Transcriptome-Wide Analysis of Human Chondrocyte Expansion on Synoviocyte Matrix



Abstract
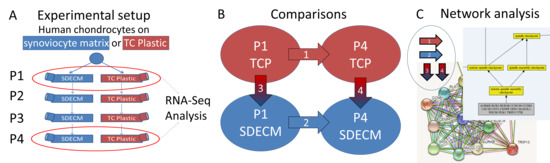
:
1. Introduction
2. Materials and Methods
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Murphy, L.B.; Cisternas, M.G.; Pasta, D.J.; Helmick, C.G.; Yelin, E.H. Medical Expenditures and Earnings Losses Among US Adults With Arthritis in 2013. Arthritis Care Res. 2018, 70, 869–876. [Google Scholar] [CrossRef] [PubMed]
- Giovannini, S.; Diaz-Romero, J.; Aigner, T.; Mainil-Varlet, P.; Nesic, D. Population doublings and percentage of S100-positive cells as predictors of in vitro chondrogenicity of expanded human articular chondrocytes. J. Cell. Physiol. 2010, 222, 411–420. [Google Scholar] [CrossRef] [PubMed]
- Kean, T.J.; Dennis, J.E. Synoviocyte Derived-Extracellular Matrix Enhances Human Articular Chondrocyte Proliferation and Maintains Re-Differentiation Capacity at Both Low and Atmospheric Oxygen Tensions. PLoS ONE 2015, 10, e0129961. [Google Scholar]
- Pei, M.; He, F. Extracellular matrix deposited by synovium-derived stem cells delays replicative senescent chondrocyte dedifferentiation and enhances redifferentiation. J. Cell. Physiol. 2012, 227, 2163–2174. [Google Scholar] [CrossRef] [PubMed]
- Everaert, C.; Luypaert, M.; Maag, J.L.V.; Cheng, Q.X.; Dinger, M.E.; Hellemans, J.; Mestdagh, P. Benchmarking of RNA-sequencing analysis workflows using whole-transcriptome RT-qPCR expression data. Sci Rep. 2017, 7, 1559. [Google Scholar] [CrossRef] [PubMed]
- Kean, T.J.; Koelewyn, S.; Ge, Z.; Li, Y.; Chen, R.; Dennis, J.E. Transcriptome-Wide Analysis of Human Chondrocyte Dedifferentiation on TC Plastic. In Proceedings of the Orthopedic Research Society Annual Meeting, Orlando, FL, USA, 5–8 March 2016. [Google Scholar]
- Langmead, B.; Trapnell, C.; Pop, M.; Salzberg, S.L. Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genome Biol. 2009, 10. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Pachter, L.; Salzberg, S.L. TopHat: Discovering splice junctions with RNA-Seq. Bioinformatics 2009, 25. [Google Scholar] [CrossRef] [PubMed]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing: Vienna, Austria, 2018. Available online: https://www.R-project.org/ (accessed on 9 December 2018).
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Morris, J.H.; Cook, H.; Kuhn, M.; Wyder, S.; Simonovic, M.; Santos, A.; Doncheva, N.T.; Roth, A.; Bork, P.; et al. The STRING database in 2017: quality-controlled protein-protein association networks, made broadly accessible. Nucleic Acids Res. 2017, 45. [Google Scholar] [CrossRef] [PubMed]
- Boyle, E.I.; Weng, S.; Gollub, J.; Jin, H.; Botstein, D.; Cherry, J.M.; Sherlock, G. GO::TermFinder—Open source software for accessing Gene Ontology information and finding significantly enriched Gene Ontology terms associated with a list of genes. Bioinformatics 2004, 20, 3710–3715. [Google Scholar] [CrossRef] [PubMed]
- Jax. Gene Ontology Browser. 2018. Available online: http://www.informatics.jax.org/vocab/gene_ontology/ (accessed on 8 November 2018).
