apyrimidinic Endonuclease 1 (APE1) Expression in Breast Cancer. PLoS ONE 2014, 9, e99528. [Google Scholar] [CrossRef]
- Jian, D.; Li, X.-M.; Dai, N.; Liang, D.-D.; Zhang, G.; Mao, C.-Y.; Wang, D.; Song, G.-B.; Li, M.-X.; Luo, H. Inhibition of APE1 Expression Enhances the Antitumor Activity of Olaparib in Triple-Negative Breast Cancer. Evid.-Based Complement. Altern. Med. 2022, 2022, 6048017. [Google Scholar] [CrossRef]
- Poletto, M.; Di Loreto, C.; Marasco, D.; Poletto, E.; Puglisi, F.; Damante, G.; Tell, G. Acetylation on Critical Lysine Residues of Apurinic/Apyrimidinic Endonuclease 1 (APE1) in Triple Negative Breast Cancers. Biochem. Biophys. Res. Commun. 2012, 424, 34–39. [Google Scholar] [CrossRef]
- Malfatti, M.C.; Gerratana, L.; Dalla, E.; Isola, M.; Damante, G.; Di Loreto, C.; Puglisi, F.; Tell, G. APE1 and NPM1 Protect Cancer Cells from Platinum Compounds Cytotoxicity and Their Expression Pattern Has a Prognostic Value in TNBC. J. Exp. Clin. Cancer Res. 2019, 38, 309. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Fatah, T.M.A.; Perry, C.; Moseley, P.; Johnson, K.; Arora, A.; Chan, S.; Ellis, I.O.; Madhusudan, S. Clinicopathological Significance of Human Apurinic/Apyrimidinic Endonuclease 1 (APE1) Expression in Oestrogen-Receptor-Positive Breast Cancer. Breast Cancer Res. Treat. 2014, 143, 411–421. [Google Scholar] [CrossRef]
- Herring, C.J.; West, C.M.L.; Wilks, D.P.; Davidson, S.E.; Hunter, R.D.; Berry, P.; Forster, G.; MacKinnon, J.; Rafferty, J.A.; Elder, R.H.; et al. Levels of the DNA Repair Enzyme Human Apurinic/Apyrimidinic Endonuclease (APE1, APEX, Ref-1) Are Associated with the Intrinsic Radiosensitivity of Cervical Cancers. Br. J. Cancer 1998, 78, 1128–1133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schindl, M.; Oberhuber, G.; Pichlbauer, E.G.; Obermair, A.; Birner, P.; Kelley, M.R. DNA Repair-Redox Enzyme Apurinic Endonuclease in Cervical Cancer: Evaluation of Redox Control of HIF-1alpha and Prognostic Significance. Int. J. Oncol. 2001, 19, 799–802. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Wei, X.; Zhou, Z.-W.; Wang, S.-N.; Jin, H.; Chen, K.-J.; Luo, J.; Westover, K.D.; Wang, J.-M.; Wang, D.; et al. GADD45α Sensitizes Cervical Cancer Cells to Radiotherapy via Increasing Cytoplasmic APE1 Level. Cell Death Dis. 2018, 9, 524. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Zhou, Z.-W.; Duan, W.; Qian, C.-Y.; Wang, S.-N.; Deng, M.-S.; Zi, D.; Wang, J.-M.; Mao, C.-Y.; Song, G.; et al. Inhibiting the Redox Function of APE1 Suppresses Cervical Cancer Metastasis via Disengagement of ZEB1 from E-Cadherin in EMT. J. Exp. Clin. Cancer Res. 2021, 40, 220. [Google Scholar] [CrossRef] [PubMed]
- Bhakat, K.K.; Sengupta, S.; Adeniyi, V.F.; Roychoudhury, S.; Nath, S.; Bellot, L.J.; Feng, D.; Mantha, A.K.; Sinha, M.; Qiu, S.; et al. Regulation of Limited N-Terminal Proteolysis of APE1 in Tumor via Acetylation and Its Role in Cell Proliferation. Oncotarget 2016, 7, 22590–22604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kakolyris, S.; Kaklamanis, L.; Engels, K.; Turley, H.; Hickson, I.D.; Gatter, K.C.; Harris, A.L. Human Apurinic Endonuclease 1 Expression in a Colorectal Adenoma-Carcinoma Sequence. Cancer Res. 1997, 57, 1794–1797. [Google Scholar]
- Lou, D.; Zhu, L.; Ding, H.; Dai, H.-Y.; Zou, G.-M. Aberrant Expression of Redox Protein Ape1 in Colon Cancer Stem Cells. Oncol. Lett. 2014, 7, 1078–1082. [Google Scholar] [CrossRef] [Green Version]
- Codrich, M.; Comelli, M.; Malfatti, M.C.; Mio, C.; Ayyildiz, D.; Zhang, C.; Kelley, M.R.; Terrosu, G.; Pucillo, C.E.M.; Tell, G. Inhibition of APE1-Endonuclease Activity Affects Cell Metabolism in Colon Cancer Cells via a P53-Dependent Pathway. DNA Repair 2019, 82, 102675. [Google Scholar] [CrossRef]
- Kühl Svoboda Baldin, R.; Austrália Paredes Marcondes Ribas, C.; de Noronha, L.; Veloso da Silva-Camargo, C.C.; Santos Sotomaior, V.; Martins Sebastião, A.P.; Vasconcelos de Castilho, A.P.; Rodrigues Montemor Netto, M. Expression of Parkin, APC, APE1, and Bcl-XL in Colorectal Polyps. J. Histochem. Cytochem. 2021, 69, 437–449. [Google Scholar] [CrossRef]
- Noike, T.; Miwa, S.; Soeda, J.; Kobayashi, A.; Miyagawa, S. Increased Expression of Thioredoxin-1, Vascular Endothelial Growth Factor, and Redox Factor-1 Is Associated with Poor Prognosis in Patients with Liver Metastasis from Colorectal Cancer. Hum. Pathol. 2008, 39, 201–208. [Google Scholar] [CrossRef]
- Song, H.; Zeng, J.; Roychoudhury, S.; Biswas, P.; Mohapatra, B.; Ray, S.; Dowlatshahi, K.; Wang, J.; Band, V.; Talmon, G.; et al. Targeting Histone Chaperone FACT Complex Overcomes 5-Fluorouracil Resistance in Colon Cancer. Mol. Cancer Ther. 2020, 19, 258–269. [Google Scholar] [CrossRef] [Green Version]
- Huajun, W.; Ying, F.; Hongxing, Z.; Weifeng, S.; Pingyang, S.; Mingde, H.; Guoguang, L. Clinical Value of Combined Detection of Serum APE1-Aabs and CEACAM-1 in the Diagnosis of Colorectal Cancer. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 1286–1289. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Zhen, P.; Niu, X.; Dai, Y.; Wang, Y.; Zhou, M. APE1 Promotes Proliferation and Migration of Cutaneous Squamous Cell Carcinoma. J. Dermatol. Sci. 2020, 100, 67–74. [Google Scholar] [CrossRef]
- Ajucarmelprecilla, A.; Pandi, J.; Dhandapani, R.; Ramanathan, S.; Chinnappan, J.; Paramasivam, R.; Thangavelu, S.; Mohammed Ghilan, A.-K.; Aljohani, S.A.S.; Oyouni, A.A.A.; et al. In Silico Identification of Hub Genes as Observing Biomarkers for Gastric Cancer Metastasis. Evid.-Based Complement. Altern. Med. 2022, 2022, 6316158. [Google Scholar] [CrossRef] [PubMed]
- Manoel-Caetano, F.S.; Rossi, A.F.T.; Calvet de Morais, G.; Severino, F.E.; Silva, A.E. Upregulation of the APE1 and H2AX Genes and miRNAs Involved in DNA Damage Response and Repair in Gastric Cancer. Genes Dis. 2019, 6, 176–184. [Google Scholar] [CrossRef] [PubMed]
- Qing, Y.; Li, Q.; Ren, T.; Xia, W.; Peng, Y.; Liu, G.-L.; Luo, H.; Yang, Y.-X.; Dai, X.-Y.; Zhou, S.-F.; et al. Upregulation of PD-L1 and APE1 Is Associated with Tumorigenesis and Poor Prognosis of Gastric Cancer. Drug Des. Dev. Ther. 2015, 9, 901–909. [Google Scholar] [CrossRef] [Green Version]
- Wei, X.; Li, Y.-B.; Li, Y.; Lin, B.-C.; Shen, X.-M.; Cui, R.-L.; Gu, Y.-J.; Gao, M.; Li, Y.-G.; Zhang, S. Prediction of Lymph Node Metastases in Gastric Cancer by Serum APE1 Expression. J. Cancer 2017, 8, 1492–1497. [Google Scholar] [CrossRef]
