Is NO the Answer? The Nitric Oxide Pathway Can Support Bone Morphogenetic Protein 2 Mediated Signaling

 ,
,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. BMP Reporter Assay
2.3. Alkaline Phosphatase Activity (ALP) Assay
2.4. Gene Expression Analysis
2.5. NO Quantification
2.6. cGMP ELISA
2.7. SMAD-1 ELISA
2.8. Statistics
3. Results
3.1. BMP2-Mediated Signaling and Osteogenic Activity Can be Enhanced by Arginine Via NOS
3.1.1. Arginine Supports BMP2-Mediated Activity
3.1.2. Arginine Support of BMP2 Activity is Mediated by NOS
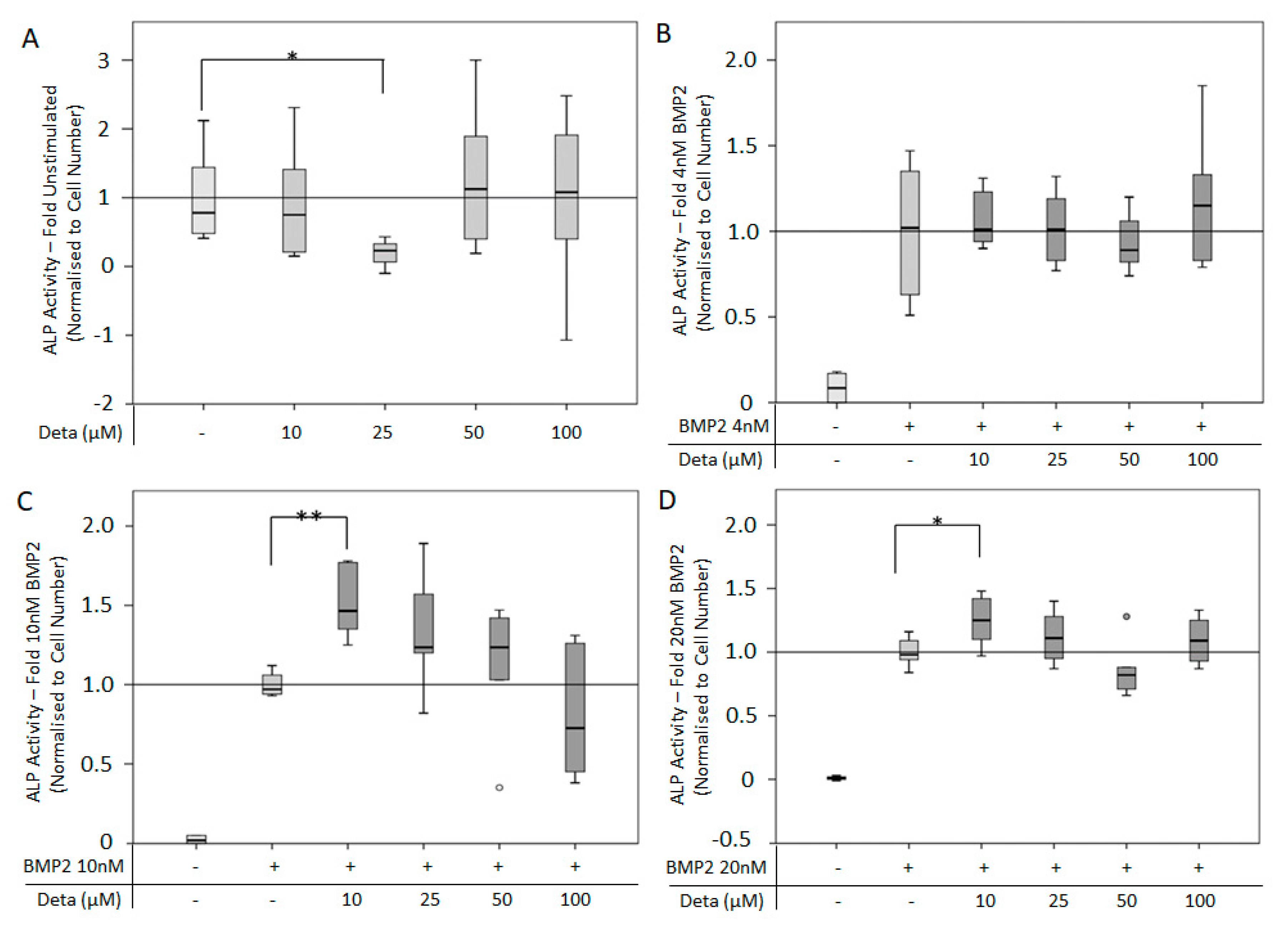
3.2. BMP2-Mediated Signaling and Osteogenesis Can be Enhanced by Direct NO Donor
3.3. NO Receptor, sGC, Assists in BMP2 Induced Signaling
3.4. SMAD Signaling after BMP and NO Stimulation
3.5. PKA Connects BMP2 and NO Pathway for BMP2 Signaling
3.6. BMP2 Can Induce NO Pathway Activity
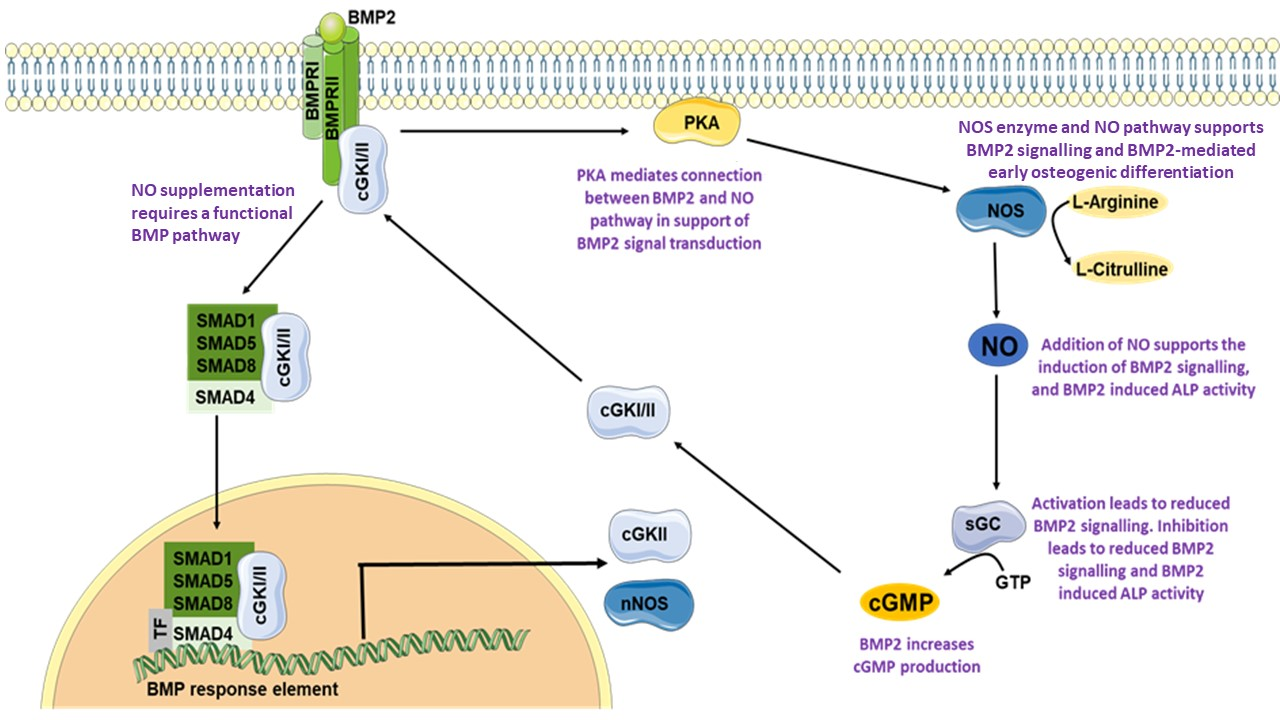
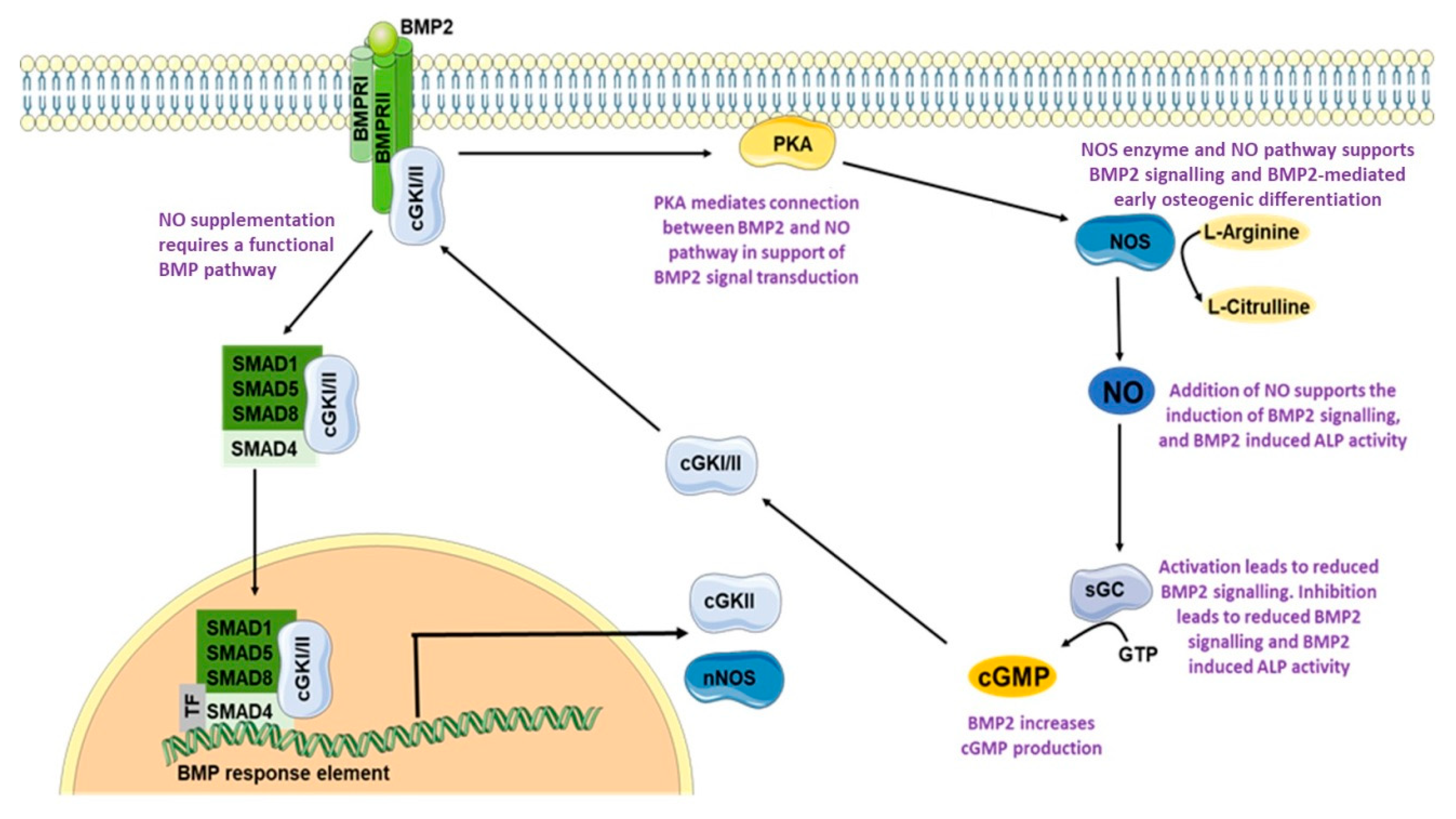
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| 18S | ribosomal unit 18S | LDN | LDN-193189 |
| ALP | Alkaline Phosphatase | Luc | Luciferase |
| AMHR | anti-Mullerian hormone receptor type | MNE | mean normalized expression |
| Arginine BMP | L-Arginine Bone Morphogenic Protein | nNOS | neuronal Nitric Oxide Synthase |
| BMPR | BMP receptor | NO OPG | Nitric Oxide Osteoprotegerin |
| BMPR2-LF/-SF | BMP receptor - long form/- short form | PAEC | pulmonary arterial endothelial cells |
| BRE | BMP response elements | PBS | Phosphate Buffered Saline |
| cGKI/II | cGMP dependent kinase I/II | PDE | phosphodiesterase’s |
| cGMP | cyclic guanosine monophosphate | Pen/Strep | Penicillin-Streptomycin |
| CO2 Col1a1 | Carbon Dioxide Type 1 collagen | PKA | Protein Kinase A |
| Co-SMAD | common mediator SMAD | pNPP | 4-nitrophenyl phosphate |
| CREB | cAMP response element-binding - | PP2 | protein phosphatase 2 |
| CT | Crossing Threshold | rhBMP2 | recombinant human BMP2 |
| DAF-2T | 4,5-Diaminofluorescein triazole | RLU | relative light units |
| Deta | Deta NONOate | R-SMAD | receptor regulated SMAD |
| DMEM | Dulbecco Modified Eagle Medium | Runx2 | Runt-related transcription factor 2 |
| ECM | extracellular matrix | sGC | soluble Guanylyl Cyclase |
| ELISA | Enzyme Linked Immunosorbent Assay | SMAD | Small Mothers Against Decapentaplegic |
| eNOS | Endothelial Nitric Oxide Synthase | TF | Transcription Factors |
| Exp. | Experiment | TGFβ | Transforming Growth Factor β |
| FCS | Fetal Calf Serum | ||
| Id1,2,3 | Inhibitor of Differentiation 1,2,3 |
References
- Wu, M.; Chen, G.; Li, Y.-P. TGF-β and BMP signaling in osteoblast, skeletal development, and bone formation, homeostasis and disease. Bone Res. 2016, 4, 16009. [Google Scholar] [CrossRef] [PubMed]
