1. Introduction
Hypertrophic cardiomyopathy (HCM) is a disorder with a prevalence of 1/500 and an annual mortality of approximately 1% [
1]. It is a monogenic inherited disease associated with cardiac dysfunction and life threatening arrhythmias [
2,
3]. HCM exhibits a wide phenotypic variability ranging from asymptomatic to severe symptoms and is an important cause of sudden cardiac death in young adults and athletes. At the cellular level, HCM is characterized by patches of cardiomyocyte hypertrophy, cardiomyocyte disarray, interstitial fibrosis, and small vessel disease.
When subjected to hemodynamic or metabolic stress, the heart returns to fetal metabolism and the fetal gene program where the cells prefer usage of glucose over fatty acid as source of energy [
4]. It has been proposed that dysfunctional regulation of the glucose metabolism and cardiac energy metabolism is a prominent feature of the maladapted failing heart and HCM hearts [
5,
6]. A rise of glucose level results in an increased influx trough of the hexosamine biosynthetic pathway (HBP) [
7], resulting in elevated levels of uridine diphosphate
N-acetyl-
d-glucosamine (UDP-GlcNAc).
A significant consequence of higher levels of UDP-GlcNAc is the increased synthesis of the extracellular matrix (ECM) glycosaminoglycan hyaluronan (HA), which has not previously been explored in HCM. HA is a polydisperse unbranched polymer that greatly varies in molecular mass ranging from 5 to 10,000 kDa, which makes it challenging to analyze. It is present in the ECM of all vertebrates, and is highly expressed during development, wound healing, and regeneration. HA is synthesized at the cellular membrane by the linkage of the UDP-sugar precursors
d-glucuronic acid (GlcUA) and
N-acetyl-
d-glucosamine (GlcNAc), a reaction catalyzed by the membrane bound enzyme hyaluronan synthases (HAS). The HA synthesis is strongly dependent on the cytoplasmic concentrations of UDP-GlcNAc and UDP-GlcUA, and it has been shown that increased levels of these UDP-sugar precursors enhance HA synthesis [
8,
9].
Several reports have shown that diverse sizes of HA exert a wide spectrum of functions. In health and tissue homeostasis, HA is present as high molecular mass (MM) HA and has structural and hydrating features as well as an anti-inflammatory effect [
10]. On the contrary, low MM HA has been shown to have a pro-inflammatory effect [
11]. In addition, many receptors and extracellular proteins have been shown to bind HA, creating a molecular network with a wide range of structural and signaling properties. Degradation of HA is mainly carried out by hyaluronidases (HYAL, CEMIP) or by reactive oxygen species (ROS). Recent work has revealed HA as a driving factor in the development of fibrosis by stimulating both fibroblast proliferation, differentiation, and motility [
12]. Furthermore, it has been shown that high MM HA depolarizes the membrane in cell cultures in a concentration dependent manner, which could be reversed by cell surface digestion of HA by hyaluronidase [
13]. If this is also true for cardiomyocytes, it could potentially change the cardiac action potential leading to rhythm disturbances and arrhythmias [
14].
We have earlier shown elevated gene expression of
HAS 1,
2 and the HA receptor
CD44 as well as increased cardiac levels of HA correlating with pro-hypertrophic gene expression, using a rat model for cardiac hypertrophy [
15,
16]. We have also identified a crosstalk between cultured cardiomyocytes and fibroblasts resulting in increased HA synthesis in the fibroblasts [
17]. In addition, HA staining was stronger in human cardiac tissues from HCM patients compared to autopsy material from previously healthy individuals [
18], and in the rat heart HA occurs around myofibrils [
19].
In this study we further investigate HA in HCM. We observed an altered metabolism of HA in HCM and changes in molecular mass distribution of HA corresponding with cardiomyocyte size.
2. Materials and Methods
2.1. Human Tissue Samples
Tissue aliquots from human septal myocardium were obtained during surgery with basal septal myectomy from five patients with hypertrophic obstructive cardiomyopathy. Two of the patients were diagnosed with coronary disease. None were diagnosed with diabetes or hypertension. Characteristics are presented in
Table 1.