- Bobola, M.S.; Blank, A.; Berger, M.S.; Stevens, B.A.; Silber, J.R. Apurinic/Apyrimidinic Endonuclease Activity Is Elevated in Human Adult Gliomas. Clin. Cancer Res. 2001, 7, 3510–3518. [Google Scholar]
- Scott, T.L.; Wicker, C.A.; Suganya, R.; Dhar, B.; Pittman, T.; Horbinski, C.; Izumi, T. Polyubiquitination of Apurinic/Apyrimidinic Endonuclease 1 by Parkin. Mol. Carcinog. 2017, 56, 325–336. [Google Scholar] [CrossRef] [Green Version]
- Naidu, M.D.; Mason, J.M.; Pica, R.V.; Fung, H.; Peña, L.A. Radiation Resistance in Glioma Cells Determined by DNA Damage Repair Activity of Ape1/Ref-1. J. Radiat. Res. 2010, 51, 393–404. [Google Scholar] [CrossRef] [Green Version]
- Hudson, A.L.; Parker, N.R.; Khong, P.; Parkinson, J.F.; Dwight, T.; Ikin, R.J.; Zhu, Y.; Chen, J.; Wheeler, H.R.; Howell, V.M. Glioblastoma Recurrence Correlates With Increased APE1 and Polarization Toward an Immuno-Suppressive Microenvironment. Front. Oncol. 2018, 8, 314. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Xiao, H.; Luo, Q.; Li, M.; Wei, S.; Zhu, X.; Xiao, H.; Chen, L. Low APE1/Ref-1 Expression Significantly Correlates with MGMT Promoter Methylation in Patients with High-Grade Gliomas. Int. J. Clin. Exp. Pathol. 2016, 9, 9562–9568. [Google Scholar]
- Perry, C.; Agarwal, D.; Abdel-Fatah, T.M.A.; Lourdusamy, A.; Grundy, R.; Auer, D.T.; Walker, D.; Lakhani, R.; Scott, I.S.; Chan, S.; et al. Dissecting DNA Repair in Adult High Grade Gliomas for Patient Stratification in the Post-Genomic Era. Oncotarget 2014, 5, 5764–5781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsia, K.-T.; Liu, C.-J.; Mar, K.; Lin, L.-H.; Lin, C.-S.; Cheng, M.-F.; Lee, H.-S.; Chiu, S.-Y. Impact of Apurinic/Apyrimidinic Endonuclease 1/Redox Factor-1 on Treatment Response and Survival in Oral Squamous Cell Carcinoma. Head Neck 2016, 38, 550–559. [Google Scholar] [CrossRef]
- Wicker, C.A.; Takiar, V.; Suganya, R.; Arnold, S.M.; Brill, Y.M.; Chen, L.; Horbinski, C.M.; Napier, D.; Valentino, J.; Kudrimoti, M.R.; et al. Evaluation of Antioxidant Network Proteins as Novel Prognostic Biomarkers for Head and Neck Cancer Patients. Oral. Oncol. 2020, 111, 104949. [Google Scholar] [CrossRef]
- Santana, T.; Sá, M.C.; de Moura Santos, E.; Galvão, H.C.; Coletta, R.D.; Freitas, R. de A. DNA Base Excision Repair Proteins APE-1 and XRCC-1 Are Overexpressed in Oral Tongue Squamous Cell Carcinoma. J. Oral. Pathol. Med. 2017, 46, 496–503. [Google Scholar] [CrossRef]
- Xie, J.; Li, Y.; Kong, J.; Li, C. Apurinic/Apyrimidinic Endonuclease 1/Redox Factor-1 Could Serve as a Potential Serological Biomarker for the Diagnosis and Prognosis of Oral Squamous Cell Carcinoma. J. Oral Maxillofac. Surg. 2019, 77, 859–866. [Google Scholar] [CrossRef]
- Wang, J.; Lun, L.; Jiang, X.; Wang, Y.; Li, X.; Du, G.; Wang, J. APE1 Facilitates PD-L1-Mediated Progression of Laryngeal and Hypopharyngeal Squamous Cell Carcinoma. Int. Immunopharmacol. 2021, 97, 107675. [Google Scholar] [CrossRef]
- Lee, J.W.; Jin, J.; Rha, K.-S.; Kim, Y.M. Expression Pattern of Apurinic/Apyrimidinic Endonuclease in Sinonasal Squamous Cell Carcinoma. Otolaryngol. Head Neck Surg. 2012, 147, 788–795. [Google Scholar] [CrossRef]
- Souza, L.R.; Fonseca-Silva, T.; Pereira, C.S.; Santos, E.P.; Lima, L.C.; Carvalho, H.A.; Gomez, R.S.; Guimarães, A.L.S.; De Paula, A.M.B. Immunohistochemical Analysis of P53, APE1, HMSH2 and ERCC1 Proteins in Actinic Cheilitis and Lip Squamous Cell Carcinoma. Histopathology 2011, 58, 352–360. [Google Scholar] [CrossRef]
- Di Maso, V.; Mediavilla, M.G.; Vascotto, C.; Lupo, F.; Baccarani, U.; Avellini, C.; Tell, G.; Tiribelli, C.; Crocè, L.S. Transcriptional Up-Regulation of APE1/Ref-1 in Hepatic Tumor: Role in Hepatocytes Resistance to Oxidative Stress and Apoptosis. PLoS ONE 2015, 10, e0143289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Z.; Zhu, Y.; Aminbuhe; Fan, Q.; Peng, J.; Zhang, N. Differential Expression of APE1 in Hepatocellular Carcinoma and the Effects on Proliferation and Apoptosis of Cancer Cells. BST 2018, 12, 456–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, X.; Zhao, H.; Yuan, H.; Chu, Y.; Zhu, X. High Nuclear Expression of APE1 Correlates with Unfavorable Prognosis and Promotes Tumor Growth in Hepatocellular Carcinoma. J. Mol. Histol. 2021, 52, 219–231. [Google Scholar] [CrossRef] [PubMed]
- Pascut, D.; Sukowati, C.H.C.; Antoniali, G.; Mangiapane, G.; Burra, S.; Mascaretti, L.G.; Buonocore, M.R.; Crocè, L.S.; Tiribelli, C.; Tell, G. Serum AP-Endonuclease 1 (SAPE1) as Novel Biomarker for Hepatocellular Carcinoma. Oncotarget 2019, 10, 383–394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Maso, V.; Avellini, C.; Crocè, L.S.; Rosso, N.; Quadrifoglio, F.; Cesaratto, L.; Codarin, E.; Bedogni, G.; Beltrami, C.A.; Tell, G.; et al. Subcellular Localization of APE1/Ref-1 in Human Hepatocellular Carcinoma: Possible Prognostic Significance. Mol. Med. 2007, 13, 89–96. [Google Scholar] [CrossRef]
- Bazzani, V.; Barchiesi, A.; Radecka, D.; Pravisani, R.; Guadagno, A.; Di Loreto, C.; Baccarani, U.; Vascotto, C. Mitochondrial Apurinic/Apyrimidinic Endonuclease 1 Enhances MtDNA Repair Contributing to Cell Proliferation and Mitochondrial Integrity in Early Stages of Hepatocellular Carcinoma. BMC Cancer 2020, 20, 969. [Google Scholar] [CrossRef]
- Sengupta, S.; Mantha, A.K.; Song, H.; Roychoudhury, S.; Nath, S.; Ray, S.; Bhakat, K.K. Elevated Level of Acetylation of APE1 in Tumor Cells Modulates DNA Damage Repair. Oncotarget 2016, 7, 75197–75209. [Google Scholar] [CrossRef] [Green Version]
- Long, K.; Gu, L.; Li, L.; Zhang, Z.; Li, E.; Zhang, Y.; He, L.; Pan, F.; Guo, Z.; Hu, Z. Small-Molecule Inhibition of APE1 Induces Apoptosis, Pyroptosis, and Necroptosis in Non-Small Cell Lung Cancer. Cell Death Dis. 2021, 12, 503. [Google Scholar] [CrossRef]
- Yoo, D.G.; Song, Y.J.; Cho, E.J.; Lee, S.K.; Park, J.B.; Yu, J.H.; Lim, S.P.; Kim, J.M.; Jeon, B.H. Alteration of APE1/Ref-1 Expression in Non-Small Cell Lung Cancer: The Implications of Impaired Extracellular Superoxide Dismutase and Catalase Antioxidant Systems. Lung Cancer 2008, 60, 277–284. [Google Scholar] [CrossRef]
- Wei, X.; Li, Q.; Li, Y.; Duan, W.; Huang, C.; Zheng, X.; Sun, L.; Luo, J.; Wang, D.; Zhang, S.; et al. Prediction of Survival Prognosis of Non-Small Cell Lung Cancer by APE1 through Regulation of Epithelial-Mesenchymal Transition. Oncotarget 2016, 7, 28523–28539. [Google Scholar] [CrossRef]
