Injured Achilles Tendons Treated with Adipose-Derived Stem Cells Transplantation and GDF-5

 , ,
, ,  , , ,
, , , 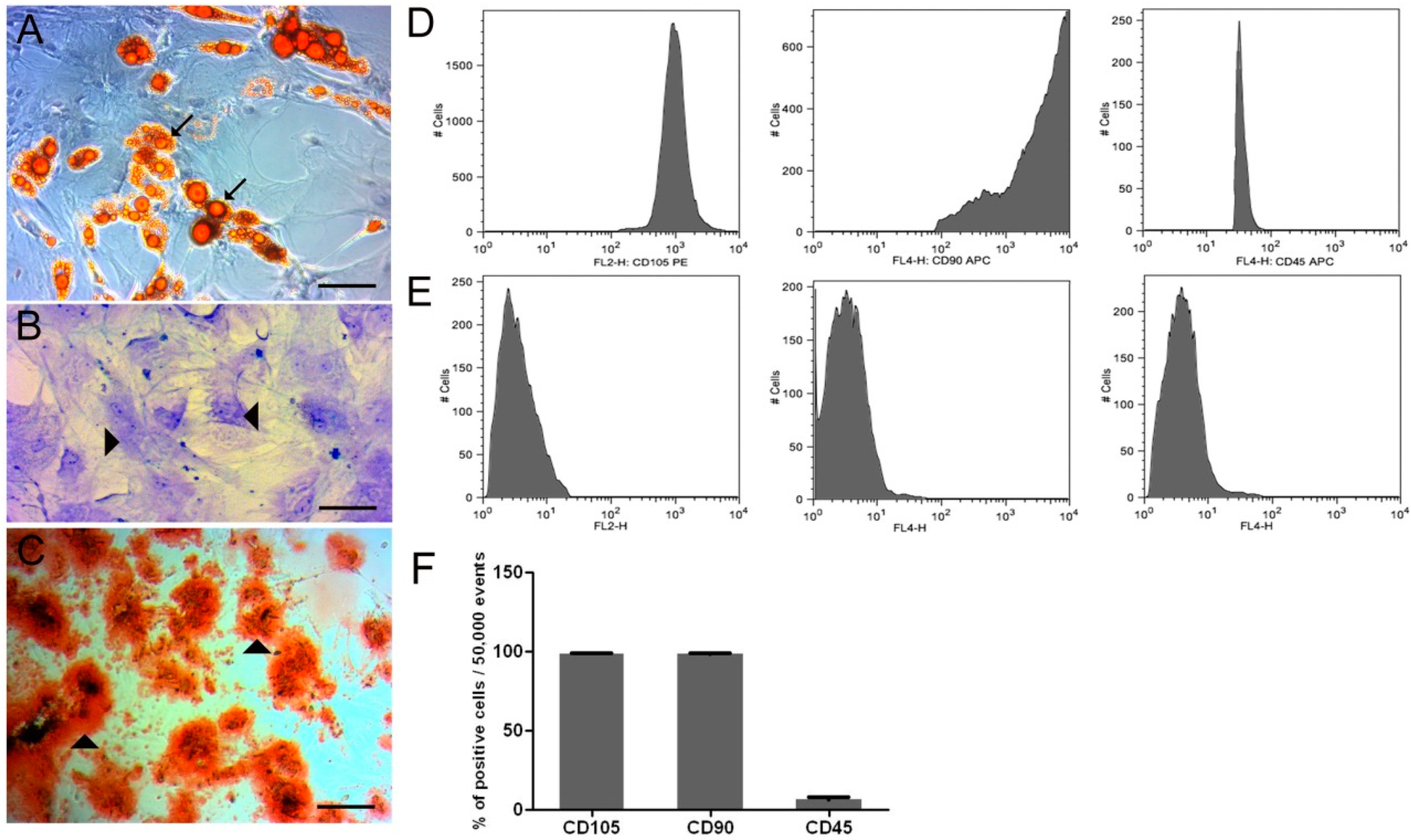{kind=link}
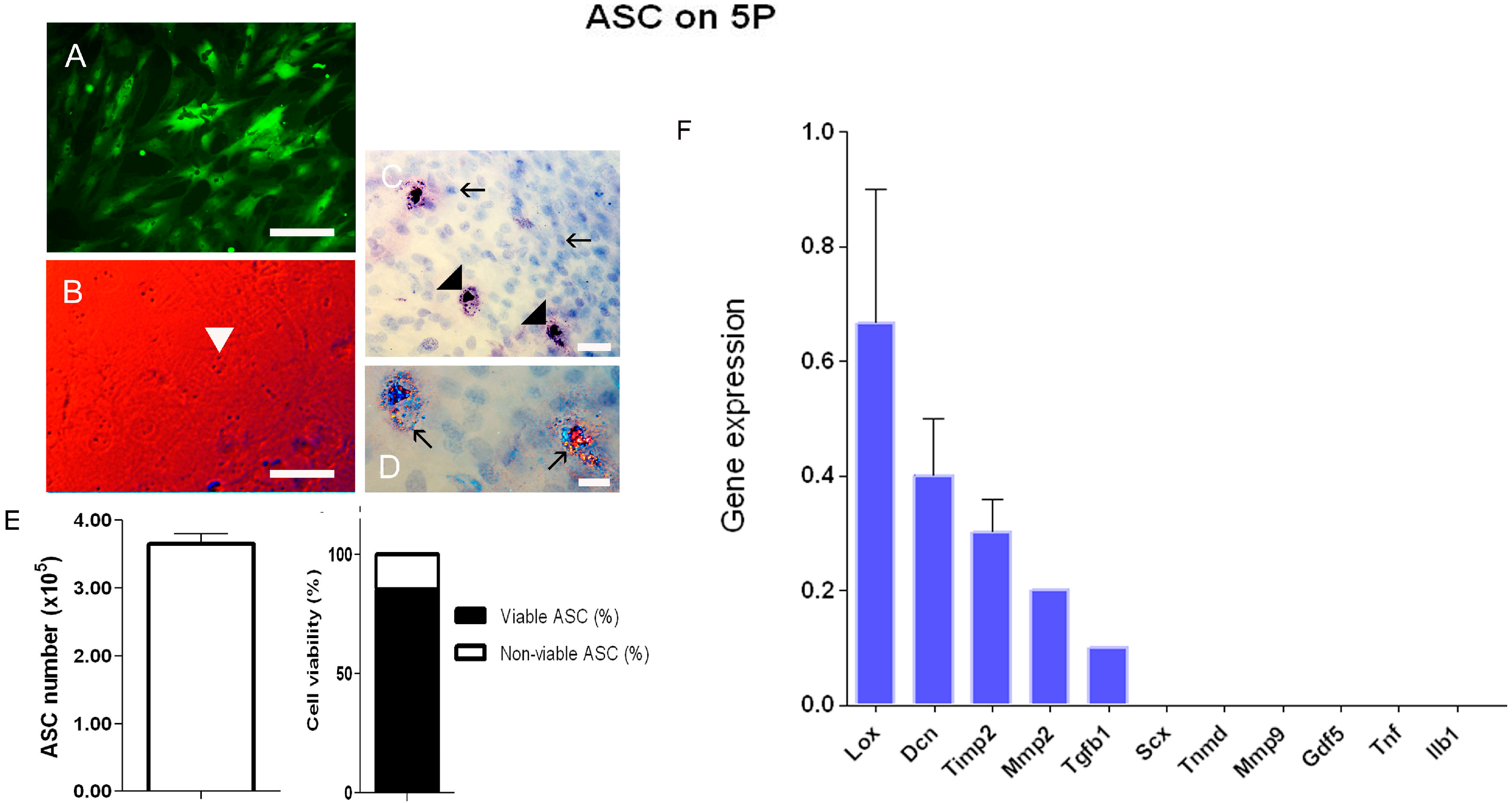{kind=link}
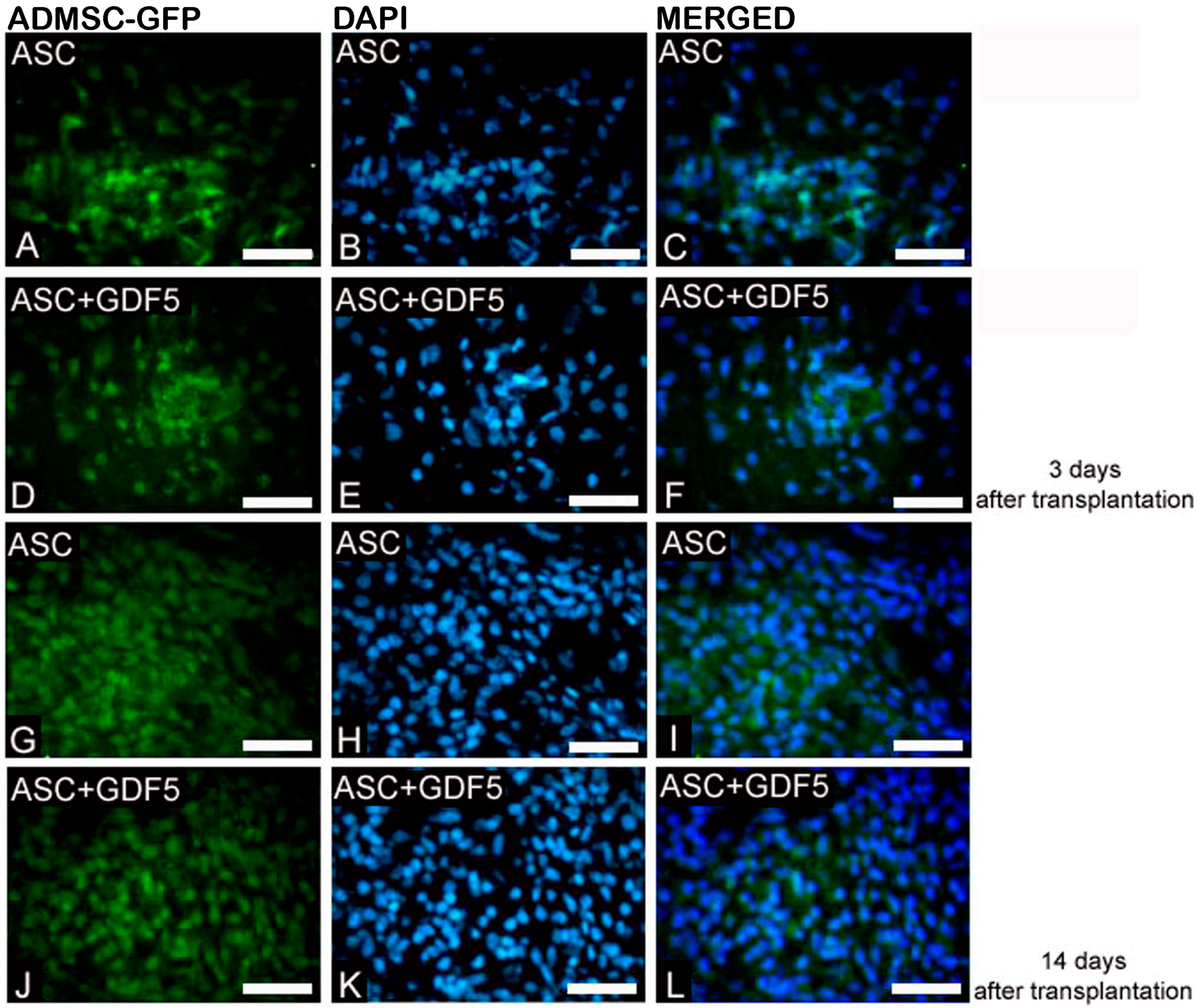{kind=link}
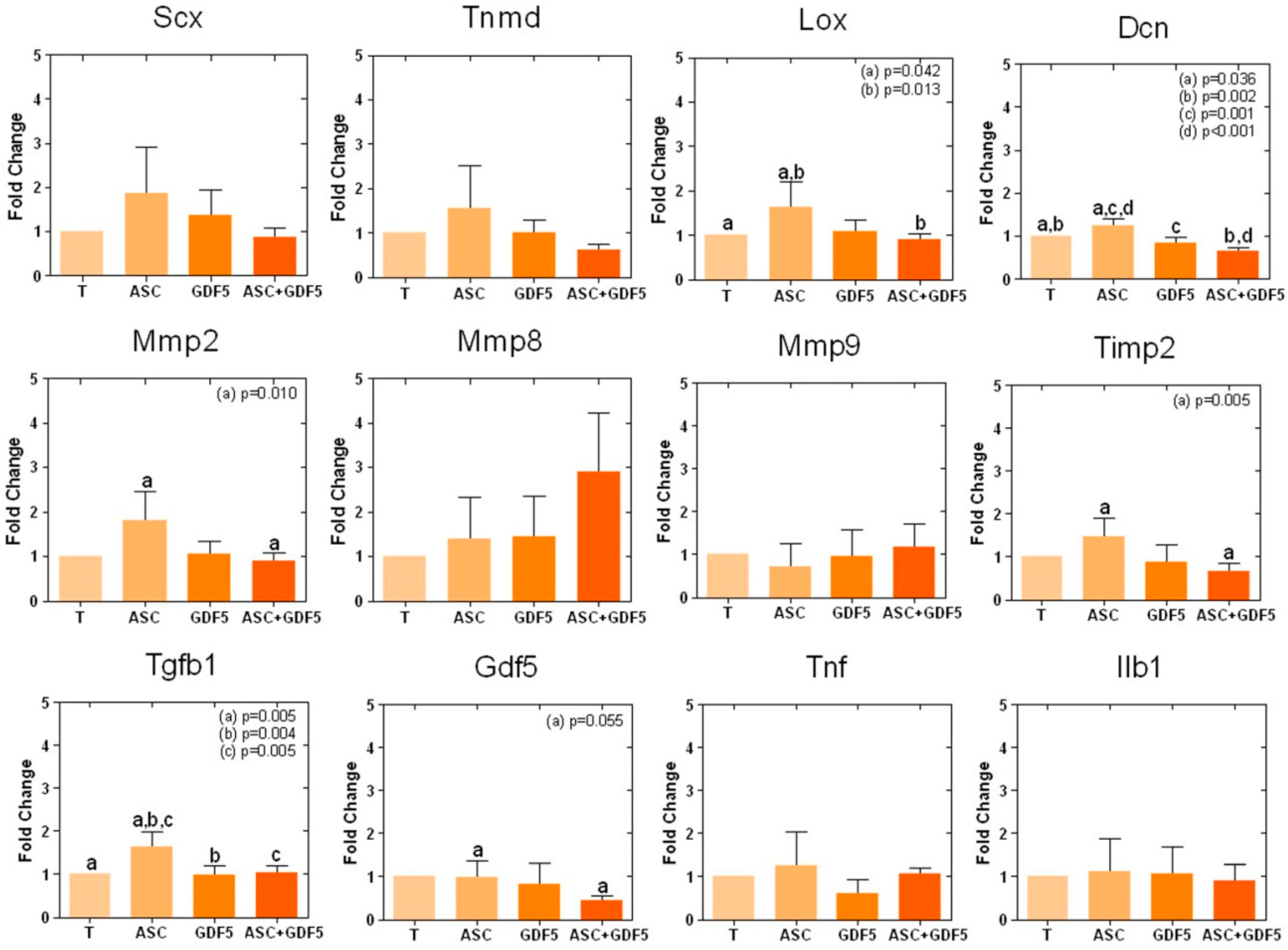{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. In Vitro Experiments
2.1.1. Isolation of ADMSC and Cell Culture
2.1.2. Flow Cytometry
2.1.3. In Vitro Differentiation Potential of ADMSC
2.1.4. Cell Viability
2.1.5. Contrast by Differential Interference (DIC)
2.1.6. Real Time-PCR Array for Analysis of ADMSC Gene Expression
2.2. In Vivo Experiments
2.2.1. Experimental Groups
2.2.2. Partial Transection of the Calcaneal Tendon and Application of ADMSC and GDF-5
2.2.3. Preparation of Sections in Freezing
2.2.4. DAPI Staining
2.2.5. Polarization Microscopy: Birefringence Measurements
2.2.6. Linear Dichroism Measurements
2.2.7. Real Time-PCR Array
2.2.8. Dosage of Hydroxyproline
2.2.9. Evaluation of the Max Contact Intensity of the Rat Paw after Partial Transection
2.2.10. Biomechanical Parameters
2.2.11. Statistical Analysis
3. Results
3.1. In Vitro
3.1.1. In Vitro Differentiation Potential of ADMSC and Flow Cytometry
3.1.2. Fluorescence, Birefringence, and Contrast by Differential Interference (DIC)
3.1.3. Cell Viability
3.1.4. Real-Time PCR Array
3.2. In Vivo
3.2.1. Immunofluorescence
3.2.2. Real-Time PCR Array
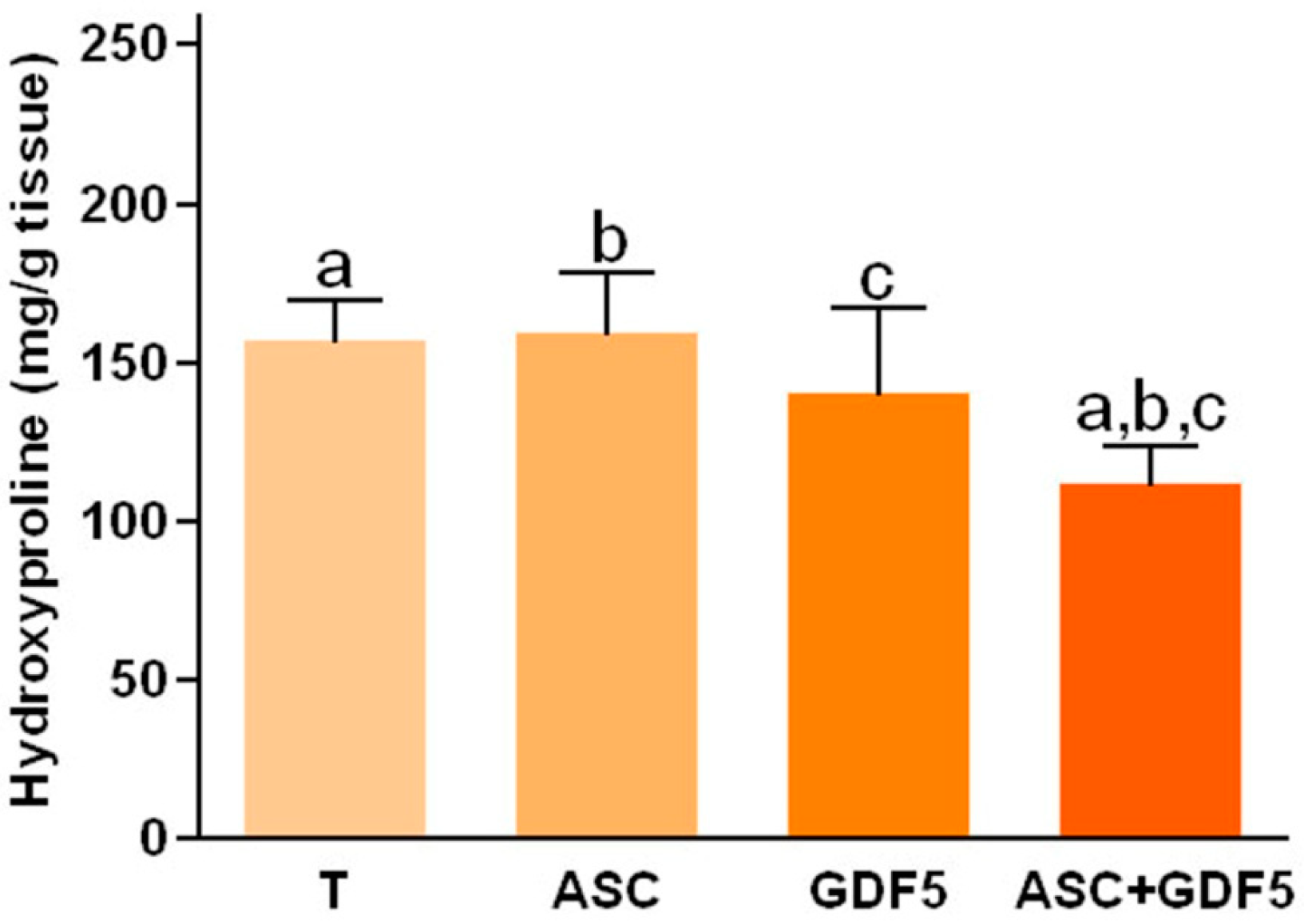
3.2.3. Dosage of Hydroxyproline
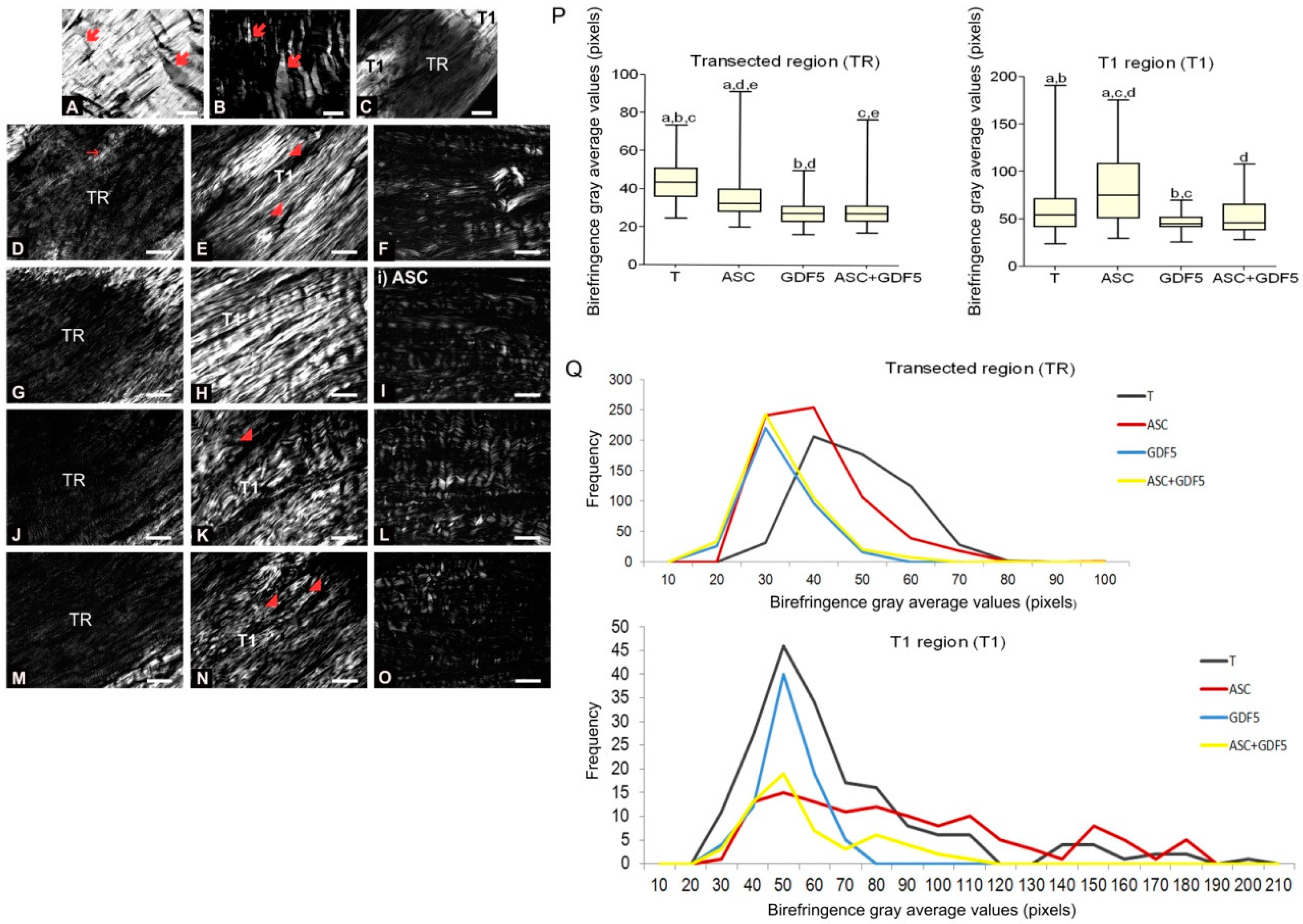
3.2.4. Birefringence Measurements
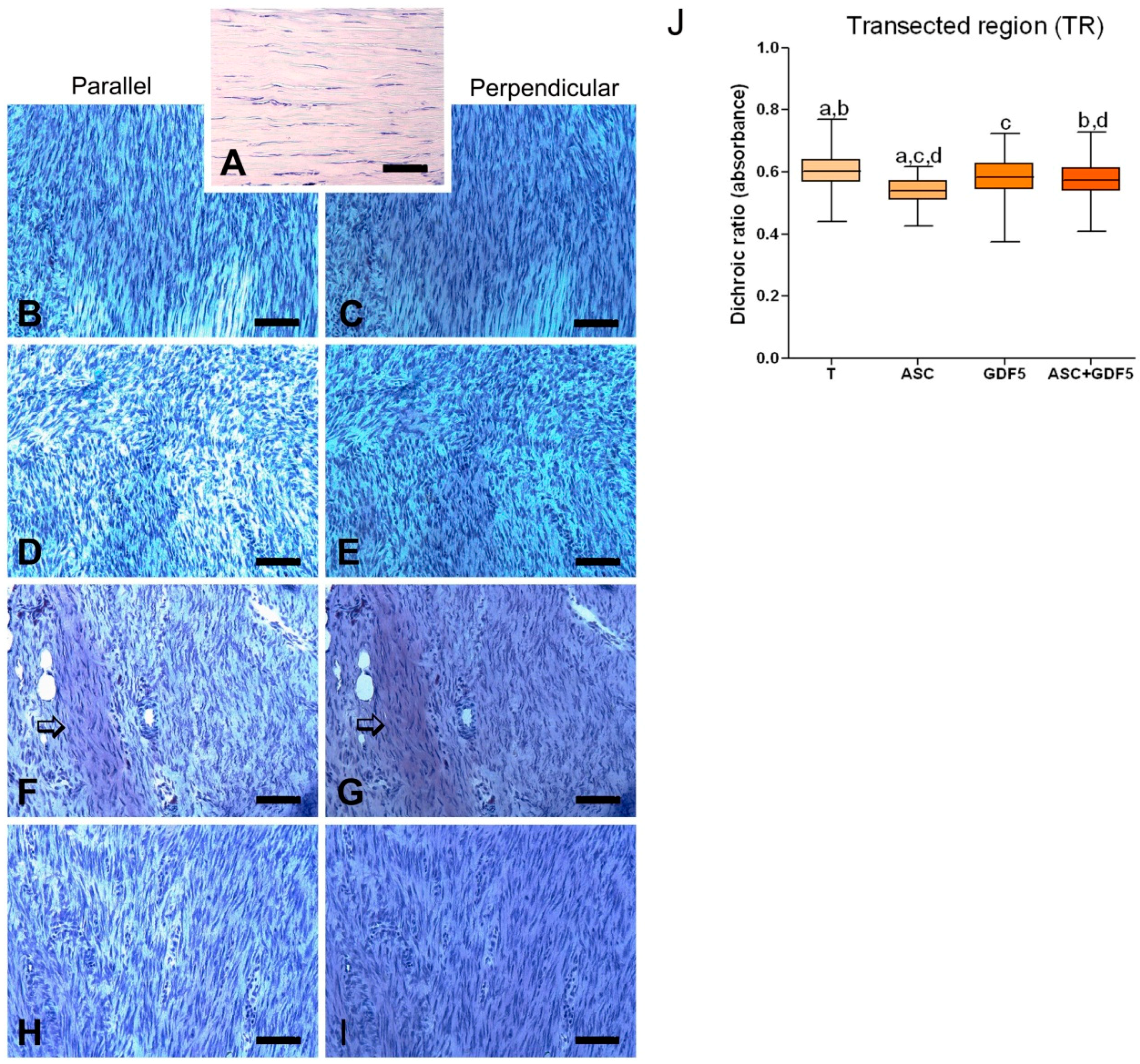
3.2.5. Linear Dichroism Measurements
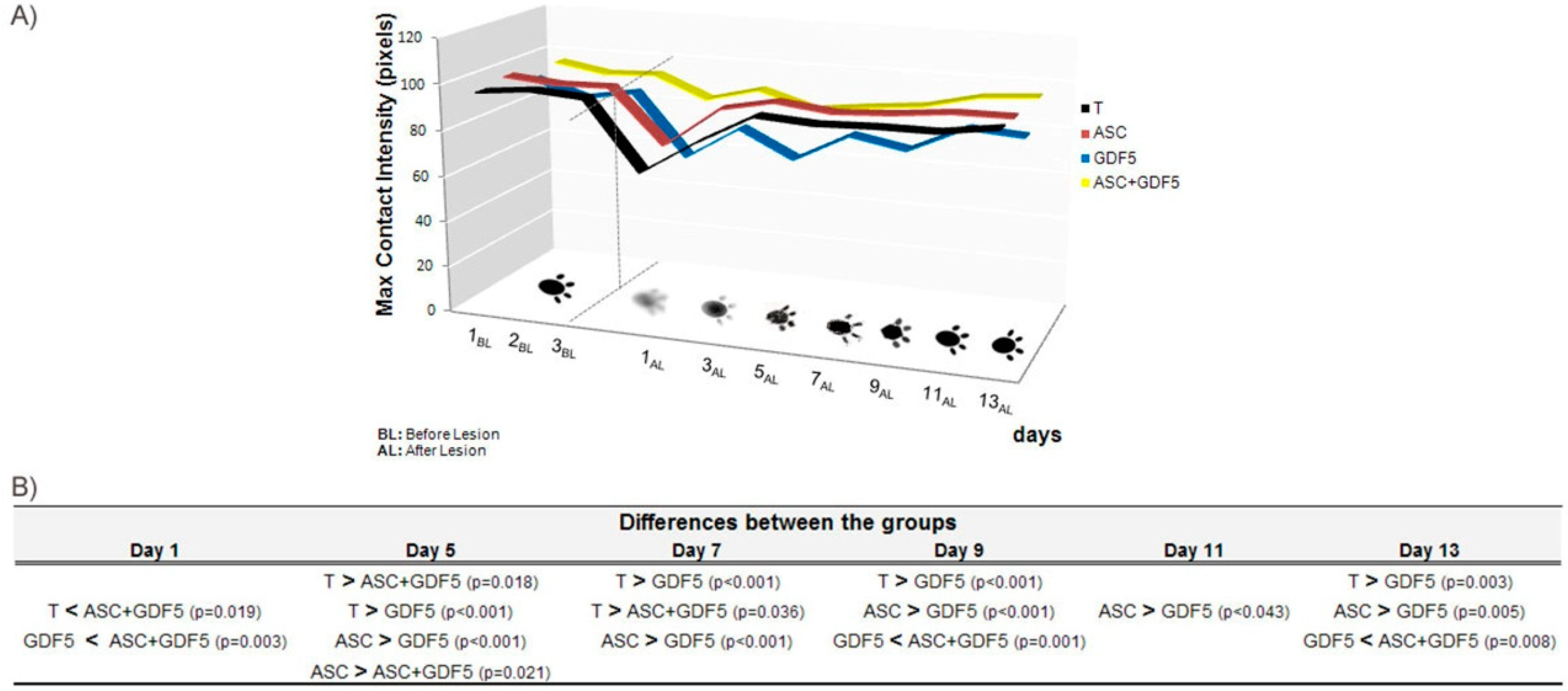
3.2.6. CatWalk System
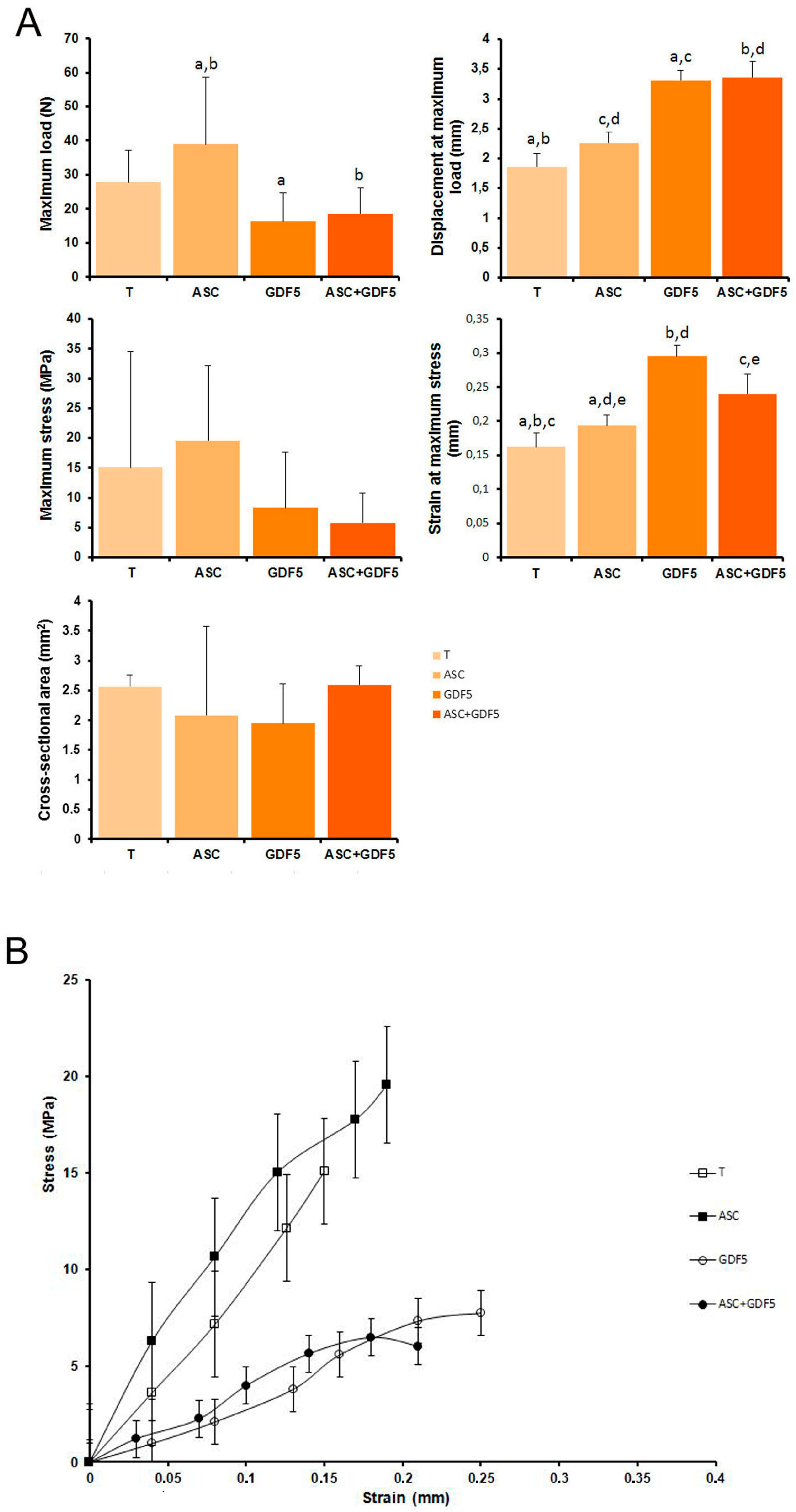
3.2.7. Biomechanical Parameters
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Goncalves, A.I.; Rodrigues, M.T.; Lee, S.J.; Atala, A.; Yoo, J.J.; Reis, R.L.; Gomes, M.E. Understanding the role of growth factors in modulating stem cell tenogenesis. PLoS ONE 2013, 8, e83734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zabrzynski, J.; Lapaj, L.; Paczesny, L.; Zabrzynska, A.; Grzanka, D. Tendon–function related structure, simple healing process and mysterious ageing. Folia Morphol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Vidal, B.C.; Mello, M.L. Proteoglycan arrangement in tendon collagen bundles. Cell Mol. Biol. 1984, 30, 195–204. [Google Scholar] [PubMed]
- De Campos Vidal, B.; Mello, M.L. Optical anisotropy of collagen fibers of rat calcaneal tendons: An approach to spatially resolved supramolecular organization. Acta Histochem. 2010, 112, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Marqueti, R.C.; Durigan, J.L.Q.; Oliveira, A.J.S.; Mekaro, M.S.; Guzzoni, V.; Aro, A.A.; Pimentel, E.R.; Selistre-de-Araujo, H.S. Effects of aging and resistance training in rat tendon remodeling. FASEB J. 2018, 32, 353–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Douglas, T.; Heinemann, S.; Bierbaum, S.; Scharnweber, D.; Worch, H. Fibrillogenesis of collagen types I, II, and III with small leucine-rich proteoglycans decorin and biglycan. Biomacromolecules 2006, 7, 2388–2393. [Google Scholar] [CrossRef] [PubMed]
- Uysal, C.A.; Tobita, M.; Hyakusoku, H.; Mizuno, H. Adipose-derived stem cells enhance primary tendon repair: Biomechanical and immunohistochemical evaluation. J. Plast. Reconstr. Aesthet. Surg. 2012, 65, 1712–1719. [Google Scholar] [CrossRef] [PubMed]