Control (non-failing) human left ventricular, right ventricular, and septal tissue was obtained from five sex- and age-matched subjects whose hearts were rejected as cardiac donors for surgical reasons. The cause of death of donors was cerebrovascular accident or carbon monoxide poisoning, and none had a history of heart disease or known previous medication. Myocardium from these subjects was kept on ice for 1 to 4 h before tissue sampling. All cardiac tissues were snap-frozen in liquid nitrogen and stored at −80 °C until use.
2.2. Rat Model for Hypertrophy
Cardiac tissues were analyzed from a rat model of cardiac hypertrophy from a study published several years ago (2008) [
15]. Due to the newly developed GEMMA (gas-phase electrophoretic molecular mobility analyzer) analysis of HA mass distribution, it was now possible to perform HA mass analysis on the small pieces of cardiac tissue saved from the previous study [
20]. The surgical procedure of the rats has been described elsewhere [
15]. Briefly, rats were anesthetized with 0.2 mL pentobarbital intra-abdominal and after abdominal incision, a titanium clip of 1.15 mm inner diameter was administered on the aorta proximal to the renal arteries on male Wistar rats (n = 3). Age-matched control rats (n = 3) were sham operated, i.e., subjected to the same procedure but without the administration of a clip on the aorta. After being anesthetized with 0.4 mL pentobarbital, the rats were sacrificed at 1 and 42 days after operation and their hearts were harvested.
2.3. Compliance with Ethical Standards
All procedures performed in studies involving human participants were in accordance with the ethical standards of the Regional Health Authorities of South-Eastern Norway and with the 1964 Helsinki declaration and its later amendments. Informed consent was obtained from all individual participants included in the study.
The animal study was performed in Paris at Inserm U689 and all procedures performed in studies involving animals were in accordance with animal welfare regulations, and the study protocol was approved by the Ethical Committee of Inserm. All investigations conformed to guidelines set by the French Ministry of Agriculture and the Guide for the Care and Use of Laboratory Animals published by the US National Institutes of Health.
2.4. 1H High-Resolution Magic Angle Spinning Nuclear Magnetic Resonance (HR MAS NMR) Spectroscopy
Tissue samples were thawed at room temperature and kept on ice during preparation. Each tissue sample (20–30 mg wet weight) was inserted into disposable 30-µL Teflon NMR inserts followed by the addition of deuterium oxide to the insert to complete the required volume and homogenize insert contents. Inserts were then packed into a 4 mm zirconia MAS rotor (40 μL capacity). All the NMR experiments on tissue samples were carried out on a 500 MHz MAS-NMR spectrometer (Bruker, Billerica, MA, USA) at 300 K.
2.5. NMR Data Processing and Analysis
Spectra were imported into MATLAB (R2015a) (MathWorks Inc., Natick, MA, USA) integrated using in-house developed scripts and normalized by the sum of all intensities. All metabolites in human cardiac tissues were identified using Chenomx NMR Suite 7.7 software (Chenomx Inc., Edmonton, AB, Canada) with full resolution NMR data and a standard two-dimensional (2D) NMR experiment on a selected sample. For identifying metabolites contributing to the discrimination between groups, the normalized 1H-NMR data were uploaded to SIMCA (version 14, Umetrics, Umeå, Sweden) for orthogonal partial least squares-discriminant analysis (OPLS-DA). The spectral variables were scaled to unit variance, and 7-fold internal cross-validation was performed to evaluate the quality of the resulting statistical models by considering the diagnostic measures R2 and Q2. Potential metabolites were selected based on the variable importance in projection (VIP) score > 1.0.
2.6. Immunohistochemistry
Before paraffin embedding, human cardiac tissues were fixed in 4% buffered formaldehyde for 48 h in room temperature. Paraffin sections (4 µm) were then mounted on SuperFrost Plus Slides (Thermo Fisher Scientific Inc., Waltham, MA, USA) and dried overnight at 37 °C. Prior to staining, sections were deparaffinized in xylene, rehydrated in series of graded ethanol, and washed in PBS. To visualize HA only, deparaffinized histological sections were blocked for 30 min using 2% bovine serum in PBS. Samples were incubated with a biotinylated HA binding protein (HABP) probe overnight at 4 °C [
21]. HABP was prepared at the Institute of Medical and Physiological Chemistry, University of Uppsala, at a concentration of 100 µg/mL and was diluted 1:40 [
22]. The samples were rinsed three times in PBS and then incubated with streptavidin 488 (1:500, Thermo Fisher Scientific Inc., MA, USA) for 1 h at room temperature.