- Gu, X.; Cun, Y.; Li, M.; Qing, Y.; Jin, F.; Zhong, Z.; Dai, N.; Qian, C.; Sui, J.; Wang, D. Human Apurinic/Apyrimidinic Endonuclease SiRNA Inhibits the Angiogenesis Induced by X-Ray Irradiation in Lung Cancer Cells. Int. J. Med. Sci. 2013, 10, 870–882. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; He, L.; Dai, N.; Guan, W.; Shan, J.; Yang, X.; Zhong, Z.; Qing, Y.; Jin, F.; Chen, C.; et al. Serum APE1 as a Predictive Marker for Platinum-Based Chemotherapy of Non-Small Cell Lung Cancer Patients. Oncotarget 2016, 7, 77482–77494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kakolyris, S.; Giatromanolaki, A.; Koukourakis, M.; Kaklamanis, L.; Kanavaros, P.; Hickson, I.D.; Barzilay, G.; Georgoulias, V.; Gatter, K.C.; Harris, A.L. Nuclear Localization of Human AP Endonuclease 1 (HAP1/Ref-1) Associates with Prognosis in Early Operable Non-Small Cell Lung Cancer (NSCLC). J. Pathol. 1999, 189, 351–357. [Google Scholar] [CrossRef]
- Puglisi, F.; Aprile, G.; Minisini, A.M.; Barbone, F.; Cataldi, P.; Tell, G.; Kelley, M.R.; Damante, G.; Beltrami, C.A.; Di Loreto, C. Prognostic Significance of Ape1/Ref-1 Subcellular Localization in Non-Small Cell Lung Carcinomas. Anticancer Res. 2001, 21, 4041–4049. [Google Scholar] [PubMed]
- Wu, H.-H.; Chu, Y.-C.; Wang, L.; Tsai, L.-H.; Lee, M.-C.; Chen, C.-Y.; Shieh, S.-H.; Cheng, Y.-W.; Lee, H. Cytoplasmic Ape1 Expression Elevated by P53 Aberration May Predict Survival and Relapse in Resected Non-Small Cell Lung Cancer. Ann. Surg. Oncol. 2013, 20 (Suppl. 3), S336–S347. [Google Scholar] [CrossRef]
- Wu, H.-H.; Cheng, Y.-W.; Chang, J.T.; Wu, T.-C.; Liu, W.-S.; Chen, C.-Y.; Lee, H. Subcellular Localization of Apurinic Endonuclease 1 Promotes Lung Tumor Aggressiveness via NF-κB Activation. Oncogene 2010, 29, 4330–4340. [Google Scholar] [CrossRef] [Green Version]
- Abbotts, R.; Jewell, R.; Nsengimana, J.; Maloney, D.J.; Simeonov, A.; Seedhouse, C.; Elliott, F.; Laye, J.; Walker, C.; Jadhav, A.; et al. Targeting Human Apurinic/Apyrimidinic Endonuclease 1 (APE1) in Phosphatase and Tensin Homolog (PTEN) Deficient Melanoma Cells for Personalized Therapy. Oncotarget 2014, 5, 3273–3286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohammed, M.Z.; Vyjayanti, V.N.; Laughton, C.A.; Dekker, L.V.; Fischer, P.M.; Wilson, D.M.; Abbotts, R.; Shah, S.; Patel, P.M.; Hickson, I.D.; et al. Development and Evaluation of Human AP Endonuclease Inhibitors in Melanoma and Glioma Cell Lines. Br. J. Cancer 2011, 104, 653–663. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Irani, K.; Heffron, S.E.; Jurnak, F.; Meyskens, F.L. Alterations in the Expression of the Apurinic/Apyrimidinic Endonuclease-1/Redox Factor-1 (APE/Ref-1) in Human Melanoma and Identification of the Therapeutic Potential of Resveratrol as an APE/Ref-1 Inhibitor. Mol. Cancer Ther. 2005, 4, 1923–1935. [Google Scholar] [CrossRef] [Green Version]
- Guida, M.; Tommasi, S.; Strippoli, S.; Natalicchio, M.I.; De Summa, S.; Pinto, R.; Cramarossa, A.; Albano, A.; Pisconti, S.; Aieta, M.; et al. The Search for a Melanoma-Tailored Chemotherapy in the New Era of Personalized Therapy: A Phase II Study of Chemo-Modulating Temozolomide Followed by Fotemustine and a Cooperative Study of GOIM (Gruppo Oncologico Italia Meridionale). BMC Cancer 2018, 18, 552. [Google Scholar] [CrossRef] [Green Version]
- Al-Attar, A.; Gossage, L.; Fareed, K.R.; Shehata, M.; Mohammed, M.; Zaitoun, A.M.; Soomro, I.; Lobo, D.N.; Abbotts, R.; Chan, S.; et al. Human Apurinic/Apyrimidinic Endonuclease (APE1) Is a Prognostic Factor in Ovarian, Gastro-Oesophageal and Pancreatico-Biliary Cancers. Br. J. Cancer 2010, 102, 704–709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, J.; Chen, Z.; Peng, D.; Zaika, A.; Revetta, F.; Washington, M.K.; Belkhiri, A.; El-Rifai, W. APE1-Mediated DNA Damage Repair Provides Survival Advantage for Esophageal Adenocarcinoma Cells in Response to Acidic Bile Salts. Oncotarget 2016, 7, 16688–16702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhat, A.A.; Lu, H.; Soutto, M.; Capobianco, A.; Rai, P.; Zaika, A.; El-Rifai, W. Exposure of Barrett’s and Esophageal Adenocarcinoma Cells to Bile Acids Activates EGFR-STAT3 Signaling Axis via Induction of APE1. Oncogene 2018, 37, 6011–6024. [Google Scholar] [CrossRef] [PubMed]
- Sriramajayam, K.; Peng, D.; Lu, H.; Zhou, S.; Bhat, N.; McDonald, O.G.; Que, J.; Zaika, A.; El-Rifai, W. Activation of NRF2 by APE1/REF1 Is Redox-Dependent in Barrett’s Related Esophageal Adenocarcinoma Cells. Redox Biol. 2021, 43, 101970. [Google Scholar] [CrossRef]
- Han, G.; Tian, Y.; Duan, B.; Sheng, H.; Gao, H.; Huang, J. Association of Nuclear Annexin A1 with Prognosis of Patients with Esophageal Squamous Cell Carcinoma. Int. J. Clin. Exp. Pathol. 2014, 7, 751–759. [Google Scholar]
- Song, J.; Futagami, S.; Nagoya, H.; Kawagoe, T.; Yamawaki, H.; Kodaka, Y.; Tatsuguchi, A.; Gudis, K.; Wakabayashi, T.; Yonezawa, M.; et al. Apurinic/Apyrimidinic Endonuclease-1 (APE-1) Is Overexpressed via the Activation of NF-κB-P65 in MCP-1-Positive Esophageal Squamous Cell Carcinoma Tissue. J. Clin. Biochem. Nutr. 2013, 52, 112–119. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Zhang, Z.; Li, Q.; Zhang, L.; Cheng, Y.; Zhong, Z. Mitochondrial APE1 Promotes Cisplatin Resistance by Downregulating ROS in Osteosarcoma. Oncol. Rep. 2020, 44, 499–508. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, Z.; Zhang, L.; Zhong, Z. Cytoplasmic APE1 Promotes Resistance Response in Osteosarcoma Patients with Cisplatin Treatment. Cell Biochem. Funct. 2020, 38, 195–203. [Google Scholar] [CrossRef]
- Wang, D.; Luo, M.; Kelley, M.R. Human Apurinic Endonuclease 1 (APE1) Expression and Prognostic Significance in Osteosarcoma: Enhanced Sensitivity of Osteosarcoma to DNA Damaging Agents Using Silencing RNA APE1 Expression Inhibition. Mol. Cancer Ther. 2004, 3, 679–686. [Google Scholar] [CrossRef]
- Chen, Y.; Yang, Y.; Yuan, Z.; Wang, C.; Shi, Y. Predicting Chemosensitivity in Osteosarcoma Prior to Chemotherapy: An Investigational Study of Biomarkers with Immunohistochemistry. Oncol. Lett. 2012, 3, 1011–1016. [Google Scholar] [CrossRef] [Green Version]