- Klatte-Schulz, F.; Giese, G.; Differ, C.; Minkwitz, S.; Ruschke, K.; Puts, R.; Knaus, P.; Wildemann, B. An investigation of BMP-7 mediated alterations to BMP signalling components in human tenocyte-like cells. Sci. Rep. 2016, 6, 29703. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Michalski, N.; Kronander, E.; Gjoni, E.; Genoud, C.; Knott, G.; Schneggenburger, R. BMP signaling specifies the development of a large and fast CNS synapse. Nat. Neurosci. 2013, 16, 856–864. [Google Scholar] [CrossRef]
- Dyer, L.A.; Pi, X.; Patterson, C. The role of BMPs in endothelial cell function and dysfunction. Trends Endocrinol. Metab. 2014, 25, 472–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, I.H.; Brazil, D.P. Bone morphogenetic proteins and their antagonists: Current and emerging clinical uses. Br. J. Pharmacol. 2014, 171, 3620–3632. [Google Scholar] [CrossRef]
- Alt, V.; Borgman, B.; Eicher, A.; Heiss, C.; Kanakaris, N.K.; Giannoudis, P.V.; Song, F. Effects of recombinant human Bone Morphogenetic Protein-2 (rhBMP-2) in grade III open tibia fractures treated with unreamed nails—A clinical and health-economic analysis. Injury 2015, 46, 2267–2272. [Google Scholar] [CrossRef]
- Lyon, T.R.; Pollak, A.N.; Valentin-Opran, A.; Jones, A.L.; Bucholz, R.W.; Bosse, M.J.; Mirza, S.K.; Webb, L.X.; Golden, J.D. Recombinant Human BMP-2 and Allograft Compared with Autogenous Bone Graft for Reconstruction of Diaphyseal Tibial Fractures with Cortical Defects. J. Bone Jt. Surg. 2006, 88, 1431–1441. [Google Scholar] [CrossRef]
- Wei, S.; Cai, X.; Huang, J.; Xu, F.; Liu, X.; Wang, Q. Recombinant Human BMP-2 for the Treatment of Open Tibial Fractures. Orthopedics 2012, 35, e847–e854. [Google Scholar] [CrossRef]
- Barcak, E.A.; Beebe, M.J. Bone Morphogenetic Protein: Is there Still a Role in Orthopedic Trauma in 2017? Orthop. Clin. North Am. 2017, 48, 301–309. [Google Scholar] [CrossRef]
- Aro, H.T.; Govender, S.; Patel, A.D.; Hernigou, P.; De Gregorio, A.P.; Popescu, G.I.; Golden, J.D.; Christensen, J.; Valentin, A. Recombinant Human Bone Morphogenetic Protein-2: A Randomized Trial in Open Tibial Fractures Treated with Reamed Nail Fixation. J. Bone Jt. Surg. 2011, 93, 801–808. [Google Scholar] [CrossRef]
- Govender, S.; Csimma, C.; Genant, H.K.; Valentin-Opran, A.; Amit, Y.; Arbel, R.; Aro, H.; Atar, D.; Bishay, M.; Börner, M.G.; et al. Recombinant human bone morphogenetic protein-2 for treatment of open tibial fractures: A prospective, controlled, randomized study of four hundred and fifty patients. J. Bone Jt. Surg. 2002, 84, 2123–2134. [Google Scholar] [CrossRef] [PubMed]
- TerMaat, M.; Boer, F.D.; Bakker, F.; Patka, P.; Haarman, H. Bone Morphogenetic ProteinsDevelopment and Clinical Efficacy in the Treatment of Fractures and Bone Defects. J. Bone Jt. Surg. 2005, 87, 1367–1378. [Google Scholar] [CrossRef]
- Zara, J.N.; Siu, R.K.; Zhang, X.; Shen, J.; Ngo, R.; Lee, M.; Li, W.; Chiang, M.; Chung, J.; Kwak, J.; et al. High Doses of Bone Morphogenetic Protein 2 Induce Structurally Abnormal Bone and Inflammation In Vivo. Tissue Eng. Part A 2011, 17, 1389–1399. [Google Scholar] [CrossRef]