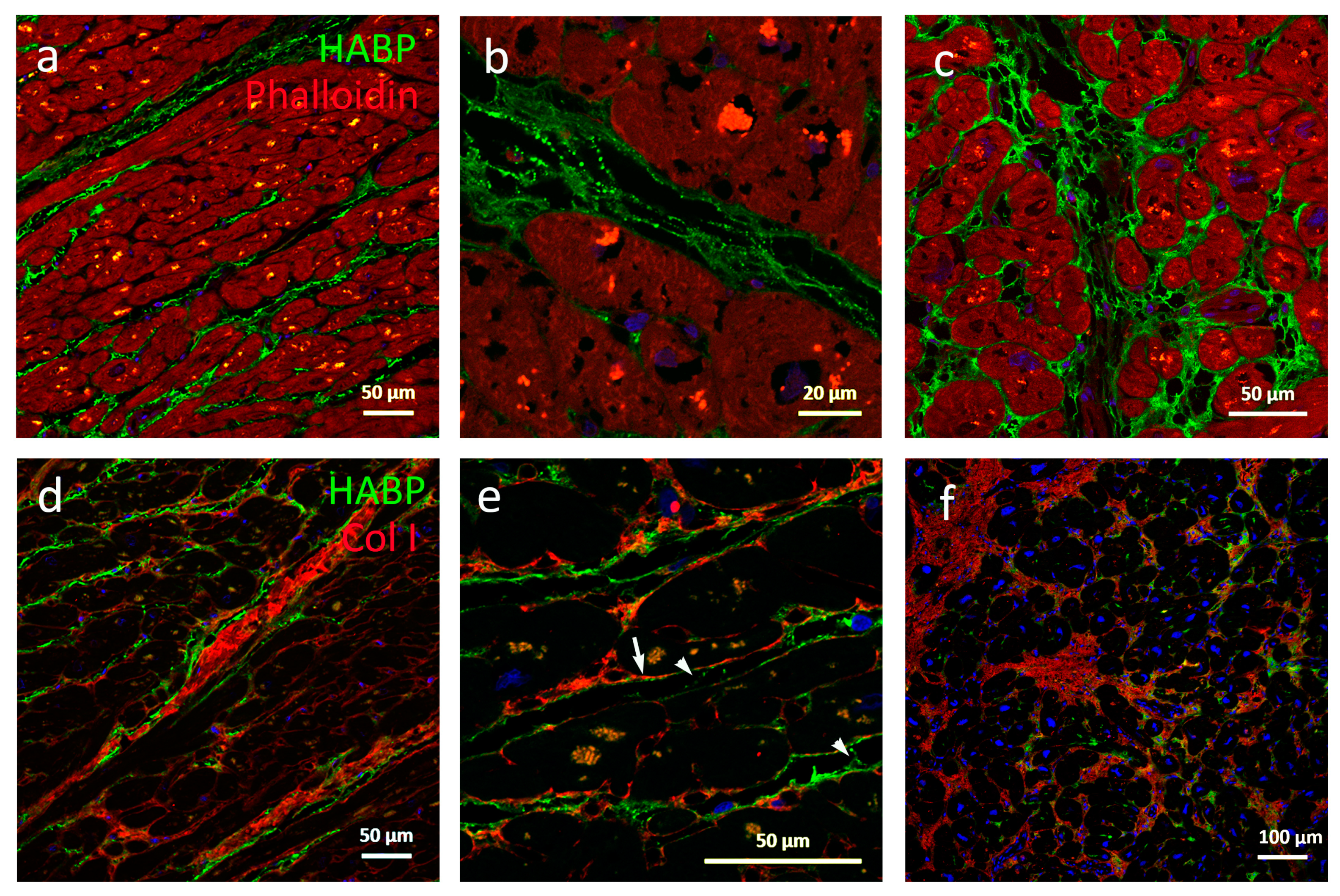
To visualize HA and collagen (I, III, and VI, respectively) in the same section, antigen retrieval was performed by boiling the sections in a citrate buffer in a microwave oven for 9 min followed by cooling for 30 min before washing in water for 5 min. The sections were then blocked for 30 min using 2% bovine serum in PBS. Finally, samples were incubated overnight at 4 °C with the primary reagents HABP and rabbit anti collagen I (ab292 1:200, Abcam, Cambridge, UK), rabbit anti collagen III (ab7778 1:400, Abcam, UK), or rabbit anti collagen VI (ab6588 1:400, Abcam, UK), respectively. After washing in PBS, the samples were incubated with streptavidin 488 (1:500, Thermo Fisher Scientific Inc., MA, USA) and donkey anti rabbit 594 (1:500, Thermo Fisher Scientific Inc., MA, USA). Nuclei were stained with Hoechst 33,343 (1:5000, Thermo Fisher Scientific Inc., MA, USA), and slides were mounted in ProLong Gold Antifade (Thermo Fisher Scientific Inc., MA, USA). Phalloidin-iFluor 594 1:1000 (ab176757, Abcam, UK) was used as a cell marker. All stainings were visualized using the Zeiss LSM 710 laser-scanning confocal microscope (Zeiss, Oberkochen, Germany).
2.7. Cardiomyocyte Area Analysis
Cardiomyocytes were visualized by iFluor 594-labeled phalloidin (1:1000, Abcam, UK) staining actin and ECM was visualized by staining collagen III (ab 7778 1:400, Abcam, UK). Each slide was objected to 10 images. Morphometric analysis was performed using the software Fiji (
http://fiji.sc). An average area value from each heart was calculated by the use of the measurements of 30–50 cells containing a central nucleus. Statistical analysis was performed using a two-tailed Student t test assuming unequal variances. Each pixel was calibrated to 0.83 μm according to Zeiss confocal data. Fluorescence images were collected at 20× objective using Zeiss LSM 710 laser-scanning confocal microscope (Zeiss, Oberkochen, Germany).
2.8. HA Molecular Mass Distribution