- Ren, T.; Qing, Y.; Dai, N.; Li, M.; Qian, C.; Yang, Y.; Cheng, Y.; Li, Z.; Zhang, S.; Zhong, Z.; et al. Apurinic/Apyrimidinic Endonuclease 1 Induced Upregulation of Fibroblast Growth Factor 2 and Its Receptor 3 Induces Angiogenesis in Human Osteosarcoma Cells. Cancer Sci. 2014, 105, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Shan, J.; Dai, N.; Zhong, Z.; Qing, Y.; Yang, Y.; Zhang, S.; Li, C.; Sui, J.; Ren, T.; et al. Apurinic/Apyrimidinic Endonuclease 1 Regulates Angiogenesis in a Transforming Growth Factor β-Dependent Manner in Human Osteosarcoma. Cancer Sci. 2015, 106, 1394–1401. [Google Scholar] [CrossRef] [Green Version]
- Dai, N.; Qing, Y.; Cun, Y.; Zhong, Z.; Li, C.; Zhang, S.; Shan, J.; Yang, X.; Dai, X.; Cheng, Y.; et al. MiR-513a-5p Regulates Radiosensitivity of Osteosarcoma by Targeting Human Apurinic/Apyrimidinic Endonuclease. Oncotarget 2016, 9, 25414–25426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Londero, A.P.; Orsaria, M.; Tell, G.; Marzinotto, S.; Capodicasa, V.; Poletto, M.; Vascotto, C.; Sacco, C.; Mariuzzi, L. Expression and Prognostic Significance of APE1/Ref-1 and NPM1 Proteins in High-Grade Ovarian Serous Cancer. Am. J. Clin. Pathol. 2014, 141, 404–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen, X.; Lu, R.; Xie, S.; Zheng, H.; Wang, H.; Wang, Y.; Sun, J.; Gao, X.; Guo, L. APE1 Overexpression Promotes the Progression of Ovarian Cancer and Serves as a Potential Therapeutic Target. Cancer Biomark. 2016, 17, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, J.; Xiang, D.; Wang, D.; Xin, X. Alterations in the Expression of the Apurinic/Apyrimidinic Endonuclease-1/Redox Factor-1 (APE1/Ref-1) in Human Ovarian Cancer and Indentification of the Therapeutic Potential of APE1/Ref-1 Inhibitor. Int. J. Oncol. 2009, 35, 1069–1079. [Google Scholar]
- Fan, X.; Wen, L.; Li, Y.; Lou, L.; Liu, W.; Zhang, J. The Expression Profile and Prognostic Value of APE/Ref-1 and NPM1 in High-Grade Serous Ovarian Adenocarcinoma. APMIS 2017, 125, 857–862. [Google Scholar] [CrossRef]
- Poletto, M.; Malfatti, M.C.; Dorjsuren, D.; Scognamiglio, P.L.; Marasco, D.; Vascotto, C.; Jadhav, A.; Maloney, D.J.; Wilson, D.M.; Simeonov, A.; et al. Inhibitors of the Apurinic/Apyrimidinic Endonuclease 1 (APE1)/Nucleophosmin (NPM1) Interaction That Display Anti-Tumor Properties. Mol. Carcinog. 2016, 55, 688–704. [Google Scholar] [CrossRef] [Green Version]
- Sheng, Q.; Zhang, Y.; Wang, R.; Zhang, J.; Chen, B.; Wang, J.; Zhang, W.; Xin, X. Prognostic Significance of APE1 Cytoplasmic Localization in Human Epithelial Ovarian Cancer. Med. Oncol. 2012, 29, 1265–1271. [Google Scholar] [CrossRef]
- Moore, D.H.; Michael, H.; Tritt, R.; Parsons, S.H.; Kelley, M.R. Alterations in the Expression of the DNA Repair/Redox Enzyme APE/Ref-1 in Epithelial Ovarian Cancers. Clin. Cancer Res. 2000, 6, 602–609. [Google Scholar]
- Tanner, B.; Grimme, S.; Schiffer, I.; Heimerdinger, C.; Schmidt, M.; Dutkowski, P.; Neubert, S.; Oesch, F.; Franzen, A.; Kölbl, H.; et al. Nuclear Expression of Apurinic/Apyrimidinic Endonuclease Increases with Progression of Ovarian Carcinomas. Gynecol. Oncol. 2004, 92, 568–577. [Google Scholar] [CrossRef]
- Pramanik, S.; Chen, Y.; Song, H.; Khutsishvili, I.; Marky, L.A.; Ray, S.; Natarajan, A.; Singh, P.K.; Bhakat, K.K. The Human AP-Endonuclease 1 (APE1) Is a DNA G-Quadruplex Structure Binding Protein and Regulates KRAS Expression in Pancreatic Ductal Adenocarcinoma Cells. Nucleic Acids Res. 2022, 50, 3394–3412. [Google Scholar] [CrossRef]
- Jiang, Y.; Zhou, S.; Sandusky, G.E.; Kelley, M.R.; Fishel, M.L. Reduced Expression of DNA Repair and Redox Signaling Protein APE1/Ref-1 Impairs Human Pancreatic Cancer Cell Survival, Proliferation, and Cell Cycle Progression. Cancer Investig. 2010, 28, 885–895. [Google Scholar] [CrossRef] [PubMed]
- Fishel, M.L.; Jiang, Y.; Rajeshkumar, N.V.; Scandura, G.; Sinn, A.L.; He, Y.; Shen, C.; Jones, D.R.; Pollok, K.E.; Ivan, M.; et al. Impact of APE1/Ref-1 Redox Inhibition on Pancreatic Tumor Growth. Mol. Cancer Ther. 2011, 10, 1698–1708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelley, M.R.; Cheng, L.; Foster, R.; Tritt, R.; Jiang, J.; Broshears, J.; Koch, M. Elevated and Altered Expression of the Multifunctional DNA Base Excision Repair and Redox Enzyme Ape1/Ref-1 in Prostate Cancer. Clin. Cancer Res. 2001, 7, 824–830. [Google Scholar] [PubMed]
- Juhnke, M.; Heumann, A.; Chirico, V.; Höflmayer, D.; Menz, A.; Hinsch, A.; Hube-Magg, C.; Kluth, M.; Lang, D.S.; Möller-Koop, C.; et al. Apurinic/Apyrimidinic Endonuclease 1 (APE1/Ref-1) Overexpression Is an Independent Prognostic Marker in Prostate Cancer without TMPRSS2:ERG Fusion. Mol. Carcinog. 2017, 56, 2135–2145. [Google Scholar] [CrossRef]
- Silva, L.P.; Santana, T.; Sedassari, B.T.; de Sousa, S.M.; Sobral, A.P.V.; Freitas, R.d.A.; Barboza, C.A.G.; de Souza, L.B. Apurinic/Apyrimidinic Endonuclease 1 (APE1) Is Overexpressed in Malignant Transformation of Salivary Gland Pleomorphic Adenoma. Eur. Arch. Otorhinolaryngol. 2017, 274, 3203–3209. [Google Scholar] [CrossRef] [PubMed]
- Felix, F.A.; da Silva, L.P.; Lopes, M.L.D.d.S.; Sobral, A.P.V.; Freitas, R.d.A.; de Souza, L.B.; Barboza, C.A.G. DNA Base Excision Repair and Nucleotide Excision Repair Proteins in Malignant Salivary Gland Tumors. Arch. Oral. Biol. 2021, 121, 104987. [Google Scholar] [CrossRef]
- Lee, Y.R.; Lim, J.S.; Shin, J.H.; Choi, S.; Joo, H.K.; Jeon, B.H. Altered Secretory Activity of APE1/Ref-1 D148E Variants Identified in Human Patients With Bladder Cancer. Int. Neurourol. J. 2016, 20, S30–S37. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Wilson, D.M. Human Apurinic/Apyrimidinic Endonuclease 1. Antioxid. Redox Signal. 2014, 20, 678–707. [Google Scholar] [CrossRef] [Green Version]
- Castillo-Acosta, V.M.; Ruiz-Perez, L.M.; Yang, W.; Gonzalez-Pacanowska, D.; Vidal, A.E. Identification of a Residue Critical for the Excision of 3′-Blocking Ends in Apurinic/Apyrimidinic Endonucleases of the Xth Family. Nucleic Acids Res. 2009, 37, 1829–1842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Timofeyeva, N.A.; Koval, V.V.; Ishchenko, A.A.; Saparbaev, M.K.; Fedorova, O.S. Lys98 Substitution in Human AP Endonuclease 1 Affects the Kinetic Mechanism of Enzyme Action in Base Excision and Nucleotide Incision Repair Pathways. PLoS ONE 2011, 6, e24063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitaker, A.M.; Flynn, T.S.; Freudenthal, B.D. Molecular Snapshots of APE1 Proofreading Mismatches and Removing DNA Damage. Nat. Commun. 2018, 9, 399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, D.M.; Simeonov, A. Small Molecule Inhibitors of DNA Repair Nuclease Activities of APE1. Cell. Mol. Life Sci. 2010, 67, 3621–3631. [Google Scholar] [CrossRef] [Green Version]