- Stanton, T. Lessons Learned: What the BMP Trials Controversy Has Taught Us: Lessons Learned: What the BMP Trials Controversy Has Taught Us. Available online: https://www.aaos.org/News/The_Daily_Edition_of_AAOS_Now/2012/Thursday,_February_9/AAOS9_2_9/?ssopc=1. (accessed on 12 March 2018).
- Lyon, T.; Scheele, W.; Bhandari, M.; Koval, K.J.; Sanchez, E.G.; Christensen, J.; Valentin, A.; Huard, F. Efficacy and safety of recombinant human bone morphogenetic protein-2/calcium phosphate matrix for closed tibial diaphyseal fracture: A double-blind, randomized, controlled phase-II/III trial. J. Bone Jt. Surg. 2013, 95, 2088–2096. [Google Scholar] [CrossRef] [PubMed]
- Chan, D.S.; Garland, J.; Infante, A.; Sanders, R.W.; Sagi, H.C. Wound Complications Associated with Bone Morphogenetic Protein-2 in Orthopaedic Trauma Surgery. J. Orthop. Trauma 2014, 28, 599–604. [Google Scholar] [CrossRef] [PubMed]
- Heldin, C.-H.; Miyazono, K.; Dijke, P.T. TGF-β signalling from cell membrane to nucleus through SMAD proteins. Nature 1997, 390, 465–471. [Google Scholar] [CrossRef] [PubMed]
- Derynck, R.; Zhang, Y.E. Smad-dependent and Smad-independent pathways in TGF-β family signalling. Nature 2003, 425, 577–584. [Google Scholar] [CrossRef] [PubMed]
- Yadin, D.; Knaus, P.; Mueller, T.D.; Information, P.E.K.F.C. Structural insights into BMP receptors: Specificity, activation and inhibition. Cytokine Growth Factor Rev. 2016, 27, 13–34. [Google Scholar] [CrossRef]
- Liu, F.; Ventura, F.; Doody, J.; Massague, J. Human type II receptor for bone morphogenic proteins (BMPs): Extension of the two-kinase receptor model to the BMPs. Mol. Cell. Biol. 1995, 15, 3479–3486. [Google Scholar] [CrossRef]
- Ishikawa, T.; Nohno, T.; Saito, T.; Hosokawa, K.; Noji, S.; Wolsing, D.H.; Rosenbaum, J.S. Identification of a Human Type II Receptor for Bone Morphogenetic Protein-4 That Forms Differential Heteromeric Complexes with Bone Morphogenetic Protein Type I Receptors. J. Biol. Chem. 1995, 270, 22522–22526. [Google Scholar] [Green Version]
- Rosenzweig, B.L.; Imamura, T.; Okadome, T.; Cox, G.N.; Yamashita, H.; Dijke, P.T.; Heldin, C.H.; Miyazono, K. Cloning and characterization of a human type II receptor for bone morphogenetic proteins. Proc. Natl. Acad. Sci. USA 1995, 92, 7632–7636. [Google Scholar] [CrossRef] [PubMed]
- Bengtsson, L.; Schwappacher, R.; Roth, M.; Boergermann, J.H.; Hassel, S.; Knaus, P. PP2A regulates BMP signalling by interacting with BMP receptor complexes and by dephosphorylating both the C-terminus and the linker region of Smad1. J. Cell Sci. 2009, 122, 1248–1257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, M.C.; Nguyen, P.H.; Davis, B.N.; Ohoka, N.; Hayashi, H.; Du, K.; Lagna, G.; Hata, A. A Novel Regulatory Mechanism of the Bone Morphogenetic Protein (BMP) Signaling Pathway Involving the Carboxyl-Terminal Tail Domain of BMP Type II Receptor. Mol. Cell. Biol. 2007, 27, 5776–5789. [Google Scholar] [CrossRef] [Green Version]