- Marlovits, S.; Hombauer, M.; Truppe, M.; Vecsei, V.; Schlegel, W. Changes in the ratio of type-I and type-II collagen expression during monolayer culture of human chondrocytes. J. Bone Jt. Surg. J. 2004, 86, 286–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melrose, J.; Smith, S.; Cake, M.; Read, R.; Whitelock, J. Comparative spatial and temporal localisation of perlecan, aggrecan and type I.; II and IV collagen in the ovine meniscus: An ageing study. Histochem. Cell Biol. 2005, 124, 225–235. [Google Scholar] [CrossRef] [PubMed]
- Rosen, N.; Chalifa-Caspi, V.; Shmueli, O.; Adato, A.; Lapidot, M.; Stampnitzky, J.; Safran, M.; Lancet, D. GeneLoc: Exon-based integration of human genome maps. Bioinformatics 2003, 19, i222–i224. [Google Scholar] [CrossRef] [PubMed]
- Peffers, M.; Liu, X.; Clegg, P. Transcriptomic signatures in cartilage ageing. Arthritis Res. Ther. 2013, 15, R98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuzzocrea, S.; Mazzon, E.; Dugo, L.; Patel, N.S.; Serraino, I.; Di Paola, R.; Genovese, T.; Britti, D.; De Maio, M.; Caputi, A.P.; et al. Reduction in the evolution of murine type II collagen-induced arthritis by treatment with rosiglitazone, a ligand of the peroxisome proliferator-activated receptor gamma. Arthritis Rheum. 2003, 48, 3544–3556. [Google Scholar] [CrossRef]
- Xu, J.; Lv, S.; Hou, Y.; Xu, K.; Sun, D.; Zheng, Y.; Zhang, Z.; Li, X.; Li, Y.; Chi, G. miR-27b promotes type II collagen expression by targetting peroxisome proliferator-activated receptor-gamma2 during rat articular chondrocyte differentiation. Biosci. Rep. 2018, 38. [Google Scholar] [CrossRef]
- Liu, C.F.; Lefebvre, V. The transcription factors SOX9 and SOX5/SOX6 cooperate genome-wide through super-enhancers to drive chondrogenesis. Nucleic Acids Res. 2015, 43, 8183–8203. [Google Scholar] [CrossRef] [Green Version]
- De Crombrugghe, B.; Lefebvre, V.; Behringer, R.R.; Bi, W.; Murakami, S.; Huang, W. Transcriptional mechanisms of chondrocyte differentiation. Matrix Biol. 2000, 19, 389–394. [Google Scholar] [CrossRef]
- Cozzolino, A.M.; Noce, V.; Battistelli, C.; Marchetti, A.; Grassi, G.; Cicchini, C.; Tripodi, M.; Amicone, L. Modulating the Substrate Stiffness to Manipulate Differentiation of Resident Liver Stem Cells and to Improve the Differentiation State of Hepatocytes. Stem Cells Int. 2016, 2016, 5481493. [Google Scholar] [CrossRef] [PubMed]
- Recha-Sancho, L.; Moutos, F.T.; Abella, J.; Guilak, F.; Semino, C.E. Dedifferentiated Human Articular Chondrocytes Redifferentiate to a Cartilage-Like Tissue Phenotype in a Poly(epsilon-Caprolactone)/Self-Assembling Peptide Composite Scaffold. Materials 2016, 9, 472. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Gong, T.; Xie, J.; Lin, S.; Liu, Y.; Zhou, T.; Lin, Y. Softening Substrates Promote Chondrocytes Phenotype via RhoA/ROCK Pathway. ACS Appl. Mater. Interfaces 2016, 8, 22884–22891. [Google Scholar] [CrossRef] [PubMed]
- Callahan, L.A.; Ganios, A.M.; Childers, E.P.; Weiner, S.D.; Becker, M.L. Primary human chondrocyte extracellular matrix formation and phenotype maintenance using RGD-derivatized PEGDM hydrogels possessing a continuous Young’s modulus gradient. Acta Biomater. 2013, 9, 6095–6104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Efrat, S. Mechanisms of adult human beta-cell in vitro dedifferentiation and redifferentiation. Diabetes Obes. Metab. 2016, 18 (Suppl. 1), 97–101. [Google Scholar] [CrossRef] [PubMed]