- Oshiro, W.; Lou, J.; Xing, X.; Tu, Y.; Manske, P.R. Flexor tendon healing in the rat: A histologic and gene expression study. J. Hand Surg. Am. 2003, 28, 814–823. [Google Scholar] [CrossRef]
- Tomiosso, T.C.; Nakagaki, W.R.; Gomes, L.; Hyslop, S.; Pimentel, E.R. Organization of collagen bundles during tendon healing in rats treated with L-NAME. Cell Tissue Res. 2009, 337, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Christiansen, D.L.; Huang, E.K.; Silver, F.H. Assembly of type I collagen: Fusion of fibril subunits and the influence of fibril diameter on mechanical properties. Matrix Biol. 2000, 19, 409–420. [Google Scholar] [CrossRef]
- Almekinders, L.C.; Almekinders, S.V. Outcome in the treatment of chronic overuse sports injuries: A retrospective study. J. Orthop. Sports Phys. Ther. 1994, 19, 157–161. [Google Scholar] [CrossRef] [PubMed]
- Favata, M.; Beredjiklian, P.K.; Zgonis, M.H.; Beason, D.P.; Crombleholme, T.M.; Jawad, A.F.; Soslowsky, L.J. Regenerative properties of fetal sheep tendon are not adversely affected by transplantation into an adult environment. J. Orthop. Res. 2006, 24, 2124–2132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ni, M.; Lui, P.P.; Rui, Y.F.; Lee, Y.W.; Lee, Y.W.; Tan, Q.; Wong, Y.M.; Kong, S.K.; Lau, P.M.; Li, G.; et al. Tendon-derived stem cells (TDSCs) promote tendon repair in a rat patellar tendon window defect model. J. Orthop. Res. 2012, 30, 613–619. [Google Scholar] [CrossRef] [PubMed]
- Pittenger, M.F.; Mackay, A.M.; Beck, S.C.; Jaiswal, R.K.; Douglas, R.; Mosca, J.D.; Moorman, M.A.; Simonetti, D.W.; Craig, S.; Marshak, D.R. Multilineage potential of adult human mesenchymal stem cells. Science 1999, 284, 143–147. [Google Scholar] [CrossRef] [PubMed]
- Kern, S.; Eichler, H.; Stoeve, J.; Kluter, H.; Bieback, K. Comparative analysis of mesenchymal stem cells from bone marrow, umbilical cord blood, or adipose tissue. Stem Cells 2006, 24, 1294–1301. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, N.; Kushida, T.; Oe, K.; Umeda, M.; Ikehara, S.; Iida, H. Treating Achilles tendon rupture in rats with bone-marrow-cell transplantation therapy. J. Bone Joint Surg. Am. 2010, 92, 2776–2784. [Google Scholar] [CrossRef] [PubMed]
- Schneider, M.; Angele, P.; Jarvinen, T.A.H.; Docheva, D. Rescue plan for Achilles: Therapeutics steering the fate and functions of stem cells in tendon wound healing. Adv. Drug Deliv. Rev. 2017, 129, 352–375. [Google Scholar] [CrossRef] [PubMed]
- Gimble, J.M.; Katz, A.J.; Bunnell, B.A. Adipose-derived stem cells for regenerative medicine. Circ. Res. 2007, 100, 1249–1260. [Google Scholar] [CrossRef] [PubMed]
- Park, A.; Hogan, M.V.; Kesturu, G.S.; James, R.; Balian, G.; Chhabra, A.B. Adipose-derived mesenchymal stem cells treated with growth differentiation factor-5 express tendon-specific markers. Tissue Eng. Part A 2010, 16, 2941–2951. [Google Scholar] [CrossRef] [PubMed]
- Schneider, P.R.; Buhrmann, C.; Mobasheri, A.; Matis, U.; Shakibaei, M. Three-dimensional high-density co-culture with primary tenocytes induces tenogenic differentiation in mesenchymal stem cells. J. Orthop. Res. 2011, 29, 1351–1360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pacini, S.; Spinabella, S.; Trombi, L.; Fazzi, R.; Galimberti, S.; Dini, F.; Carlucci, F.; Petrini, M. Suspension of bone marrow-derived undifferentiated mesenchymal stromal cells for repair of superficial digital flexor tendon in race horses. Tissue Eng. 2007, 13, 2949–2955. [Google Scholar] [CrossRef] [PubMed]