Cardiac tissue samples, wet weight 27–112 mg, were dried (n = 5 in each group) and homogenized. Proteins and nucleic acids were digested with proteinase K (Sigma-Aldrich, St. Louis, MO, USA), benzonase nuclease (Sigma-Aldrich, MO, USA), and chondroitinase ABC (Sigma-Aldrich, MO, USA) on three consecutive days. At the end of each day, chloroform was added to each sample and the extracted aqueous phase was dialyzed against 0.1 M NaCl using Amicon Ultra 3K concentration units (Millipore, Burlington, MA, USA) followed by overnight precipitation in 99% ethanol (EtOH). Samples were then loaded on anion exchange mini spin columns (Thermo Fisher Scientific, MA, USA) and centrifuged to wash out sulphated glycosaminoglycans and remaining non-HA contaminants, based on NaCl-binding. Finally, to remove salt the sample was dialyzed against 20 mM ammonium acetate (pH 8.0) in Amicon Ultra 3K concentration units. HA molecular mass analyses were performed using a nano-electrospray gas-phase electrophoretic molecular mobility analyzer (GEMMA) (TSI Corp., Shoreview, MN, USA).
Each sample of purified HA (n = 5 in each group) was scanned three times in the GEMMA and the final size distribution spectrum was a sum of the three scans. The raw counts from the GEMMA spectrum were calibrated according to the previously described method [
23]. The molecule diameter analyzed in the GEMMA was converted to molecular mass by analyzing HA standards ranging from 30 to 2500 kDa (Hyalose LLC, OC, USA). The relation between area under the curve (AUC) in the GEMMA spectrum to the HA concentration enables an estimation of the relative concentration of different MM of HA. Counts on the Y-axis correspond to the number of detected molecules and the X-axis to the MM of HA. Number of counts were normalized to the dry weight of the sample. Due to the physical properties of HA and the shape dependence of the GEMMA method, the analysis achieves a good separation of low MM HA up to ca. 100 kDa whereas the resolution for higher MM is poorer. The relative amount of HA with an MM less than about 100 kDa cannot be compared with HA of an MM greater than 100 kDa [
23].