- Barta, T.; Dolezalova, D.; Holubcova, Z.; Hampl, A. Cell cycle regulation in human embryonic stem cells: Links to adaptation to cell culture. Exp. Biol. Med. 2013, 238, 271–275. [Google Scholar] [CrossRef]
- Blagosklonny, M.V. Geroconversion: Irreversible step to cellular senescence. Cell Cycle 2014, 13, 3628–3635. [Google Scholar] [CrossRef]
- Blagosklonny, M.V. Cell cycle arrest is not senescence. Aging 2011, 3, 94–101. [Google Scholar] [CrossRef]
- Leontieva, O.V.; Blagosklonny, M.V. Gerosuppression in confluent cells. Aging 2014, 6, 1010–1018. [Google Scholar] [CrossRef] [Green Version]
- De Luna-Preitschopf, A.; Zwickl, H.; Nehrer, S.; Hengstschlager, M.; Mikula, M. Rapamycin Maintains the Chondrocytic Phenotype and Interferes with Inflammatory Cytokine Induced Processes. Int. J. Mol. Sci. 2017, 18, 1494. [Google Scholar] [CrossRef]
- Yan, B.; Zhang, Z.; Jin, D.; Cai, C.; Jia, C.; Liu, W.; Wang, T.; Li, S.; Zhang, H.; Huang, B.; et al. mTORC1 regulates PTHrP to coordinate chondrocyte growth, proliferation and differentiation. Nat. Commun. 2016, 7, 11151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Preitschopf, A.; Schorghofer, D.; Kinslechner, K.; Schutz, B.; Zwickl, H.; Rosner, M.; Joo, J.G.; Nehrer, S.; Hengstschlager, M.; Mikula, M. Rapamycin-Induced Hypoxia Inducible Factor 2A Is Essential for Chondrogenic Differentiation of Amniotic Fluid Stem Cells. Stem Cells Transl. Med. 2016, 5, 580–590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morales, T.I. Chondrocyte moves: Clever strategies? Osteoarthr. Cartil. 2007, 15, 861–871. [Google Scholar] [CrossRef] [PubMed]




 ) indicates that gene expression increased and a blue downward-pointing arrow (
) indicates that gene expression increased and a blue downward-pointing arrow (  ) that it decreased.
) indicates that gene expression increased and a blue downward-pointing arrow ( ) that it decreased.
) that it decreased.
) indicates that gene expression increased and a blue downward-pointing arrow ( ) that it decreased. ) indicates that gene expression increased and a blue downward-pointing arrow ( ) that it decreased.
) indicates that gene expression increased and a blue downward-pointing arrow ( ) that it decreased.
) indicates that gene expression increased and a blue downward-pointing arrow ( ) that it decreased.
) indicates that gene expression increased and a blue downward-pointing arrow ( ) that it decreased.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Comparison | Upregulated (%) | Downregulated (%) | Total | Common 3 |
|---|---|---|---|---|
| (1) P1 vs. P4 on TCP 1 | 404 (39) | 620 (61) | 1024 | 512 |
| (2) P1 vs. P4 on SDECM 1 | 316 (41) | 447 (59) | 763 | |
| (3) TCP vs. SDECM P1 2 | 151 (46) | 180 (54) | 331 | 107 |
| (4) TCP vs. SDECM P4 2 | 162 (48) | 177 (52) | 339 |
| Comparison 1 | Comparison 2 | ||||
|---|---|---|---|---|---|
| Gene Symbol | Gene Name | FC 1 | Gene Symbol | Gene Name | FC 1 |
| MAFB | MAF BZIP transcription factor B | 25.5 D | APLN | Apelin | 21.8 D |