- Hassel, S.; Yakymovych, M.; Hellman, U.; Rönnstrand, L.; Knaus, P.; Souchelnytskyi, S. Interaction and functional cooperation between the serine/threonine kinase bone morphogenetic protein type II receptor with the tyrosine kinase stem cell factor receptor. J. Cell. Physiol. 2006, 206, 457–467. [Google Scholar] [CrossRef]
- Schwappacher, R.; Weiske, J.; Heining, E.; Ezerski, V.; Marom, B.; Henis, Y.I.; Huber, O.; Knaus, P. Novel crosstalk to BMP signalling: cGMP-dependent kinase I modulates BMP receptor and Smad activity. EMBO J. 2009, 28, 1537–1550. [Google Scholar] [CrossRef] [Green Version]
- Gangopahyay, A.; Oran, M.; Bauer, E.M.; Wertz, J.W.; Comhair, S.A.; Erzurum, S.C.; Bauer, P.M. Bone Morphogenetic Protein Receptor II Is a Novel Mediator of Endothelial Nitric-oxide Synthase Activation*. J. Biol. Chem. 2011, 286, 33134–33140. [Google Scholar] [CrossRef]
- Hassel, S.; Eichner, A.; Yakymovych, M.; Hellman, U.; Knaus, P.; Souchelnytskyi, S. Proteins associated with type II bone morphogenetic protein receptor (BMPR-II) and identified by two-dimensional gel electrophoresis and mass spectrometry. Proteomics 2004, 4, 1346–1358. [Google Scholar] [CrossRef]
- Rangaswami, H.; Schwappacher, R.; Marathe, N.; Zhuang, S.; Casteel, D.E.; Haas, B.; Chen, Y.; Pfeifer, A.; Kato, H.; Shattil, S.; et al. Cyclic GMP and protein kinase G control a Src-containing mechanosome in osteoblasts. Sci. Signal. 2010, 3, ra91. [Google Scholar] [CrossRef]
- Broulik, P.D.; Rosenkrancová, J.; Ruzicka, P.; Sedlacek, R. Effects of Triiodothyronine and Estrogen Administration on Bone Mass, Mineral Content and Bone Strength in Male Rats. Horm. Metab. Res. 2003, 35, 527–531. [Google Scholar] [CrossRef]
- Wimalawansa, S.; De Marco, G.; Gangula, P.; Yallampalli, C.; Wimalawansa, P.S. Nitric oxide donor alleviates ovariectomy-induced bone loss. Bone 1996, 18, 301–304. [Google Scholar] [CrossRef]
- Wimalawansa, S.J. Restoration of Ovariectomy-Induced Osteopenia by Nitroglycerin. Calcif. Tissue Int. 2000, 66, 56–60. [Google Scholar] [CrossRef] [PubMed]
- Wimalawansa, S.; Chapa, M.; Yallampalli, C.; Zhang, R.; Simmons, D.; Wimalawansa, P.S. Prevention of corticosteroid-induced bone loss with nitric oxide donor nitroglycerin in male rats. Bone 1997, 21, 275–280. [Google Scholar] [CrossRef]
- Wimalawansa, S.J. Nitroglycerin Therapy Is as Efficacious as Standard Estrogen Replacement Therapy (Premarin) in Prevention of Oophorectomy-Induced Bone Loss: A Human Pilot Clinical Study. J. Bone Miner. Res. 2000, 15, 2240–2244. [Google Scholar] [CrossRef] [PubMed]
- Jamal, S.A.; Hamilton, C.J.; Eastell, R.; Cummings, S.R. Effect of nitroglycerin ointment on bone density and strength in postmenopausal women: A randomized trial. JAMA 2011, 305, 800–807. [Google Scholar] [CrossRef]