In this study, we have defined low MM HA as a mass up to 50 kDa. HA extracted from cardiac tissue was degraded with hyaluronidase from Streptomyces hyalurolyticus (Sigma-Aldrich, MO, USA) and reanalyzed to test for extraction specificity.
2.9. RNA Extraction and qRT-PCR
To obtain RNA, the cardiac tissues were homogenized in Qiazol lysis agent and with beads using Precellys lysing kit (Bertin Instruments, Montigny-le-Bretonneux, France) and purified using the RNeasy plus Universal Mini Kit (QIAGEN, Waltham, MA, USA). Reverse transcription was performed with 1 µg of total RNA using the High Capacity RNA to DNA kit (Thermo Fisher Scientific, MA, USA). The extracted RNA and cDNA concentration, respectively, were quantified using a NanoDrop Spectrometer ND-1000 (NanoDrop, Thermo Fisher Scientific Inc., Waltham, MA, USA).
The real-time quantitative PCR was performed on a 7900 HT Fast Real-Time PCR system (Thermo Fisher Scientific, MA, USA) using 1 µg cDNA, TaqMan® Gene Expression Assays, and 1 µL Gene Assay Mix for the genes
HAS1-3,
HYAL 1 and
2,
CEMIP,
CD44,
VCAN, and
TSG6 (Thermo Fisher Scientific, MA, USA).
GAPDH (Thermo Fisher Scientific, MA, USA) was used as an endogenous reference gene. Forty cycles of amplification were performed. The gene of interest was normalized to the reference gene using the ΔCt method [
24].
2.10. Statistical Analysis
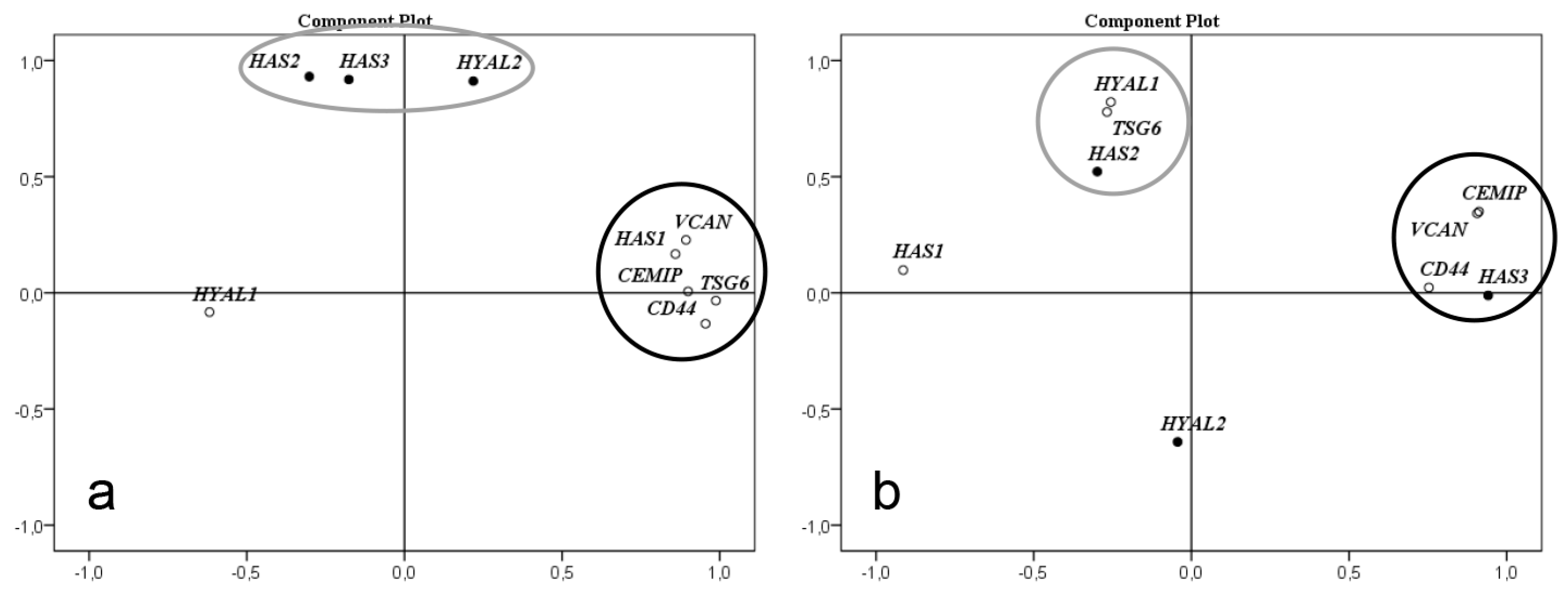
For metabolomics statistics, non-parametric Mann–Whitney U test with Benjamini–Hochberg correction using in-house software written and compiled in MATLAB (MathWorks Inc., Natick, MA, USA) was used. For analysis of gene expression and relative amount of water, low and high MM HA non-parametric independent Mann–Whitney U test was performed using the SPSS statistic software (version 25, IBM, Armonk, NY, USA). P values of less than 0.05 were considered to be significant. Factor analysis was performed with the principal components method to analyze the correlation matrix and two factors were extracted.