| PLK1 | Polo-like kinase 1 | 23.1 U | MMP1 | Matrix metallopeptidase 1 | 26.6 D |
| CENPF | Centromere protein F | 12.0 U | KIF20A | Kinesin family member 20A | 27.6 U |
| SLC40A1 | Solute carrier family 40 member 1 | 166. D | TOP2A | DNA topoisomerase II alpha | 16.3 U |
| TOP2A | DNA topoisomerase II alpha | 13.0 U | CDC20 | Cell division cycle 20 | 21.1 U |
| MKI67 | Marker of proliferation Ki-67 | 14.5 U | TPX2 | TPX2, microtubule nucleation factor | 16.0 U |
| CCNB1 | Cyclin B1 | 15.0 U | FOXM1 | Forkhead box M1 | 12.3 U |
| FGFR2 | Fibroblast growth factor receptor 2 | 19.0 D | BIRC5 | Baculoviral IAP repeat containing 5 | 19.8 U |
| ISM1 | Isthmin 1 | 25.5 D | MKI67 | Marker of proliferation Ki-67 | 21.4 U |
| CDC20 | Cell division cycle 20 | 19.6 U | PRC1 | Protein regulator of cytokinesis 1 | 12.6 U |
| PRC1 | Protein regulator of cytokinesis 1 | 12.1 U | DLGAP5 | DL-associated protein 5 | 23.6 U |
| A2M | Alpha-2-macroglobulin | 21.9 D | PLK1 | Polo-like kinase 1 | 17.7 U |
| HMMR | Hyaluronan-mediated motility receptor | 20.9 U | SLC40A1 | Solute carrier family 40 member 1 | 25.9 D |
| DLGAP5 | DLG-associated protein 5 | 17.1 U | ASPM | Abnormal spindle microtubule assembly | 10.9 U |
| AURKA | Aurora kinase A | 20.0 U | CCNB1 | Cyclin B1 | 11.7 U |
| CCNB2 | Cyclin B2 | 17.2 U | PRR11 | Proline rich 11 | 9.43 U |
| TPX2 | TPX2, microtubule nucleation factor | 14.2 U | CENPF | Centromere protein F | 12.6 U |
| STEAP4 | STEAP4 metalloreductase | 14.1 D | KIF23 | Kinesin family member 23 | 13.6 U |
| ELN | Elastin | 7.66 U | CEP55 | Centrosomal protein 55 | 14.6 U |
| PRR11 | Proline rich 11 | 11.9 U | ANLN | Anillin actin binding protein | 12.2 U |
| Comparison 3 | Comparison 4 | ||||
|---|---|---|---|---|---|
| Gene Symbol | Gene Name | FC 1 | Gene Symbol | Gene Name | FC 1 |
| POSTN | Periostin | 16.9 U | COLEC12 | Collectin subfamily member 12 | 32.3 U |
| COMP | Cartilage oligomeric matrix protein | 28.5 D | MME | Membrane metalloendopeptidase | 9.11 U |
| CLEC3B | C-type lectin domain family 3 member B | 14.7 D | CRLF1 | Cytokine receptor-like factor 1 | 7.62 D |
| DCN | Decorin | 5.87 D | MFAP5 | Tetratricopeptide repeat domain 9 | 14.5 D |
| PODN | Podocan | 8.89 D | TTC9 | Microfibril-associated protein 5 | 12.1 D |
| CTGF | Connective tissue growth factor | 5.52 D | IGFBP1 | Insulin-like growth factor binding protein 1 | 11.9 D |
| ADAMTS5 | ADAM metallopeptidase thrombospondin type 1 motif 5 | 9.99 D | CLEC3B | C-type lectin domain family 3 member B | 6.48 D |
| PPAP2B | Phospholipid phosphatase 3 | 5.57 D | DSP | Desmoplakin | 10.1 D |
| TAGLN | Transgelin | 7.23 D | LOX | Lysyl oxidase | 4.02 D |
| SFRP4 | Secreted frizzled-related protein 4 | 9.26 D | IGFBP5 | Insulin-like growth factor binding protein 5 | 4.61 D |
| CAMK2N1 | Calcium/calmodulin-dependent protein kinase II inhibitor 1 | 11.3 U | C6orf132 | Chromosome 6 open reading frame 132 | 9.53 D |
| FHL1 | Four and a half LIM domains 1 | 5.66 D | MOXD1 | Monooxygenase DBH-like 1 | 7.35 U |
| OMD | Osteomodulin | 4.92 D | CAMK2N1 | Calcium/calmodulin-dependent protein kinase II inhibitor 1 | 20.1 U |