- Vieira, M.H.; Oliveira, R.J.; Eca, L.P.; Pereira, I.S.; Hermeto, L.C.; Matuo, R.; Fernandes, W.S.; Silva, R.A.; Antoniolli, A.C. Therapeutic potential of mesenchymal stem cells to treat Achilles tendon injuries. Genet. Mol. Res. 2014, 13, 10434–10449. [Google Scholar] [CrossRef] [PubMed]
- Valencia Mora, M.; Antuna Antuna, S.; Garcia Arranz, M.; Carrascal, M.T.; Barco, R. Application of adipose tissue-derived stem cells in a rat rotator cuff repair model. Injury 2014, 45 (Suppl. 4), S22–S27. [Google Scholar] [CrossRef]
- Guilak, F.; Cohen, D.M.; Estes, B.T.; Gimble, J.M.; Liedtke, W.; Chen, C.S. Control of stem cell fate by physical interactions with the extracellular matrix. Cell Stem Cell 2009, 5, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Storm, E.E.; Huynh, T.V.; Copeland, N.G.; Jenkins, N.A.; Kingsley, D.M.; Lee, S.J. Limb alterations in brachypodism mice due to mutations in a new member of the TGF beta-superfamily. Nature 1994, 368, 639–643. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.C.; Hoang, B.; Thomas, J.T.; Vukicevic, S.; Luyten, F.P.; Ryba, N.J.; Kozak, C.A.; Reddi, A.H.; Moos, M., Jr. Cartilage-derived morphogenetic proteins. New members of the transforming growth factor-beta superfamily predominantly expressed in long bones during human embryonic development. J. Biol. Chem. 1994, 269, 28227–28234. [Google Scholar] [PubMed]
- Zhou, S.; Yates, K.E.; Eid, K.; Glowacki, J. Demineralized bone promotes chondrocyte or osteoblast differentiation of human marrow stromal cells cultured in collagen sponges. Cell Tissue Bank. 2005, 6, 33–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolfman, N.M.; Hattersley, G.; Cox, K.; Celeste, A.J.; Nelson, R.; Yamaji, N.; Dube, J.L.; DiBlasio-Smith, E.; Nove, J.; Song, J.J. Ectopic induction of tendon and ligament in rats by growth and differentiation factors 5, 6, and 7, members of the TGF-beta gene family. J. Clin. Investig. 1997, 100, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Forslund, C.; Aspenberg, P. Tendon healing stimulated by injected CDMP-2. Med. Sci. Sports Exerc. 2001, 33, 685–687. [Google Scholar] [CrossRef] [PubMed]
- Forslund, C.; Rueger, D.; Aspenberg, P. A comparative dose-response study of cartilage-derived morphogenetic protein (CDMP)-1, -2 and -3 for tendon healing in rats. J. Orthop. Res. 2003, 21, 617–621. [Google Scholar] [CrossRef]
- Chhabra, A.; Tsou, D.; Clark, R.T.; Gaschen, V.; Hunziker, E.B.; Mikic, B. GDF-5 deficiency in mice delays Achilles tendon healing. J. Orthop. Res. 2003, 21, 826–835. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.F.; He, X.; He, J.; Zhang, L.H.; Su, X.J.; Dong, Z.Y.; Xu, Y.J.; Li, Y.; Li, Y.L. High efficient isolation and systematic identification of human adipose-derived mesenchymal stem cells. J. Biomed. Sci. 2011, 18, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vidal, B.C. Form birefringence as applied to biopolymer and inorganic material supraorganization. Biotech. Histochem. 2010, 85, 365–378. [Google Scholar] [CrossRef] [PubMed]
- Vidal, B.C.; Dos Anjos, E.H.; Mello, M.L. Optical anisotropy reveals molecular order in a mouse enthesis. Cell Tissue Res. 2015, 362, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar]
- Aro, A.A.; Freitas, K.M.; Foglio, M.A.; Carvalho, J.E.; Dolder, H.; Gomes, L.; Vidal, B.C.; Pimentel, E.R. Effect of the Arrabidaea chica extract on collagen fiber organization during healing of partially transected tendon. Life Sci. 2013, 92, 799–807. [Google Scholar] [CrossRef] [PubMed]