- Al-Safi, R.I.; Odde, S.; Shabaik, Y.; Neamati, N. Small-Molecule Inhibitors of APE1 DNA Repair Function: An Overview. Curr. Mol. Pharmacol. 2012, 5, 14–35. [Google Scholar] [CrossRef]
- Laev, S.S.; Salakhutdinov, N.F.; Lavrik, O.I. Inhibitors of Nuclease and Redox Activity of Apurinic/Apyrimidinic Endonuclease 1/Redox Effector Factor 1 (APE1/Ref-1). Bioorg. Med. Chem. 2017, 25, 2531–2544. [Google Scholar] [CrossRef]
- Caston, R.A.; Gampala, S.; Armstrong, L.; Messmann, R.A.; Fishel, M.L.; Kelley, M.R. The Multifunctional APE1 DNA Repair-Redox Signaling Protein as a Drug Target in Human Disease. Drug Discov. Today 2021, 26, 218–228. [Google Scholar] [CrossRef]
- Xue, Z.; Demple, B. Knockout and Inhibition of Ape1: Roles of Ape1 in Base Excision DNA Repair and Modulation of Gene Expression. Antioxidants 2022, 11, 1817. [Google Scholar] [CrossRef]
- Liuzzi, M.; Weinfeld, M.; Paterson, M.C. Selective Inhibition by Methoxyamine of the Apurinic/Apyrimidinic Endonuclease Activity Associated with Pyrimidine Dimer-DNA Glycosylases from Micrococcus Luteus and Bacteriophage T4. Biochemistry 1987, 26, 3315–3321. [Google Scholar] [CrossRef]
- Liu, L.; Gerson, S.L. Therapeutic Impact of Methoxyamine: Blocking Repair of Abasic Sites in the Base Excision Repair Pathway. Curr. Opin. Investig. Drugs 2004, 5, 623–627. [Google Scholar]
- Liu, L.; Nakatsuru, Y.; Gerson, S.L. Base Excision Repair as a Therapeutic Target in Colon Cancer. Clin. Cancer Res. 2002, 8, 2985–2991. [Google Scholar] [PubMed]
- Fishel, M.L.; He, Y.; Smith, M.L.; Kelley, M.R. Manipulation of Base Excision Repair to Sensitize Ovarian Cancer Cells to Alkylating Agent Temozolomide. Clin. Cancer Res. 2007, 13, 260–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bases, R. Enhancement of X-Ray Damage in HeLa Cells by Exposure to Lucanthone (Miracil D) Following Radiation. Cancer Res. 1970, 30, 2007–2011. [Google Scholar] [PubMed]
- Luo, M.; Kelley, M.R. Inhibition of the Human Apurinic/Apyrimidinic Endonuclease (Ape1) Repair Activity and Sensitization of Breast Cancer Cells to DNA Alkylating Agents with Lucanthone. Anticancer Res. 2004, 24, 2127–2134. [Google Scholar] [PubMed]
- Naidu, M.D.; Agarwal, R.; Pena, L.A.; Cunha, L.; Mezei, M.; Shen, M.; Wilson, D.M.; Liu, Y.; Sanchez, Z.; Chaudhary, P.; et al. Lucanthone and Its Derivative Hycanthone Inhibit Apurinic Endonuclease-1 (APE1) by Direct Protein Binding. PLoS ONE 2011, 6, e23679. [Google Scholar] [CrossRef] [Green Version]
- Madhusudan, S.; Smart, F.; Shrimpton, P.; Parsons, J.L.; Gardiner, L.; Houlbrook, S.; Talbot, D.C.; Hammonds, T.; Freemont, P.A.; Sternberg, M.J.E.; et al. Isolation of a Small Molecule Inhibitor of DNA Base Excision Repair. Nucleic Acids Res. 2005, 33, 4711–4724. [Google Scholar] [CrossRef] [Green Version]
- Simeonov, A.; Kulkarni, A.; Dorjsuren, D.; Jadhav, A.; Shen, M.; McNeill, D.R.; Austin, C.P.; Wilson, D.M. Identification and Characterization of Inhibitors of Human Apurinic/Apyrimidinic Endonuclease APE1. PLoS ONE 2009, 4, e5740. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Y.; Fu, D.; Xu, Y.; Wang, X.; Deng, X.; Zhou, S.; Du, F.; Cui, X.; Deng, Y.; Tang, Z. Pt(IV) Prodrug as a Potential Antitumor Agent with APE1 Inhibitory Activity. J. Med. Chem. 2022, 65, 15344–15357. [Google Scholar] [CrossRef]
- Seiple, L.A.; Cardellina, J.H.; Akee, R.; Stivers, J.T. Potent Inhibition of Human Apurinic/Apyrimidinic Endonuclease 1 by Arylstibonic Acids. Mol. Pharmacol. 2008, 73, 669–677. [Google Scholar] [CrossRef] [Green Version]
- Zawahir, Z.; Dayam, R.; Deng, J.; Pereira, C.; Neamati, N. Pharmacophore Guided Discovery of Small-Molecule Human Apurinic/Apyrimidinic Endonuclease 1 Inhibitors. J. Med. Chem. 2009, 52, 20–32. [Google Scholar] [CrossRef]
- Bapat, A.; Glass, L.S.; Luo, M.; Fishel, M.L.; Long, E.C.; Georgiadis, M.M.; Kelley, M.R. Novel Small-Molecule Inhibitor of Apurinic/Apyrimidinic Endonuclease 1 Blocks Proliferation and Reduces Viability of Glioblastoma Cells. J. Pharmacol. Exp. Ther. 2010, 334, 988–998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aiello, F.; Shabaik, Y.; Esqueda, A.; Sanchez, T.W.; Grande, F.; Garofalo, A.; Neamati, N. Design and Synthesis of 3-Carbamoylbenzoic Acid Derivatives as Inhibitors of Human Apurinic/Apyrimidinic Endonuclease 1 (APE1). ChemMedChem 2012, 7, 1825–1839. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, F.M.; Francis, S.M.; Tintoré, M.; Ferreira, R.; Gil-Redondo, R.; Morreale, A.; Ortiz, Á.R.; Eritja, R.; Fàbrega, C. Receptor-Based Virtual Screening and Biological Characterization of Human Apurinic/Apyrimidinic Endonuclease (Ape1) Inhibitors. ChemMedChem 2012, 7, 2168–2178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dorjsuren, D.; Kim, D.; Vyjayanti, V.N.; Maloney, D.J.; Jadhav, A.; Wilson, D.M.; Simeonov, A. Diverse Small Molecule Inhibitors of Human Apurinic/Apyrimidinic Endonuclease APE1 Identified from a Screen of a Large Public Collection. PLoS ONE 2012, 7, e47974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rai, G.; Vyjayanti, V.N.; Dorjsuren, D.; Simeonov, A.; Jadhav, A.; Wilson, D.M.; Maloney, D.J. Synthesis, Biological Evaluation, and Structure–Activity Relationships of a Novel Class of Apurinic/Apyrimidinic Endonuclease 1 Inhibitors. J. Med. Chem. 2012, 55, 3101–3112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srinivasan, A.; Wang, L.; Cline, C.J.; Xie, Z.; Sobol, R.W.; Xie, X.-Q.; Gold, B. Identification and Characterization of Human Apurinic/Apyrimidinic Endonuclease-1 Inhibitors; American Chemical Society: Washington, DC, USA, 2012; Volume 51, ISBN 00062960. [Google Scholar]
- Feng, Z.; Kochanek, S.; Close, D.; Wang, L.; Srinivasan, A.; Almehizia, A.A.; Iyer, P.; Xie, X.-Q.; Johnston, P.A.; Gold, B. Design and Activity of AP Endonuclease-1 Inhibitors. J. Chem. Biol. 2015, 8, 79–93. [Google Scholar] [CrossRef] [Green Version]
- Trilles, R.; Beglov, D.; Chen, Q.; He, H.; Wireman, R.; Reed, A.; Chennamadhavuni, S.; Panek, J.S.; Brown, L.E.; Vajda, S.; et al. Discovery of Macrocyclic Inhibitors of Apurinic/Apyrimidinic Endonuclease 1. J. Med. Chem. 2019, 62, 1971–1988. [Google Scholar] [CrossRef]