| COL18A1 | Collagen type XVIII alpha 1 chain | 6.22 U | PHLDA1 | Pleckstrin homology-like domain family A Member 1 | 4.67 U |
| SEMA5A | Semaphorin 5A | 4.22 U | EMB | Embigin | 6.48 U |
| SCD | Stearoyl-CoA desaturase | 5.78 U | PITPNM3 | PITPNM family member 3 | 4.76 D |
| MYL9 | Myosin light chain 9 | 6.15 D | INHBB | Inhibin subunit beta B | 10.0 D |
| CPM | Carboxypeptidase M | 6.23 U | ACTA2 | Actin, alpha 2, smooth muscle, aorta | 4.66 D |
| ITIH5 | Inter-alpha-trypsin inhibitor heavy chain family member 5 | 6.58 D | LOC100505633 | Long intergenic non-protein coding RNA 1133 | 5.19 D |
| AP1S3 | Adaptor-related protein complex 1 subunit sigma 3 | 11.3 D | FOXQ1 | Forkhead box Q1 | 4.96 U |
| Gene Symbol | Gene Name | TCP FC | TCP padj | SDECM FC | SDECM padj |
|---|---|---|---|---|---|
| BRCA1 | BRCA1, DNA repair associated | 4.60 | 3.9 × 10−38 | 6.54 | 2.1 × 10−31 |
| CEBPA | CCAAT enhancer binding protein alpha | 2.11 | 2.7 × 10−1 | 4.82 | 8.0 × 10−4 |
| E2F8 | E2F transcription factor 8 | 8.16 | 1.1 × 10−22 | 21.21 | 3.2 × 10−22 |
| EZH2 | Enhancer of zeste 2 polycomb repressive complex 2 subunit | 5.76 | 1.5 × 10−46 | 3.87 | 1.1 × 10−18 |
| FOSL1 | FOS-like 1, AP-1 transcription factor subunit | 4.82 | 5.0 × 10−43 | 1.10 | 6.5 × 10−1 |
| MXD3 | MAX dimerization protein 3 | 4.79 | 1.8 ×10−24 | 4.43 | 2.7 × 10−11 |
| Gene Symbol | Gene Name | Fold Change | padj |
|---|---|---|---|
| FOSL1 | FOS Like 1, AP-1 transcription factor subunit | 4.81 U | 5.0 × 10−43 |
| STC1 | Stanniocalcin 1 | 4.73 D | 2.2 × 10−40 |
| CDH2 | Cadherin 2 | 11.7 U | 1.0 × 10−27 |
| NXPH3 | Neurexophilin 3 | 4.46 D | 7.1 × 10−19 |
| POM121L9P | POM121 transmembrane nucleoporin-like 9, pseudogene | 10.9 D | 9.6 × 10−19 |
| MSC | Musculin | 4.62 U | 4.3 × 10−12 |
| POSTN | Periostin | 7.64 U | 1.2 × 10−11 |
| CCRL1 | Atypical chemokine receptor 4 | 8.45 D | 4.7 × 10−10 |
| ADAMTS14 | ADAM metallopeptidase with thrombospondin type 1 motif 14 | 4.60 U | 8.6 × 10−10 |
| ACSS1 | Acyl-CoA synthetase short chain family member 1 | 4.24 D | 1.5 × 10−8 |
| SH3TC2 | SH3 domain and tetratricopeptide repeats 2 | 5.20 U | 1.1 × 10−6 |
| NTF3 | Neurotrophin 3 | 4.00 U | 2.1 × 10−6 |
| MYBPH | Myosin binding protein H | 6.36 D | 1.2 × 10−5 |
| ELOVL2 | ELOVL fatty acid elongase 2 | 9.81 U | 1.2 × 10−4 |
| GRIK5 | Glutamate ionotropic receptor kainate type subunit 5 | 4.46 D | 9.2 × 10−3 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kean, T.J.; Ge, Z.; Li, Y.; Chen, R.; Dennis, J.E. Transcriptome-Wide Analysis of Human Chondrocyte Expansion on Synoviocyte Matrix. Cells 2019, 8, 85. https://doi.org/10.3390/cells8020085
Kean TJ, Ge Z, Li Y, Chen R, Dennis JE. Transcriptome-Wide Analysis of Human Chondrocyte Expansion on Synoviocyte Matrix. Cells. 2019; 8(2):85. https://doi.org/10.3390/cells8020085
Chicago/Turabian StyleKean, Thomas J., Zhongqi Ge, Yumei Li, Rui Chen, and James E. Dennis. 2019. "Transcriptome-Wide Analysis of Human Chondrocyte Expansion on Synoviocyte Matrix" Cells 8, no. 2: 85. https://doi.org/10.3390/cells8020085
APA StyleKean, T. J., Ge, Z., Li, Y., Chen, R., & Dennis, J. E. (2019). Transcriptome-Wide Analysis of Human Chondrocyte Expansion on Synoviocyte Matrix. Cells, 8(2), 85. https://doi.org/10.3390/cells8020085