- Aro, A.A.; Perez, M.O.; Vieira, C.P.; Esquisatto, M.A.; Rodrigues, R.A.; Gomes, L.; Pimentel, E.R. Effect of Calendula officinalis cream on achilles tendon healing. Anat. Rec. 2015, 298, 428–435. [Google Scholar] [CrossRef] [PubMed]
- Vidal, B.d.C. Evaluation of the carbohydrate role in the molecular order of collagen bundles: Microphotometric measurements of textural birefringence. Cell. Mol. Biol. 1986, 32, 527–535. [Google Scholar]
- Feitosa, V.; Vidal, B.C.; Pimentel, E.R. Optical anisotropy of a pig tendon under compression. J. Anat. 2002, 200, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Mello, M.L.; de Campos Vidal, B. Experimental tendon repair: Glycosaminoglycan arrangement in newly synthesized collagen fibers. Cell. Mol. Biol. (Noisy-le-grand) 2003, 49, 579–585. [Google Scholar]
- Stegemann, H.; Stalder, K. Determination of hydroxyproline. Clin. Chim. Acta 1967, 18, 267–273. [Google Scholar] [CrossRef]
- Biancalana, A.; Veloso, L.A.; Gomes, L. Obesity affects collagen fibril diameter and mechanical properties of tendons in Zucker rats. Connect. Tissue Res. 2010, 51, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Sparrow, K.J.; Finucane, S.D.; Owen, J.R.; Wayne, J.S. The effects of low-intensity ultrasound on medial collateral ligament healing in the rabbit model. Am. J. Sports Med. 2005, 33, 1048–1056. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Liang, L.; Zong, C.; Lai, F.; Zhu, P.; Liu, Y.; Jiang, J.; Yang, Y.; Gao, L.; Ye, F.; et al. Kupffer cells-dependent inflammation in the injured liver increases recruitment of mesenchymal stem cells in aging mice. Oncotarget 2016, 7, 1084–1095. [Google Scholar] [CrossRef] [PubMed]
- Heissig, B.; Dhahri, D.; Eiamboonsert, S.; Salama, Y.; Shimazu, H.; Munakata, S.; Hattori, K. Role of mesenchymal stem cell-derived fibrinolytic factor in tissue regeneration and cancer progression. Cell. Mol. Life Sci. 2015, 72, 4759–4770. [Google Scholar] [CrossRef] [PubMed]
- Parry, D.A. The molecular and fibrillar structure of collagen and its relationship to the mechanical properties of connective tissue. Biophys. Chem. 1988, 29, 195–209. [Google Scholar] [CrossRef]
- Birk, D.E.; Mayne, R. Localization of collagen types I, III and V during tendon development. Changes in collagen types I and III are correlated with changes in fibril diameter. Eur. J. Cell Biol. 1997, 72, 352–361. [Google Scholar] [PubMed]
- Oryan, A.; Moshiri, A. A long term study on the role of exogenous human recombinant basic fibroblast growth factor on the superficial digital flexor tendon healing in rabbits. J. Musculoskelet. Neuronal Interact. 2011, 11, 185–195. [Google Scholar] [PubMed]
- Del Bue, M.; Ricco, S.; Ramoni, R.; Conti, V.; Gnudi, G.; Grolli, S. Equine adipose-tissue derived mesenchymal stem cells and platelet concentrates: Their association in vitro and in vivo. Vet. Res. Commun. 2008, 32 (Suppl. 1), S51–S55. [Google Scholar] [CrossRef] [PubMed]
- Uysal, A.C.; Mizuno, H. Differentiation of adipose-derived stem cells for tendon repair. Methods Mol. Biol. 2011, 702, 443–451. [Google Scholar] [PubMed]
- Bolt, P.; Clerk, A.N.; Luu, H.H.; Kang, Q.; Kummer, J.L.; Deng, Z.L.; Olson, K.; Primus, F.; Montag, A.G.; He, T.C. BMP-14 gene therapy increases tendon tensile strength in a rat model of Achilles tendon injury. J. Bone Joint Surg. Am. 2007, 89, 1315–1320. [Google Scholar] [CrossRef] [PubMed]