- Jamal, S.A.; Browner, W.S.; Bauer, D.C.; Cummings, S.R. Intermittent Use of Nitrates Increases Bone Mineral Density: The Study of Osteoporotic Fractures. J. Bone Miner. Res. 1998, 13, 1755–1759. [Google Scholar] [CrossRef]
- Boergermann, J.; Kopf, J.; Yu, P.; Knaus, P. Dorsomorphin and LDN-193189 inhibit BMP-mediated Smad, p38 and Akt signalling in C2C12 cells. Int. J. Biochem. Cell Biol. 2010, 42, 1802–1807. [Google Scholar] [CrossRef]
- Sondag, G.R.; Salihoglu, S.; Lababidi, S.L.; Crowder, D.C.; Moussa, F.M.; Abdelmagid, S.M.; Safadi, F.F. Osteoactivin Induces Transdifferentiation of C2C12 Myoblasts Into Osteoblasts. J. Cell. Physiol. 2014, 229, 955–966. [Google Scholar] [CrossRef]
- Tanaka, K.-I.; Matsumoto, E.; Higashimaki, Y.; Katagiri, T.; Sugimoto, T.; Seino, S.; Kaji, H. Role of Osteoglycin in the Linkage between Muscle and Bone. J. Biol. Chem. 2012, 287, 11616–11628. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.H.; Javed, A.; Kim, H.J.; Shin, H.I.; Gutierrez, S.; Choi, J.Y.; Rosen, V.; Stein, J.L.; Van Wijnen, A.J.; Stein, G.S.; et al. Transient upregulation of CBFA1 in response to bone morphogenetic protein-2 and transforming growth factor beta1 in C2C12 myogenic cells coincides with suppression of the myogenic phenotype but is not sufficient for osteoblast differentiation. J. Cell. Biochem. 1999, 73, 114–125. [Google Scholar] [CrossRef]
- Herrera, B.; Inman, G.J. A rapid and sensitive bioassay for the simultaneous measurement of multiple bone morphogenetic proteins. Identification and quantification of BMP4, BMP6 and BMP9 in bovine and human serum. BMC Cell Biol. 2009, 10, 20. [Google Scholar] [CrossRef]
- Hinchee-Rodriguez, K.; Garg, N.; Venkatakrishnan, P.; Roman, M.G.; Adamo, M.L.; Masters, B.S.; Roman, L.J. Neuronal nitric oxide synthase is phosphorylated in response to insulin stimulation in skeletal muscle. Biochem. Biophys. Res. Commun. 2013, 435, 501–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.; Jiao, H.; Zhao, J.; Wang, X.; Lin, H. L-Arginine Enhances Protein Synthesis by Phosphorylating mTOR (Thr 2446) in a Nitric Oxide-Dependent Manner in C2C12 Cells. Oxidative Med. Cell. Longev. 2018, 2018, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strobel, C.; Bormann, N.; Kadow-Romacker, A.; Schmidmaier, G.; Wildemann, B. Sequential release kinetics of two (gentamicin and BMP-2) or three (gentamicin, IGF-I and BMP-2) substances from a one-component polymeric coating on implants. J. Control. Release 2011, 156, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Garrison, K.R.; Shemilt, I.; Donell, S.; Ryder, J.J.; Mugford, M.; Harvey, I.; Song, F.; Alt, V. Bone morphogenetic protein (BMP) for fracture healing in adults. Cochrane Database Syst. Rev. 2010, 2010, CD006950. [Google Scholar] [CrossRef] [PubMed]