Generation of box plots for cardiomyocyte area was performed using SPSS statistic software.
4. Discussion
During development of HCM, the heart is subjected to hemodynamic or metabolic stress and returns to the fetal gene program and to fetal metabolism where cells prefer usage of glucose over fatty acid as a source of energy [
4,
25]. Our study material is based on basal septal myectomies from HCM patients subjected to surgical intervention due to a symptomatic and advanced disease where fibrosis and disarranged morphology is already established.
NMR analysis of metabolites in cardiac tissue from the HCM patients compared to healthy heart tissues from non-failing controls showed that fatty acids were decreased, and lactate was increased in the heart of HCM patients, indicating altered and less efficient glucose metabolism. UDP-sugar precursors, e.g., UDP-GlcNAc and UDP-GlcUA, the two substrates for HA synthesis, were also increased in the heart of HCM patients, which could be explained by the observed increase of metabolites in the HBP resulting in subsequent increase of UDP-GlcNAc. In addition, myo-Inositol and UDP-glucose, which are precursors for UDP-GlcUA, were also increased.
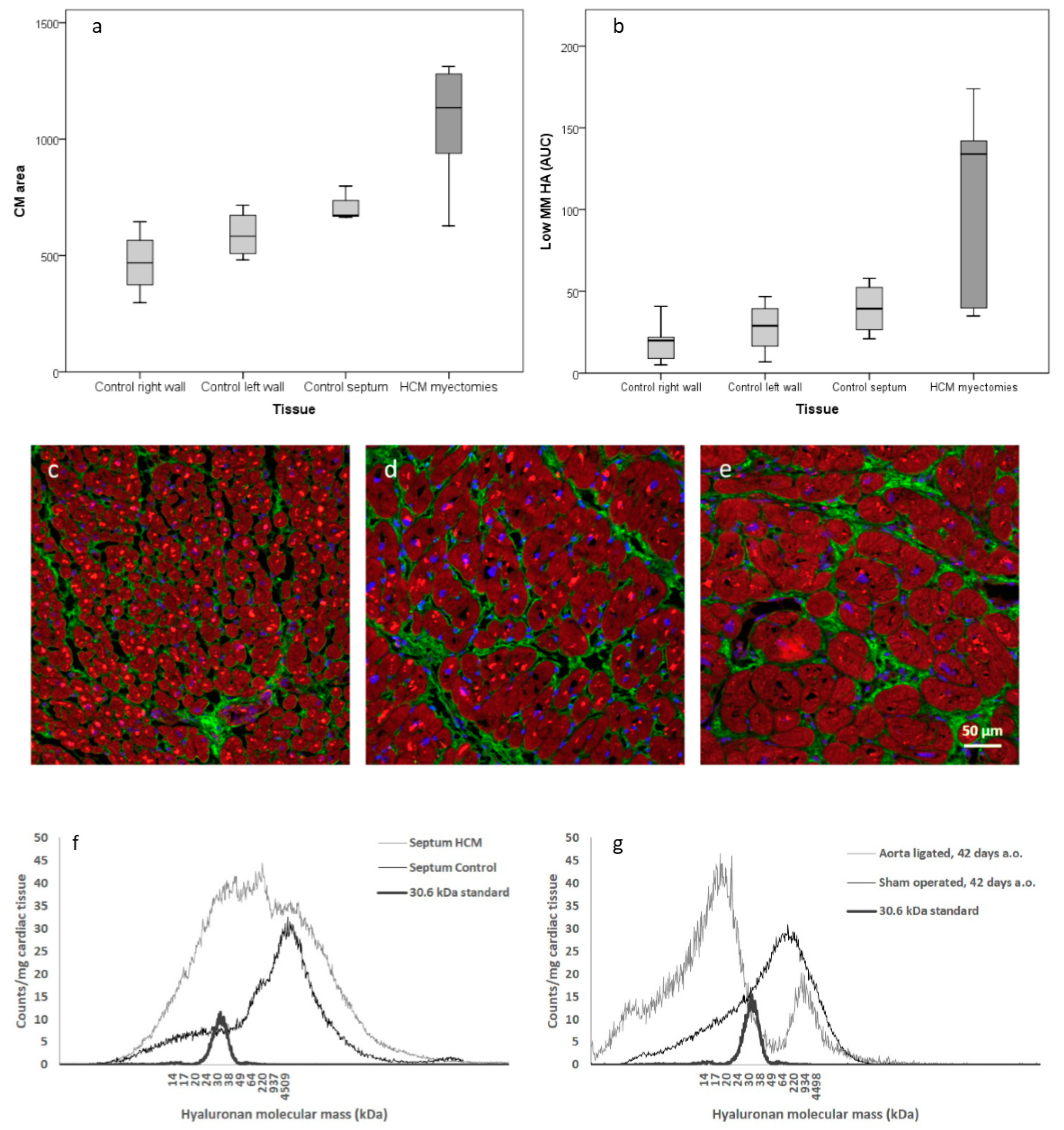
It has previously been shown that elevated levels of HA substrates enhance the synthesis of HA [
8] and our metabolic results could be confirmed with an increase of low MM HA in myectomies from HCM patients compared to non-failing hearts, as shown in
Figure 2f. An unexpected finding was a small but significant decrease of water content in cardiac tissue from HCM patients. Tissue accumulation of HA normally causes increased water content. Clinically human HCM is not associated with edema and all patients had end stage HCM. Possibly the low MM HA in the hypertrophic heart does not retain water in the tissue.