- Pidugu, L.S.; Servius, H.W.; Sevdalis, S.E.; Cook, M.E.; Varney, K.M.; Pozharski, E.; Drohat, A.C. Characterizing Inhibitors of Human AP Endonuclease 1. PLoS ONE 2023, 18, e0280526. [Google Scholar] [CrossRef]
- Masani, S.; Han, L.; Yu, K. Apurinic/Apyrimidinic Endonuclease 1 Is the Essential Nuclease during Immunoglobulin Class Switch Recombination. Mol. Cell. Biol. 2013, 33, 1468–1473. [Google Scholar] [CrossRef] [Green Version]
- Georgiadis, M.M.; Luo, M.; Gaur, R.K.; Delaplane, S.; Li, X.; Kelley, M.R. Evolution of the Redox Function in Mammalian Apurinic/Apyrimidinic Endonuclease. Mutat. Res./Fundam. Mol. Mech. Mutagen. 2008, 643, 54–63. [Google Scholar] [CrossRef] [Green Version]
- Ando, K.; Hirao, S.; Kabe, Y.; Ogura, Y.; Sato, I.; Yamaguchi, Y.; Wada, T.; Handa, H. A New APE1/Ref-1-Dependent Pathway Leading to Reduction of NF-κB and AP-1, and Activation of Their DNA-Binding Activity. Nucleic Acids Res. 2008, 36, 4327–4336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Logsdon, D.P.; Grimard, M.; Luo, M.; Shahda, S.; Jiang, Y.; Tong, Y.; Yu, Z.; Zyromski, N.; Schipani, E.; Carta, F.; et al. Regulation of HIF1α under Hypoxia by APE1/Ref-1 Impacts CA9 Expression: Dual Targeting in Patient-Derived 3D Pancreatic Cancer Models. Mol. Cancer Ther. 2016, 15, 2722–2732. [Google Scholar] [CrossRef] [Green Version]
- Ray, S.; Lee, C.; Hou, T.; Bhakat, K.K.; Brasier, A.R. Regulation of Signal Transducer and Activator of Transcription 3 Enhanceosome Formation by Apurinic/Apyrimidinic Endonuclease 1 in Hepatic Acute Phase Response. Mol. Endocrinol. 2010, 24, 391–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardoso, A.A.; Jiang, Y.; Luo, M.; Reed, A.M.; Shahda, S.; He, Y.; Maitra, A.; Kelley, M.R.; Fishel, M.L. APE1/Ref-1 Regulates STAT3 Transcriptional Activity and APE1/Ref-1–STAT3 Dual-Targeting Effectively Inhibits Pancreatic Cancer Cell Survival. PLoS ONE 2012, 7, e47462. [Google Scholar] [CrossRef]
- Jayaraman, L.; Murthy, K.G.; Zhu, C.; Curran, T.; Xanthoudakis, S.; Prives, C. Identification of Redox/Repair Protein Ref-1 as a Potent Activator of P53. Genes Dev. 1997, 11, 558–570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fishel, M.L.; Wu, X.; Devlin, C.M.; Logsdon, D.P.; Jiang, Y.; Luo, M.; He, Y.; Yu, Z.; Tong, Y.; Lipking, K.P.; et al. Apurinic/Apyrimidinic Endonuclease/Redox Factor-1 (APE1/Ref-1) Redox Function Negatively Regulates NRF2. J. Biol. Chem. 2015, 290, 3057–3068. [Google Scholar] [CrossRef] [PubMed]
- Tell, G.; Scaloni, A.; Pellizzari, L.; Formisano, S.; Pucillo, C.; Damante, G. Redox Potential Controls the Structure and DNA Binding Activity of the Paired Domain. J. Biol. Chem. 1998, 273, 25062–25072. [Google Scholar] [CrossRef] [Green Version]
- Evans, A.R.; Limp-Foster, M.; Kelley, M.R. Going APE over Ref-1. Mutat. Res. DNA Repair 2000, 461, 83–108. [Google Scholar] [CrossRef]
- Xanthoudakis, S.; Curran, T. Identification and Characterization of Ref-1, a Nuclear Protein That Facilitates AP-1 DNA-Binding Activity. EMBO J. 1992, 11, 653–665. [Google Scholar] [CrossRef]
- Xanthoudakis, S.; Miao, G.; Wang, F.; Pan, Y.C.; Curran, T. Redox Activation of Fos-Jun DNA Binding Activity Is Mediated by a DNA Repair Enzyme. EMBO J. 1992, 11, 3323–3335. [Google Scholar] [CrossRef]
- Arlt, A.; Vorndamm, J.; Breitenbroich, M.; Fölsch, U.R.; Kalthoff, H.; Schmidt, W.E.; Schäfer, H. Inhibition of NF-ΚB Sensitizes Human Pancreatic Carcinoma Cells to Apoptosis Induced by Etoposide (VP16) or Doxorubicin. Oncogene 2001, 20, 859–868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arlt, A.; Gehrz, A.; Müerköster, S.; Vorndamm, J.; Kruse, M.-L.; Fölsch, U.R.; Schäfer, H. Role of NF-ΚB and Akt/PI3K in the Resistance of Pancreatic Carcinoma Cell Lines against Gemcitabine-Induced Cell Death. Oncogene 2003, 22, 3243–3251. [Google Scholar] [CrossRef] [Green Version]
- Raffoul, J.J.; Heydari, A.R.; Hillman, G.G. DNA Repair and Cancer Therapy: Targeting APE1/Ref-1 Using Dietary Agents. J. Oncol. 2012, 2012, 370481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, S.C.; Patchva, S.; Aggarwal, B.B. Therapeutic Roles of Curcumin: Lessons Learned from Clinical Trials. AAPS J. 2013, 15, 195–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Zhong, C.; Wang, Q.; Chen, W.; Yuan, Y. Curcumin Is an APE1 Redox Inhibitor and Exhibits an Antiviral Activity against KSHV Replication and Pathogenesis. Antivir. Res. 2019, 167, 98–103. [Google Scholar] [CrossRef]
- Escobar, I.; Xu, J.; Jackson, C.W.; Stegelmann, S.D.; Fagerli, E.A.; Dave, K.R.; Perez-Pinzon, M.A. Resveratrol Preconditioning Protects Against Ischemia-Induced Synaptic Dysfunction and Cofilin Hyperactivation in the Mouse Hippocampal Slice. Neurotherapeutics 2023. [Google Scholar] [CrossRef]
- Raffoul, J.J.; Banerjee, S.; Singh-Gupta, V.; Knoll, Z.E.; Fite, A.; Zhang, H.; Abrams, J.; Sarkar, F.H.; Hillman, G.G. Down-Regulation of Apurinic/Apyrimidinic Endonuclease 1/Redox Factor-1 Expression by Soy Isoflavones Enhances Prostate Cancer Radiotherapy In Vitro and In Vivo. Cancer Res. 2007, 67, 2141–2149. [Google Scholar] [CrossRef] [Green Version]
- Kelley, M.R.; Wikel, J.H.; Guo, C.; Pollok, K.E.; Bailey, B.J.; Wireman, R.; Fishel, M.L.; Vasko, M.R. Identification and Characterization of New Chemical Entities Targeting Apurinic/Apyrimidinic Endonuclease 1 for the Prevention of Chemotherapy-Induced Peripheral Neuropathy. J. Pharmacol. Exp. Ther. 2016, 359, 300–309. [Google Scholar] [CrossRef] [Green Version]
- Shimizu, N.; Sugimoto, K.; Tang, J.; Nishi, T.; Sato, I.; Hiramoto, M.; Aizawa, S.; Hatakeyama, M.; Ohba, R.; Hatori, H.; et al. High-Performance Affinity Beads for Identifying Drug Receptors. Nat. Biotechnol. 2000, 18, 877–881. [Google Scholar] [CrossRef]
- Zou, G.-M.; Karikari, C.; Kabe, Y.; Handa, H.; Anders, R.A.; Maitra, A. The Ape-1/Ref-1 Redox Antagonist E3330 Inhibits the Growth of Tumor Endothelium and Endothelial Progenitor Cells: Therapeutic Implications in Tumor Angiogenesis. J. Cell. Physiol. 2009, 219, 209–218. [Google Scholar] [CrossRef]