- Ferrero, R.; Rodríguez-Pascual, F.; Miras-Portugal, M.T.; Torres, M. Comparative effects of several nitric oxide donors on intracellular cyclic GMP levels in bovine chromaffin cells: Correlation with nitric oxide production. Br. J. Pharmacol. 1999, 127, 779–787. [Google Scholar] [CrossRef] [PubMed]
- Keefer, L.K.; Nims, R.W.; Davies, K.M.; Wink, D.A. “NONOates” (1-substituted diazen-1-ium-1,2-diolates) as nitric oxide donors: Convenient nitric oxide dosage forms. In Methods in Enzymology; Packer, L., Cadenas, E., Eds.; Academic Press: Cambridge, MA, USA, 1996; pp. 281–293. [Google Scholar]
- Wang, P.G.; Xian, M.; Tang, X.; Wu, X.; Wen, Z.; Cai, T.; Janczuk, A.J. Nitric Oxide Donors: Chemical Activities and Biological Applications. Chem. Rev. 2002, 102, 1091–1134. [Google Scholar] [CrossRef]
- Soltow, Q.A.; Lira, V.A.; Betters, J.L.; Long, J.H.D.; Sellman, J.E.; Zeanah, E.H.; Criswell, D.S. Nitric oxide regulates stretch-induced proliferation in C2C12 myoblasts. J. Muscle Res. Cell Motil. 2010, 31, 215–225. [Google Scholar] [CrossRef]
- Babich, H.; Zuckerbraun, H.L.; Hirsch, S.T.; Blau, L. In vitro Cytotoxicity of the Nitric Oxide Donor, S -Nitroso- N -acetyl-peraciUamine, towards Cells from Human Oral Tissue. Pharmacol. Toxicol. 1999, 84, 218–225. [Google Scholar] [CrossRef]
- Pervin, S.; Singh, R.; Gau, C.L.; Edamatsu, H.; Tamanoi, F.; Chaudhuri, G. Potentiation of nitric oxide-induced apoptosis of MDA-MB-468 cells by farnesyltransferase inhibitor: Implications in breast cancer. Cancer Res. 2001, 61, 4701–4706. [Google Scholar]
- Dubey, M.; Nagarkoti, S.; Awasthi, D.; Singh, A.K.; Chandra, T.; Kumaravelu, J.; Barthwal, M.K.; Dikshit, M. Nitric oxide-mediated apoptosis of neutrophils through caspase-8 and caspase-3-dependent mechanism. Cell Death Dis. 2016, 7, e2348. [Google Scholar] [CrossRef]
- Makeeva, N.; Roomans, G.M.; Welsh, N. Role of TAB1 in nitric oxide-induced p38 activation in insulin-producing cells. Int. J. Biol. Sci. 2006, 3, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.-H.; Jang, M.-H.; Kim, E.-K.; Han, S.-W.; Cho, S.-Y.; Kim, C.-J. Nitric oxide induces apoptosis in mouse C2C12 myoblast cells. J. Pharmacol. Sci. 2005, 97, 369–376. [Google Scholar] [CrossRef] [PubMed]
- Makris, A.C.; Sotzios, Y.; Zhou, Z.; Makropoulou, M.; Papapetropoulos, N.; Zacharatos, P.; Pyriochou, A.; Roussos, C.; Papapetropoulos, A.; Vassilakopoulos, T. Nitric Oxide Stimulates Interleukin-6 Production in Skeletal Myotubes. J. Interf. Cytokine Res. 2010, 30, 321–327. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; De Castro, L.F.; Shin, M.H.; Dubois, W.; Yang, H.H.; Jiang, S.; Mishra, P.J.; Ren, L.; Gou, H.; Lal, A.; et al. p53 Loss Increases the Osteogenic Differentiation of Bone Marrow Stromal Cells. Stem Cells 2015, 33, 1304–1319. [Google Scholar] [CrossRef]