- Mikic, B.; Schalet, B.J.; Clark, R.T.; Gaschen, V.; Hunziker, E.B. GDF-5 deficiency in mice alters the ultrastructure, mechanical properties and composition of the Achilles tendon. J. Orthop. Res. 2001, 19, 365–371. [Google Scholar] [CrossRef] [Green Version]
- Rickert, M.; Wang, H.; Wieloch, P.; Lorenz, H.; Steck, E.; Sabo, D.; Richter, W. Adenovirus-mediated gene transfer of growth and differentiation factor-5 into tenocytes and the healing rat Achilles tendon. Connect. Tissue Res. 2005, 46, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Chan, B.P.; Chan, K.M.; Maffulli, N.; Webb, S.; Lee, K.K. Effect of basic fibroblast growth factor. An in vitro study of tendon healing. Clin. Orthop. Relat. Res. 1997, 239–247. [Google Scholar] [CrossRef]
- McGeachie, J.; Tennant, M. Growth factors and their implications for clinicians: A brief review. Aust. Dent. J. 1997, 42, 375–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, P.; Maffulli, N. Tendon injury and tendinopathy: Healing and repair. J. Bone Joint Surg. Am. 2005, 87, 187–202. [Google Scholar] [PubMed]
- Sheng, J.; Zeng, B.; Jiang, P. Effects of exogenous basic fibroblast growth factor on in-sheathed tendon healing and adhesion formation. Zhongguo xiu fu chong jian wai ke za zhi Chin. J. Reparative Reconstr. Surg. 2007, 21, 733–737. [Google Scholar]
- Gospodarowicz, D.; Ferrara, N.; Schweigerer, L.; Neufeld, G. Structural characterization and biological functions of fibroblast growth factor. Endocr. Rev. 1987, 8, 95–114. [Google Scholar] [CrossRef] [PubMed]
- McNeil, P.L.; Muthukrishnan, L.; Warder, E.; D’Amore, P.A. Growth factors are released by mechanically wounded endothelial cells. J. Cell Biol. 1989, 109, 811–822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kannus, P. Structure of the tendon connective tissue. Scand. J. Med. Sci. Sports 2000, 10, 312–320. [Google Scholar] [CrossRef] [PubMed]
- Wurgler-Hauri, C.C.; Dourte, L.M.; Baradet, T.C.; Williams, G.R.; Soslowsky, L.J. Temporal expression of 8 growth factors in tendon-to-bone healing in a rat supraspinatus model. J. Shoulder Elb. Surg. 2007, 16, S198–S203. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Ding, Y.; Chen, J.; Liu, D.; Ding, M.; Zhang, Y. Treatment of corneal epithelial wounds in dogs using basic fibroblast growth factor. Vet. Med. (Praha) 2009, 54, 280–286. [Google Scholar] [CrossRef] [Green Version]
- Dunkman, A.A.; Buckley, M.R.; Mienaltowski, M.J.; Adams, S.M.; Thomas, S.J.; Satchell, L.; Kumar, A.; Pathmanathan, L.; Beason, D.P.; Iozzo, R.V.; et al. The tendon injury response is influenced by decorin and biglycan. Ann. Biomed. Eng. 2013, 42, 619–630. [Google Scholar] [CrossRef] [PubMed]
- Herchenhan, A.; Uhlenbrock, F.; Eliasson, P.; Weis, M.; Eyre, D.; Kadler, K.E.; Magnusson, S.P.; Kjaer, M. Lysyl Oxidase Activity Is Required for Ordered Collagen Fibrillogenesis by Tendon Cells. J. Biol. Chem. 2015, 290, 16440–16450. [Google Scholar] [CrossRef] [PubMed]
- Aro, A.A.; Nishan, U.; Perez, M.O.; Rodrigues, R.A.; Foglio, M.A.; Carvalho, J.E.; Gomes, L.; Vidal, B.C.; Pimentel, E.R. Structural and biochemical alterations during the healing process of tendons treated with Aloe vera. Life Sci. 2012, 91, 885–893. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.; An, J.; Kim, J.H.; Kim, E.S.; Kim, S.H.; Cho, Y.K.; Cha, D.H.; Han, M.Y.; Lee, K.H.; Sheen, Y.H. Low levels of tissue inhibitor of metalloproteinase-2 at birth may be associated with subsequent development of bronchopulmonary dysplasia in preterm infants. Korean J. Pediatr. 2015, 58, 415–420. [Google Scholar] [CrossRef] [PubMed]