- Zou, G.-M.; Maitra, A. Small-Molecule Inhibitor of the AP Endonuclease 1/REF-1 E3330 Inhibits Pancreatic Cancer Cell Growth and Migration. Mol. Cancer Ther. 2008, 7, 2012–2021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, A.; Gao, H.; Kelley, M.R.; Qiao, X. Inhibition of APE1/Ref-1 Redox Activity with APX3330 Blocks Retinal Angiogenesis in Vitro and in Vivo. Vis. Res. 2011, 51, 93–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manguinhas, R.; Fernandes, A.S.; Costa, J.G.; Saraiva, N.; Camões, S.P.; Gil, N.; Rosell, R.; Castro, M.; Miranda, J.P.; Oliveira, N.G. Impact of the APE1 Redox Function Inhibitor E3330 in Non-Small Cell Lung Cancer Cells Exposed to Cisplatin: Increased Cytotoxicity and Impairment of Cell Migration and Invasion. Antioxidants 2020, 9, 550. [Google Scholar] [CrossRef] [PubMed]
- Guerreiro, P.S.; Corvacho, E.; Costa, J.G.; Saraiva, N.; Fernandes, A.S.; Castro, M.; Miranda, J.P.; Oliveira, N.G. The APE1 Redox Inhibitor E3330 Reduces Collective Cell Migration of Human Breast Cancer Cells and Decreases Chemoinvasion and Colony Formation When Combined with Docetaxel. Chem. Biol. Drug Des. 2017, 90, 561–571. [Google Scholar] [CrossRef]
- Li, Y.; Liu, X.; Zhou, T.; Kelley, M.R.; Edwards, P.A.; Gao, H.; Qiao, X. Suppression of Choroidal Neovascularization Through Inhibition of APE1/Ref-1 Redox Activity. Investig. Ophthalmol. Vis. Sci. 2014, 55, 4461–4469. [Google Scholar] [CrossRef] [PubMed]
- Hartman, G.D.; Lambert-Cheatham, N.A.; Kelley, M.R.; Corson, T.W. Inhibition of APE1/Ref-1 for Neovascular Eye Diseases: From Biology to Therapy. Int. J. Mol. Sci. 2021, 22, 10279. [Google Scholar] [CrossRef]
- Hu, J.; Wang, Y.; Yuan, Y. Inhibitors of APE1 Redox Function Effectively Inhibit γ-Herpesvirus Replication in Vitro and in Vivo. Antivir. Res. 2021, 185, 104985. [Google Scholar] [CrossRef]
- Shahda, S.; Lakhani, N.J.; O’Neil, B.; Rasco, D.W.; Wan, J.; Mosley, A.L.; Liu, H.; Kelley, M.R.; Messmann, R.A. A Phase I Study of the APE1 Protein Inhibitor APX3330 in Patients with Advanced Solid Tumors. JCO 2019, 37 (Suppl. 15), 3097. [Google Scholar] [CrossRef]
- Kelley, M.R.; Luo, M.; Reed, A.; Su, D.; Delaplane, S.; Borch, R.F.; Nyland, R.L.; Gross, M.L.; Georgiadis, M.M. Functional Analysis of Novel Analogues of E3330 That Block the Redox Signaling Activity of the Multifunctional AP Endonuclease/Redox Signaling Enzyme APE1/Ref-1. Antioxid. Redox Signal. 2011, 14, 1387–1401. [Google Scholar] [CrossRef] [Green Version]
- Nyland, R.L.; Luo, M.; Kelley, M.R.; Borch, R.F. Design and Synthesis of Novel Quinone Inhibitors Targeted to the Redox Function of Apurinic/Apyrimidinic Endonuclease 1/Redox Enhancing Factor-1 (Ape1/Ref-1). J. Med. Chem. 2010, 53, 1200–1210. [Google Scholar] [CrossRef] [Green Version]
- Qian, C.; Li, M.; Sui, J.; Ren, T.; Li, Z.; Zhang, L.; Zhou, L.; Cheng, Y.; Wang, D. Identification of a Novel Potential Antitumor Activity of Gossypol as an APE1/Ref-1 Inhibitor. Drug Des. Dev. Ther. 2014, 8, 485–496. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Li, X.; Zhang, L.; Li, M.; Dai, N.; Luo, H.; Shan, J.; Yang, X.; Xu, M.; Feng, Y.; et al. A Randomized, Double-Blind, Placebo-Controlled Study of B-Cell Lymphoma 2 Homology 3 Mimetic Gossypol Combined with Docetaxel and Cisplatin for Advanced Non-Small Cell Lung Cancer with High Expression of Apurinic/Apyrimidinic Endonuclease 1. Investig. New Drugs 2020, 38, 1862–1871. [Google Scholar] [CrossRef] [PubMed]
- Ren, T.; Shan, J.; Qing, Y.; Qian, C.; Li, Q.; Lu, G.; Li, M.; Li, C.; Peng, Y.; Luo, H.; et al. Sequential Treatment with AT-101 Enhances Cisplatin Chemosensitivity in Human Non-Small Cell Lung Cancer Cells through Inhibition of Apurinic/Apyrimidinic Endonuclease 1-Activated IL-6/STAT3 Signaling Pathway. Drug Des. Dev. Ther. 2014, 8, 2517–2529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, X.; Duan, W.; Li, Y.; Zhang, S.; Xin, X.; Sun, L.; Gao, M.; Li, Q.; Wang, D. AT101 Exerts a Synergetic Efficacy in Gastric Cancer Patients with 5-FU Based Treatment through Promoting Apoptosis and Autophagy. Oncotarget 2016, 7, 34430–34441. [Google Scholar] [CrossRef] [Green Version]
- Poletto, M.; Vascotto, C.; Scognamiglio, P.L.; Lirussi, L.; Marasco, D.; Tell, G. Role of the Unstructured N-Terminal Domain of the HAPE1 (Human Apurinic/Apyrimidinic Endonuclease 1) in the Modulation of Its Interaction with Nucleic Acids and NPM1 (Nucleophosmin). Biochem. J. 2013, 452, 545–557. [Google Scholar] [CrossRef] [Green Version]
- Antoniali, G.; Dalla, E.; Mangiapane, G.; Zhao, X.; Jing, X.; Cheng, Y.; De Sanctis, V.; Ayyildiz, D.; Piazza, S.; Li, M.; et al. APE1 Controls DICER1 Expression in NSCLC through MiR-33a and MiR-130b. Cell. Mol. Life Sci. 2022, 79, 446. [Google Scholar] [CrossRef]
- Vascotto, C.; Cesaratto, L.; Zeef, L.A.H.; Deganuto, M.; D’Ambrosio, C.; Scaloni, A.; Romanello, M.; Damante, G.; Taglialatela, G.; Delneri, D.; et al. Genome-Wide Analysis and Proteomic Studies Reveal APE1/Ref-1 Multifunctional Role in Mammalian Cells. Proteomics 2009, 9, 1058–1074. [Google Scholar] [CrossRef] [Green Version]
- Ayyildiz, D.; Antoniali, G.; D’Ambrosio, C.; Mangiapane, G.; Dalla, E.; Scaloni, A.; Tell, G.; Piazza, S. Architecture of The Human Ape1 Interactome Defines Novel Cancers Signatures. Sci. Rep. 2020, 10, 28. [Google Scholar] [CrossRef] [Green Version]
- Wilson, D.M.; Deacon, A.M.; Duncton, M.A.J.; Pellicena, P.; Georgiadis, M.M.; Yeh, A.P.; Arvai, A.S.; Moiani, D.; Tainer, J.A.; Das, D. Fragment- and Structure-Based Drug Discovery for Developing Therapeutic Agents Targeting the DNA Damage Response. Prog. Biophys. Mol. Biol. 2021, 163, 130–142. [Google Scholar] [CrossRef]
- Codrich, M.; Degrassi, M.; Malfatti, M.C.; Antoniali, G.; Gorassini, A.; Ayyildiz, D.; De Marco, R.; Verardo, G.; Tell, G. APE1 Interacts with the Nuclear Exosome Complex Protein MTR4 and Is Involved in Cisplatin- and 5-Fluorouracil-Induced RNA Damage Response. FEBS J. 2022, 290, 1740–1764. [Google Scholar] [CrossRef]
- Garutti, M.; Pelizzari, G.; Bartoletti, M.; Malfatti, M.C.; Gerratana, L.; Tell, G.; Puglisi, F. Platinum Salts in Patients with Breast Cancer: A Focus on Predictive Factors. Int. J. Mol. Sci. 2019, 20, 3390. [Google Scholar] [CrossRef] [Green Version]
- Dumas, L.; Herviou, P.; Dassi, E.; Cammas, A.; Millevoi, S. G-Quadruplexes in RNA Biology: Recent Advances and Future Directions. Trends Biochem. Sci. 2021, 46, 270–283. [Google Scholar] [CrossRef]
- Kwok, C.K.; Sahakyan, A.B.; Balasubramanian, S. Structural Analysis Using SHALiPE to Reveal RNA G-Quadruplex Formation in Human Precursor MicroRNA. Angew. Chem. Int. Ed. 2016, 55, 8958–8961. [Google Scholar] [CrossRef]