The factor analysis of expression level of genes involved in synthesis, degradation, and binding of HA clearly demonstrated that expression of genes involved in HA metabolism are altered in HCM. Interestingly, the expression levels of the three genes HYAL2, HAS2 and HAS3 correlated closely in the control left ventricle. Furthermore, levels of HYAL2 and HAS3 also correlated to amounts of low MM HA in the controls. In cardiac tissue from the HCM patients, these three genes showed significant changes in expression levels, both up and down regulated, but they had lost their mutual correlation and also to low MM HA. These clusters of gene expression levels might indicate genes involved in the same cellular process, governed by the same set of transcription factors. The pathological process of HCM induces another set of transcription factors causing different clusters of gene expression levels. This implies a common transcriptional regulation in the healthy heart which is disrupted during the development of HCM affecting the metabolism of HA, both synthesis and degradation. However, we cannot evaluate the functional significance of these gene expression clusters based on our results.
Downregulation of HYAL2 suggests that less HA is degraded and eliminated intracellularly. However, oxidative stress and excess production of ROS is a feature of HCM, which also has been implied in degrading HA into low MM HA. Glutathione has a protective function in the attenuation of ROS in HCM. In our metabolomic analysis glutamine and glutamate, substrates for glutathione, as well as glutathione itself were increased in cardiac tissue from the HCM patients, indicating a cellular response of high content of ROS. Thus, ROS might be a possible explanation for the fragmentation and accumulation of low MM HA in the hypertrophic heart.