- Pradhan, B.B.; Bae, H.W.; Dawson, E.G.; Patel, V.V.; Delamarter, R.B. Graft Resorption with the Use of Bone Morphogenetic Protein: Lessons from Anterior Lumbar Interbody Fusion Using Femoral Ring Allografts and Recombinant Human Bone Morphogenetic Protein-2. Spine 2006, 31, E277–E284. [Google Scholar] [CrossRef]
- Lee, K.-B.; Johnson, J.S.; Song, K.-J.; Taghavi, C.E.; Wang, J.C. Use of autogenous bone graft compared with RhBMP in high-risk patients: A comparison of fusion rates and time to fusion. Clin. Spine Surg. 2013, 26, 233–238. [Google Scholar] [CrossRef]
- Drescher, W.; Beckmann, R.; Kasch, R.; Pufe, M.; Knobe, M.; Kweider, N.; Hassenpflug, J.; Tingart, M.; Pufe, T.; Kadyrov, M. Nitrate patch prevents steroid-related bone necrosis. J. Orthop. Res. 2011, 29, 1517–1520. [Google Scholar] [CrossRef]
- Nichols, S.P.; Storm, W.L.; Koh, A.; Schoenfisch, M.H. Local delivery of nitric oxide: Targeted delivery of therapeutics to bone and connective tissues. Adv. Drug Deliv. Rev. 2012, 64, 1177–1188. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Lu, X.; Xiang, F.-L.; Lu, M.; Feng, Q. Nitric Oxide Synthase-3 Promotes Embryonic Development of Atrioventricular Valves. PLoS ONE 2013, 8, e77611. [Google Scholar] [CrossRef]
- Kanazawa, I.; Yamaguchi, T.; Yano, S.; Yamauchi, M.; Sugimoto, T. Metformin enhances the differentiation and mineralization of osteoblastic MC3T3-E1 cells via AMP kinase activation as well as eNOS and BMP-2 expression. Biochem. Biophys. Res. Commun. 2008, 375, 414–419. [Google Scholar] [CrossRef]
- Kanazawa, I.; Yamaguchi, T.; Yano, S.; Yamauchi, M.; Sugimoto, T. Activation of AMP kinase and inhibition of Rho kinase induce the mineralization of osteoblastic MC3T3-E1 cells through endothelial NOS and BMP-2 expression. Am. J. Physiol. Metab. 2009, 296, E139–E146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siddappa, R.; Martens, A.; Doorn, J.; Leusink, A.; Olivo, C.; Licht, R.; Van Rijn, L.; Gaspar, C.; Fodde, R.; Janssen, F.; et al. cAMP/PKA pathway activation in human mesenchymal stem cells in vitro results in robust bone formation in vivo. Proc. Natl. Acad. Sci. USA 2008, 105, 7281–7286. [Google Scholar] [CrossRef] [PubMed]
- Lo, K.W.-H.; Kan, H.M.; Ashe, K.M.; Laurencin, C.T. The small molecule PKA-specific cyclic AMP analogue as an inducer of osteoblast-like cells differentiation and mineralization. J. Tissue Eng. Regen. Med. 2011, 6, 40–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Li, L.; Dong, Q.; Wang, Y.; Feng, Q.; Ou, X.; Zhou, P.; He, T.; Luo, J. Activation of PKA/CREB Signaling is Involved in BMP9-Induced Osteogenic Differentiation of Mesenchymal Stem Cells. Cell. Physiol. Biochem. 2015, 37, 548–562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, R.L.; Lindsey, D.P.; Dhulipala, L.; Harris, A.H.S.; Goodman, S.B.; Maloney, W.J. Effects of intermittent hydrostatic pressure and BMP-2 on osteoarthritic human chondrocyte metabolism in vitro. J. Orthop. Res. 2011, 29, 361–368. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Method | Pre-Stimulation | Stimulation | |||