- Figueiredo, J.; Santos, T.; Miranda, A.; Alexandre, D.; Teixeira, B.; Simões, P.; Lopes-Nunes, J.; Cruz, C. Ligands as Stabilizers of G-Quadruplexes in Non-Coding RNAs. Molecules 2021, 26, 6164. [Google Scholar] [CrossRef] [PubMed]
- Koralewska, N.; Szczepanska, A.; Ciechanowska, K.; Wojnicka, M.; Pokornowska, M.; Milewski, M.C.; Gudanis, D.; Baranowski, D.; Nithin, C.; Bujnicki, J.M.; et al. RNA and DNA G-Quadruplexes Bind to Human Dicer and Inhibit Its Activity. Cell. Mol. Life Sci. 2021, 78, 3709–3724. [Google Scholar] [CrossRef]
- Asamitsu, S.; Shioda, N. Potential Roles of G-Quadruplex Structures in RNA Granules for Physiological and Pathological Phase Separation. J. Biochem. 2021, 169, 527–533. [Google Scholar] [CrossRef]
- Zhang, Y.; Yang, M.; Duncan, S.; Yang, X.; Abdelhamid, M.A.S.; Huang, L.; Zhang, H.; Benfey, P.N.; Waller, Z.A.E.; Ding, Y. G-Quadruplex Structures Trigger RNA Phase Separation. Nucleic Acids Res. 2019, 47, 11746–11754. [Google Scholar] [CrossRef] [PubMed]
- Mirihana Arachchilage, G.; Dassanayake, A.C.; Basu, S. A Potassium Ion-Dependent RNA Structural Switch Regulates Human Pre-MiRNA 92b Maturation. Chem. Biol. 2015, 22, 262–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]


| Cancer | Polymorphisms |
|---|---|
| Adrenocortical carcinoma | D283H; X20_splice |
| Ampullary cancer | M271T |
| Bladder cancer | D15H; S146L; Q109E; E217 * |
| Bone cancer | V131M |
| Breast cancer | L244Tfs * 8; Q51 *; H289Y; G5A; K7Rfs * 75; R185W; A60G |
| Cervical cancer | R281C; K7R |
| Colorectal cancer | A273T; R193C; R221H; G130D; R247Q; A230D; M271I; R221H; R181 *; E242D; R274 *; P293S; P49Qfs * 33; L220I; T233M; N226Efs * 26; V131M; L291Vfs * 6; R221C; K63E; D210N; R281H; X82_splice; E101D; K77N; P331H; G306S; G132D |
| Endometrial cancer | R193H; S164L; R221H; A170V; P223L; V84I; V278D; R281H; K228T |
| Gastric cancer | R281C; P122A; K27N |
| Glioma | H289Q; Q245R; G80E; D142N |
| Head and Neck cancer | P48H |
| Leukemia and Lymphoma | R181Q; L17P; K7R; R187H |
| Liver cancer | G5E; W280S; L220P; W280R; D50Rfs * 28; G8R; H289Y, N226Efs * 26; Y264_G279del |
| Lung cancer | G41C; V206Cfs * 11; V142Sfs * 8; E16Q; P331T; E149Q; X147_splice; R177*; D90H; Q51 *; X237_splice; R193C; R28S; M271del; V206Cfs * 11; S115F; G8R; I146V; D148E |
| Melanoma | G241W; E16K; G127V; R136S; P122T; D148E; A263V; K7Rfs * 75; L108F; V69L |
| Oesophageal cancer | E46D; D251N; M270Nfs * 14; F165V; N102I; L291Vfs * 6; K3R; G145D |
| Ovarian cancer | Q95 *, V168I; R193H; L291Vfs * 6 |
| Pancreatic cancer | R193C; M271del |
| Prostate cancer | P139Q; A30T; R187H |
| Renal cancer | E149 * |
| Sarcoma | R187L; K35Rfs * 11; K35Q |
| Skin cancer | P89S |
| Cancer | Expression | Diagnostic Value | Localization | Refs |
|---|---|---|---|---|
| Bladder cancer (Bca) | Protein overexpression, associated with poor survival and invasion. | Serum and urine levels as a diagnostic biomarker. |
| [38,39,40,41,42,43,44] |
| Breast cancer |
| n.d. | Nuclear localization. | [45,46,47,48,49,50] |
| Cervical cancer | High protein expression is associated with lymph node metastasis, EMT, and decreasing radiosensitivity. | n.d. |
| [51,52,53,54] |
| Colorectal cancer (CRC) |
| Serum APE1-autoantibody levels as a diagnostic biomarker. |
| [55,56,57,58,59,60,61,62] |
| Cutaneous Squamous Cell carcinoma (cSCC) | Protein overexpression in tumor tissues, which promotes cell proliferation and migration by EMT. | n.d. | n.d. | [63] |
| Gastric carcinoma | mRNA and protein upregulation are correlated with lymph node metastasis, depth of invasion, and poor prognosis. | Serum levels as a diagnostic biomarker for metastasis prediction. | Nuclear and cytoplasmic localization. | [64,65,66,67] |
| Glioma |
| n.d. | Nuclear localization. | [68,69,70,71,72,73] |
| Head and Neck Squamous Cell carcinoma (HNSCC) |
| In oSCC: serum levels are used as a diagnostic biomarker, with high levels correlated with late TNM stages, lymph node metastasis, and worse pathologic differentiation. |
| [74,75,76,77,78,79,80] |
| Liver cancer |
| Serum levels as a diagnostic biomarker. |
| [81,82,83,84,85,86] |
| Lung cancer |
| High post-treatment serum levels are associated with lower OS. |
| [55,87,88,89,90,91,92,93,94,95,96] |
| Melanoma | mRNA and protein overexpression are associated with vascular invasion, a high mitotic rate, lower response to therapy, and a poor prognosis. | n.d. | Nuclear localization. | [97,98,99,100] |
| Oesophageal carcinoma (EAC) | Protein overexpression in tumor tissues is associated with worse OS. | n.d. | Mainly nuclear localization. | [101,102,103,104,105,106] |
| Osteosarcoma |
| n.d. |
| [107,108,109,110,111,112,113] |
| Ovarian cancer | Protein overexpression in tumor tissues is associated with advanced stages, platinum resistance, poor chemosensitivity, decreased OS, and lymph node metastasis. | n.d. |
| [101,114,115,116,117,118,119,120,121] |
| Pancreatic adenocarcinoma (PDAC) |
| n.d. | Strong nuclear staining in tumor tissues, with cytosol staining only in advanced stages. | [55,87,101,122,123,124] |
| Prostate carcinoma (PCa) | Protein overexpression in cancer samples. | n.d. | Nuclear and cytoplasmic localization. | [125,126] |
| Salivary gland carcinoma |
| n.d. | Mainly nuclear staining, with nuclear and cytosolic staining in some malignant forms. | [127,128] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Malfatti, M.C.; Bellina, A.; Antoniali, G.; Tell, G. Revisiting Two Decades of Research Focused on Targeting APE1 for Cancer Therapy: The Pros and Cons. Cells 2023, 12, 1895. https://doi.org/10.3390/cells12141895
Malfatti MC, Bellina A, Antoniali G, Tell G. Revisiting Two Decades of Research Focused on Targeting APE1 for Cancer Therapy: The Pros and Cons. Cells. 2023; 12(14):1895. https://doi.org/10.3390/cells12141895
Chicago/Turabian StyleMalfatti, Matilde Clarissa, Alessia Bellina, Giulia Antoniali, and Gianluca Tell. 2023. "Revisiting Two Decades of Research Focused on Targeting APE1 for Cancer Therapy: The Pros and Cons" Cells 12, no. 14: 1895. https://doi.org/10.3390/cells12141895
APA StyleMalfatti, M. C., Bellina, A., Antoniali, G., & Tell, G. (2023). Revisiting Two Decades of Research Focused on Targeting APE1 for Cancer Therapy: The Pros and Cons. Cells, 12(14), 1895. https://doi.org/10.3390/cells12141895