Based on our results, a hypothetical explanation of the increased levels of low MM HA seen in the cardiac tissue from HCM patients might be increased availability of HA substrates from increased glucose metabolism, upregulated expression of HAS3, downregulation of internal degradation by HYAL2 with subsequent extracellular fragmentation by ROS.
We observed that the size of cardiomyocytes corresponded with the amount of low MM HA in human cardiac tissue, both in non-failing heart tissue as well as in basal septal myectomies from HCM patients, as shown in
Figure 2a–e. We experimentally confirmed accumulation of low MM HA in cardiac tissue from a model of induced cardiac hypertrophy in rat, showing a rapid shift from high MM HA to low MM HA after surgery, as shown in
Figure 2g,f. This suggests that fragmentation of HA into low MM HA occurs both in different parts of non-failing hearts and during the development of cardiac hypertrophy with levels corresponding to cardiomyocyte size.
An interesting parallel is found in experimental myocardial infarction research [
26]. When HA-based hydrogels with HA of different mass were injected in the infarcted area, the gel with the smallest mass HA (50 kDa) showed the most significant regeneration of myocardium and functional recovery. This further supports the hypothesis that low MM HA is not pathogenic in itself but possibly a part of a compensatory process initiated by the need of increased cardiac capacity.
An important feature in HCM is the development of arrhythmias including lethal arrhythmias. Increased fibrosis is known to disrupt the electrical conductivity between cardiomyocytes and act as a substrate for re-entrant arrhythmias [
27]. In our study, we observed an increased staining intensity of HA surrounding individual cardiomyocytes in cardiac tissues from HCM patients compared to healthy controls, as shown in
Figure 1c,f. Furthermore, it has been shown that high MM HA depolarizes the membrane potential in human fibroblasts, human embryonic kidney (HEK), and neurons in a concentration dependent manner which could be reversed by digestion of HA by hyaluronidase [
13]. Our observations and others results [
13] could indicate that a local increase of high MM HA in HCM may play a role in membrane polarizing of individual cardiomyocyte cell membranes.
The morphology of hypertrophic myocardium is characterized by regions of seemingly normal tissue, neighboring large disarrayed cardiomyocytes with a remodeled and expanded ECM. We identified regions within expanded ECM areas with stronger HA and weaker collagen staining and vice versa. Accumulation of HA has been shown to precede the development of fibrosis, and low MM HA has been predicted to have an essential role in promoting fibrosis [
12]. Low MM HA binds to the Toll-like receptor 2 (TLR2) and it has been shown that inhibition of TLR2 reduces cardiac fibrosis [
28,
29]. In addition, HA occurs in the myocardial infarction border zone [
30] and degradation of HA with hyaluronidases in early treatment of myocardial infarction has been shown to reduce fibrosis and infarct size [
31]. Possibly, areas with either more HA or collagen respectively mirrors different stages of progress of the disease and formation of fibrosis.
Both HA’s effect on cellular action potential and on the development of fibrosis suggests involvement of HA in development of arrhythmia. In addition, the increased amount of HA contributes to the expanded ECM, thus separating cardiomyocytes within myofibrils and disrupting their cell-to-cell connection and impulse conduction, which also could be a potential risk for arrhythmia.
Cardiac energy metabolism in HCM affects the heart in several ways on a molecular level, e.g., O-GlcNAcylation and mTOR activation. Here we have introduced changes in HA metabolism as another consequence of the dysregulated glucose metabolism.
The study is limited by the availability of human cardiac tissue. Cardiac tissue from five patients and five healthy control hearts are few in a statistical view but valuable material to support results from experimental models.
In conclusion, we have, in various steps, shown that both HA molecular mass and amount changes in the development of HCM. The return to fetal energy metabolism in the HCM heart causes an increased generation of substrates for HA, which together with an altered gene expression changes the metabolism of HA. HA might add to the risk of arrhythmias in HCM and the process of regulating cardiomyocyte size seems to involve fragmentation of HA into low MM HA. The connection of glucose metabolism to HCM and the heart needs further investigation since diet and disease, e.g., diabetes mellitus, can affect the cellular uptake of glucose. This is a novel addition to the underlying mechanisms of hypertrophic cardiomyopathy.






{kind=link}
{kind=link}
{kind=link}