|---|---|---|---|---|---|
| Media | Additional Treatment | Media & FCS | Added Factors | Time (h) | |
| Reporter activity | DMEM without Arginine, 0.1% FCS | Inhibitors: LNAME, LY83583, H89, LDN final hour | DMEM without Arginine, 0.1% FCS | Arginine, Deta, SNAP and BMP2 | 24 6 |
| ALP activity | DMEM/HAMs, 1% FCS | Inhibitors: LNAME, LY83583 final hour | DMEM/HAMs, 1% FCS | Arginine, Deta, and BMP2 | 72 |
| NO production | DMEM High Glucose, 1% FCS | DAF-2, 1 h | DMEM High Glucose, 1% FCS | Deta, and BMP2 | 2 |
| cGMP ELISA | DMEM without Arginine, 1% FCS | IBMX final 30 min | DMEM without Arginine, 1% FCS | Deta, and BMP2 | 1 |
| SMAD-1 ELISA | DMEM without Arginine, 0.1% FCS | Inhibitors: LDN, final hour | DMEM without Arginine, 0.1% FCS | Arginine and BMP2 | 0.5 |
| Gene Expression Id1,2,3 | DMEM without Arginine, 0.1% FCS | - | DMEM without Arginine, 0.1% FCS | Arginine, and BMP2 | 6 |
| Differentiation (Gene Expression OPG, Runx, Col1a1) | DMEM without Arginine, 1% FCS | - | DMEM without Arginine, 1% FCS | Arginine and BMP2 | 144 |
| Gene | Accession Number | Forward (5′→3′) | Reverse (5′→3′) |
|---|---|---|---|
| HPRT | NM_000194 | TTGCTGACCTGCTGGATTAC | AACTTTTATGTCCCCCGTTG |
| Id1 | NM_010495.3 | ACGACATGAACGGCTGCTACT | GCTCACTTTGCGGTTCTGG |
| Id2 | NM_010496.3 | TATCAGCCATTTCACCAGGAG | TGTGAAAAGGCAAAGTCTGCT |
| Id3 | NM_008321 | AAGGACAAGAGGAGCTTTTGC | GCGTTGAGTTCAGGGTAAGTG |
| OPG | NM_002546.3 | TGCAGTACGTCAAGCAGGAG | CCCATCTGGACATCTTTTGC |
| Col1a1 | NM_000088.3 | TGACCTCAAGATGTGCCACT | ACCAGACATGCCTCTTGTCC |
| Runx2 | NR_103532 | GCCCCCAAACAGTATCTTGA | GCCTGAAGTGAGGTTTTAGGC |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Differ, C.; Klatte-Schulz, F.; Bormann, N.; Minkwitz, S.; Knaus, P.; Wildemann, B. Is NO the Answer? The Nitric Oxide Pathway Can Support Bone Morphogenetic Protein 2 Mediated Signaling. Cells 2019, 8, 1273. https://doi.org/10.3390/cells8101273
Differ C, Klatte-Schulz F, Bormann N, Minkwitz S, Knaus P, Wildemann B. Is NO the Answer? The Nitric Oxide Pathway Can Support Bone Morphogenetic Protein 2 Mediated Signaling. Cells. 2019; 8(10):1273. https://doi.org/10.3390/cells8101273
Chicago/Turabian StyleDiffer, Christopher, Franka Klatte-Schulz, Nicole Bormann, Susann Minkwitz, Petra Knaus, and Britt Wildemann. 2019. "Is NO the Answer? The Nitric Oxide Pathway Can Support Bone Morphogenetic Protein 2 Mediated Signaling" Cells 8, no. 10: 1273. https://doi.org/10.3390/cells8101273
APA StyleDiffer, C., Klatte-Schulz, F., Bormann, N., Minkwitz, S., Knaus, P., & Wildemann, B. (2019). Is NO the Answer? The Nitric Oxide Pathway Can Support Bone Morphogenetic Protein 2 Mediated Signaling. Cells, 8(10), 1273. https://doi.org/10.3390/cells8101273