- Van Buul, G.M.; Villafuertes, E.; Bos, P.K.; Waarsing, J.H.; Kops, N.; Narcisi, R.; Weinans, H.; Verhaar, J.A.; Bernsen, M.R.; van Osch, G.J. Mesenchymal stem cells secrete factors that inhibit inflammatory processes in short-term osteoarthritic synovium and cartilage explant culture. Osteoarthritis Cartil. 2012, 20, 1186–1196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manferdini, C.; Maumus, M.; Gabusi, E.; Piacentini, A.; Filardo, G.; Peyrafitte, J.A.; Jorgensen, C.; Bourin, P.; Fleury-Cappellesso, S.; Facchini, A. Adipose-derived mesenchymal stem cells exert antiinflammatory effects on chondrocytes and synoviocytes from osteoarthritis patients through prostaglandin E2. Arthritis Rheum. 2013, 65, 1271–1281. [Google Scholar] [CrossRef] [PubMed]
- Molloy, T.; Wang, Y.; Murrell, G. The roles of growth factors in tendon and ligament healing. Sports Med. 2003, 33, 381–394. [Google Scholar] [CrossRef] [PubMed]
- Fu, S.C.; Wong, Y.P.; Cheuk, Y.C.; Lee, K.M.; Chan, K.M. TGF-beta1 reverses the effects of matrix anchorage on the gene expression of decorin and procollagen type I in tendon fibroblasts. Clin. Orthop. Res. 2005, 226–232. [Google Scholar] [CrossRef]
- Mehta, V.; Mass, D. The use of growth factors on tendon injuries. J. Hand Ther. 2005, 18, 87–92; quiz 93. [Google Scholar] [CrossRef] [PubMed]
- Finnson, K.W.; McLean, S.; Di Guglielmo, G.M.; Philip, A. Dynamics of Transforming Growth Factor Beta Signaling in Wound Healing and Scarring. Adv. Wound Care 2013, 2, 195–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schweitzer, R.; Chyung, J.H.; Murtaugh, L.C.; Brent, A.E.; Rosen, V.; Olson, E.N.; Lassar, A.; Tabin, C.J. Analysis of the tendon cell fate using Scleraxis, a specific marker for tendons and ligaments. Development 2001, 128, 3855–3866. [Google Scholar] [PubMed]
- Holladay, C.; Abbah, S.A.; O’Dowd, C.; Pandit, A.; Zeugolis, D.I. Preferential tendon stem cell response to growth factor supplementation. J. Tissue Eng. Regen. Med. 2016, 10, 783–798. [Google Scholar] [CrossRef] [PubMed]
- Shukunami, C.; Takimoto, A.; Oro, M.; Hiraki, Y. Scleraxis positively regulates the expression of tenomodulin, a differentiation marker of tenocytes. Dev. Biol. 2006, 298, 234–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brandau, O.; Meindl, A.; Fassler, R.; Aszodi, A. A novel gene, tendin, is strongly expressed in tendons and ligaments and shows high homology with chondromodulin-I. Dev. Dyn. 2001, 221, 72–80. [Google Scholar] [CrossRef] [PubMed]
- Shukunami, C.; Oshima, Y.; Hiraki, Y. Molecular cloning of tenomodulin, a novel chondromodulin-I related gene. Biochem. Biophy. Res. Commun. 2001, 280, 1323–1327. [Google Scholar] [CrossRef] [PubMed]
- Yamana, K.; Wada, H.; Takahashi, Y.; Sato, H.; Kasahara, Y.; Kiyoki, M. Molecular cloning and characterization of CHM1L, a novel membrane molecule similar to chondromodulin-I. Biochem. Biophy. Res. Commun. 2001, 280, 1101–1106. [Google Scholar] [CrossRef] [PubMed]
- Oshima, Y.; Shukunami, C.; Honda, J.; Nishida, K.; Tashiro, F.; Miyazaki, J. Chondromodulin-I-related angiogenesis inhibitor, in mouse eyes. Investig. Ophthalmol. Vis. Sci. 2003, 44, 1814–1823. [Google Scholar] [CrossRef]
- Pisani, D.F.; Pierson, P.M.; Massoudi, A.; Leclerc, L.; Chopard, A.; Marini, J.F.; Dechesne, C.A. Myodulin is a novel potential angiogenic factor in skeletal muscle. Exp. Cell Res. 2004, 292, 40–50. [Google Scholar] [CrossRef] [PubMed]





 ); observe the crimp (B) by positioning the largest axis of the tendon parallel to one of the polarizers: The same region observed in (A). (C) Panoramic image of the transected tendon for identification of the transection region (TR) and the proximal and distal transition region (T1). Groups T (D–F), ASC (G–I), GDF5 (J–L), and ASC+GDF5 (M–O). Observe the complete disorganization of collagen fibers in TR. The TR from different groups (D,G,J,M): Observe freshly formed collagen fibrils and an overlapping (↘) of this region with the thicker fibers present in T1. T1 (E,H,K,N): Collagen fibers with a greater organization in relation to TR, however, with fragmentation presence (◢) mainly in groups T (E), GDF5 (K), and ASC+GDF5 (N). Crimp (F,I,L,O) from the collagen fibers observed on T1: Observe similar undulation patterns of the collagen fibers between the groups, represented by light and dark regions. The largest axis of the tendon was positioned at 45° in relation to the crossed polarizers as parallel to one of the polarizers (B,F,I,L,O). (P) TR birefringence measurements in T1: Same letter represents significant differences between groups (p ≤ 0.05). (Q) Histogram of the frequency and birefringence values showing differences in the distribution of values in the different groups. Bars = 100 μm and 200 μm (a).
); observe the crimp (B) by positioning the largest axis of the tendon parallel to one of the polarizers: The same region observed in (A). (C) Panoramic image of the transected tendon for identification of the transection region (TR) and the proximal and distal transition region (T1). Groups T (D–F), ASC (G–I), GDF5 (J–L), and ASC+GDF5 (M–O). Observe the complete disorganization of collagen fibers in TR. The TR from different groups (D,G,J,M): Observe freshly formed collagen fibrils and an overlapping (↘) of this region with the thicker fibers present in T1. T1 (E,H,K,N): Collagen fibers with a greater organization in relation to TR, however, with fragmentation presence (◢) mainly in groups T (E), GDF5 (K), and ASC+GDF5 (N). Crimp (F,I,L,O) from the collagen fibers observed on T1: Observe similar undulation patterns of the collagen fibers between the groups, represented by light and dark regions. The largest axis of the tendon was positioned at 45° in relation to the crossed polarizers as parallel to one of the polarizers (B,F,I,L,O). (P) TR birefringence measurements in T1: Same letter represents significant differences between groups (p ≤ 0.05). (Q) Histogram of the frequency and birefringence values showing differences in the distribution of values in the different groups. Bars = 100 μm and 200 μm (a).
); observe the crimp (B) by positioning the largest axis of the tendon parallel to one of the polarizers: The same region observed in (A). (C) Panoramic image of the transected tendon for identification of the transection region (TR) and the proximal and distal transition region (T1). Groups T (D–F), ASC (G–I), GDF5 (J–L), and ASC+GDF5 (M–O). Observe the complete disorganization of collagen fibers in TR. The TR from different groups (D,G,J,M): Observe freshly formed collagen fibrils and an overlapping (↘) of this region with the thicker fibers present in T1. T1 (E,H,K,N): Collagen fibers with a greater organization in relation to TR, however, with fragmentation presence (◢) mainly in groups T (E), GDF5 (K), and ASC+GDF5 (N). Crimp (F,I,L,O) from the collagen fibers observed on T1: Observe similar undulation patterns of the collagen fibers between the groups, represented by light and dark regions. The largest axis of the tendon was positioned at 45° in relation to the crossed polarizers as parallel to one of the polarizers (B,F,I,L,O). (P) TR birefringence measurements in T1: Same letter represents significant differences between groups (p ≤ 0.05). (Q) Histogram of the frequency and birefringence values showing differences in the distribution of values in the different groups. Bars = 100 μm and 200 μm (a).
); observe the crimp (B) by positioning the largest axis of the tendon parallel to one of the polarizers: The same region observed in (A). (C) Panoramic image of the transected tendon for identification of the transection region (TR) and the proximal and distal transition region (T1). Groups T (D–F), ASC (G–I), GDF5 (J–L), and ASC+GDF5 (M–O). Observe the complete disorganization of collagen fibers in TR. The TR from different groups (D,G,J,M): Observe freshly formed collagen fibrils and an overlapping (↘) of this region with the thicker fibers present in T1. T1 (E,H,K,N): Collagen fibers with a greater organization in relation to TR, however, with fragmentation presence (◢) mainly in groups T (E), GDF5 (K), and ASC+GDF5 (N). Crimp (F,I,L,O) from the collagen fibers observed on T1: Observe similar undulation patterns of the collagen fibers between the groups, represented by light and dark regions. The largest axis of the tendon was positioned at 45° in relation to the crossed polarizers as parallel to one of the polarizers (B,F,I,L,O). (P) TR birefringence measurements in T1: Same letter represents significant differences between groups (p ≤ 0.05). (Q) Histogram of the frequency and birefringence values showing differences in the distribution of values in the different groups. Bars = 100 μm and 200 μm (a).



© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Aro, A.A.; Carneiro, G.D.; Teodoro, L.F.R.; Da Veiga, F.C.; Ferrucci, D.L.; Simões, G.F.; Simões, P.W.; Alvares, L.E.; De Oliveira, A.L.R.; Vicente, C.P.; et al. Injured Achilles Tendons Treated with Adipose-Derived Stem Cells Transplantation and GDF-5. Cells 2018, 7, 127. https://doi.org/10.3390/cells7090127
De Aro AA, Carneiro GD, Teodoro LFR, Da Veiga FC, Ferrucci DL, Simões GF, Simões PW, Alvares LE, De Oliveira ALR, Vicente CP, et al. Injured Achilles Tendons Treated with Adipose-Derived Stem Cells Transplantation and GDF-5. Cells. 2018; 7(9):127. https://doi.org/10.3390/cells7090127
Chicago/Turabian StyleDe Aro, Andrea Aparecida, Giane Daniela Carneiro, Luis Felipe R. Teodoro, Fernanda Cristina Da Veiga, Danilo Lopes Ferrucci, Gustavo Ferreira Simões, Priscyla Waleska Simões, Lúcia Elvira Alvares, Alexandre Leite R. De Oliveira, Cristina Pontes Vicente, and et al. 2018. "Injured Achilles Tendons Treated with Adipose-Derived Stem Cells Transplantation and GDF-5" Cells 7, no. 9: 127. https://doi.org/10.3390/cells7090127
APA StyleDe Aro, A. A., Carneiro, G. D., Teodoro, L. F. R., Da Veiga, F. C., Ferrucci, D. L., Simões, G. F., Simões, P. W., Alvares, L. E., De Oliveira, A. L. R., Vicente, C. P., Gomes, C. P., Pesquero, J. B., Esquisatto, M. A. M., De Campos Vidal, B., & Pimentel, E. R. (2018). Injured Achilles Tendons Treated with Adipose-Derived Stem Cells Transplantation and GDF-5. Cells, 7(9), 127. https://doi.org/10.3390/cells7090127