The Adenosine A2B Receptor Drives Osteoclast-Mediated Bone Resorption in Hypoxic Microenvironments

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Ethics
2.2. Osteoclast Differentiation and Cell Culture
2.3. Osteoclast Formation and Activity Assays
2.4. Transfection with siRNA and Luciferase Assays
2.5. Realtime PCR
2.6. Western Blotting
2.7. Metabolic Assays
2.8. ELISAs
2.9. Immunostaining
2.10. Statistical Methods
3. Results
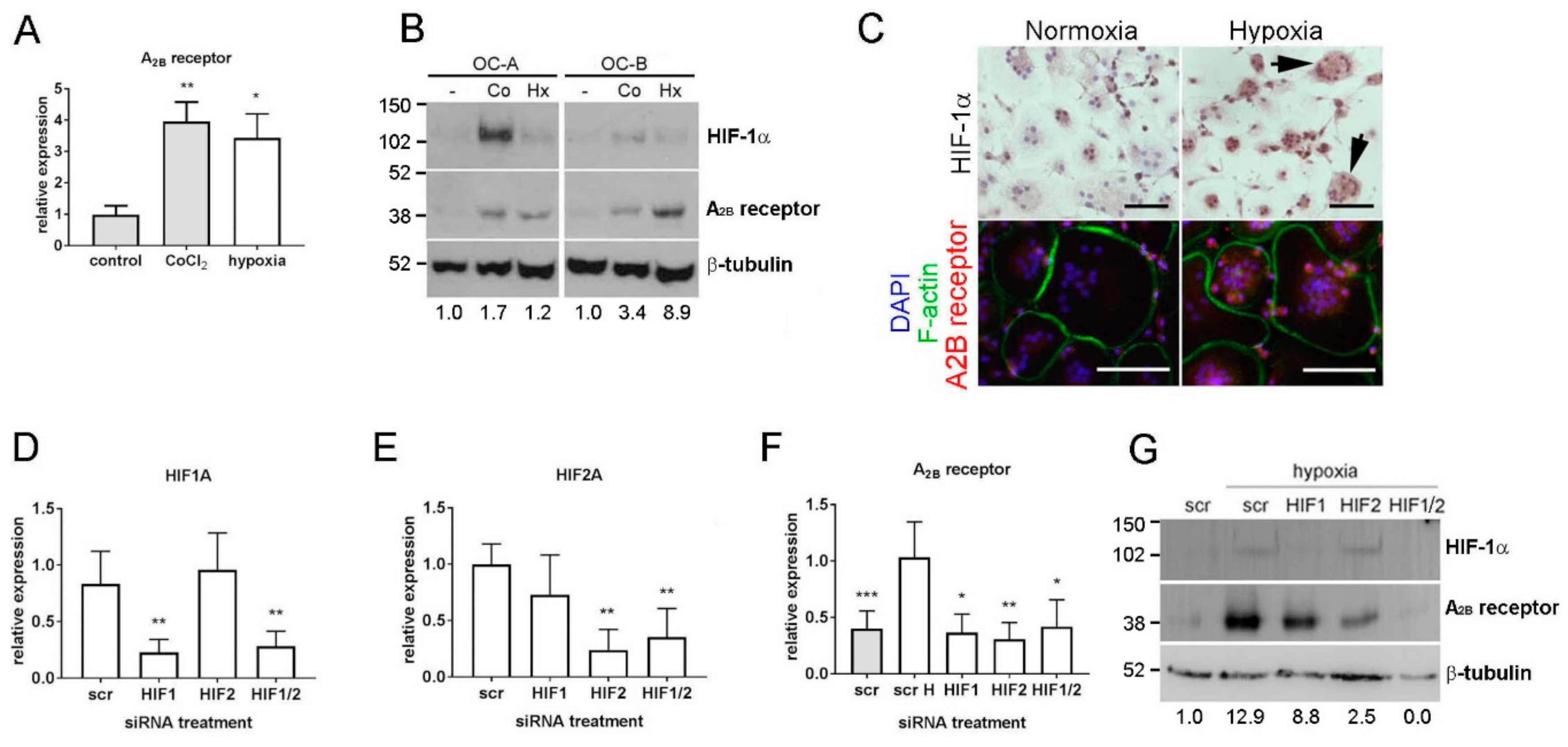
3.1. Hypoxic Induction of the A2B Receptor is HIF-Regulated in Osteoclasts
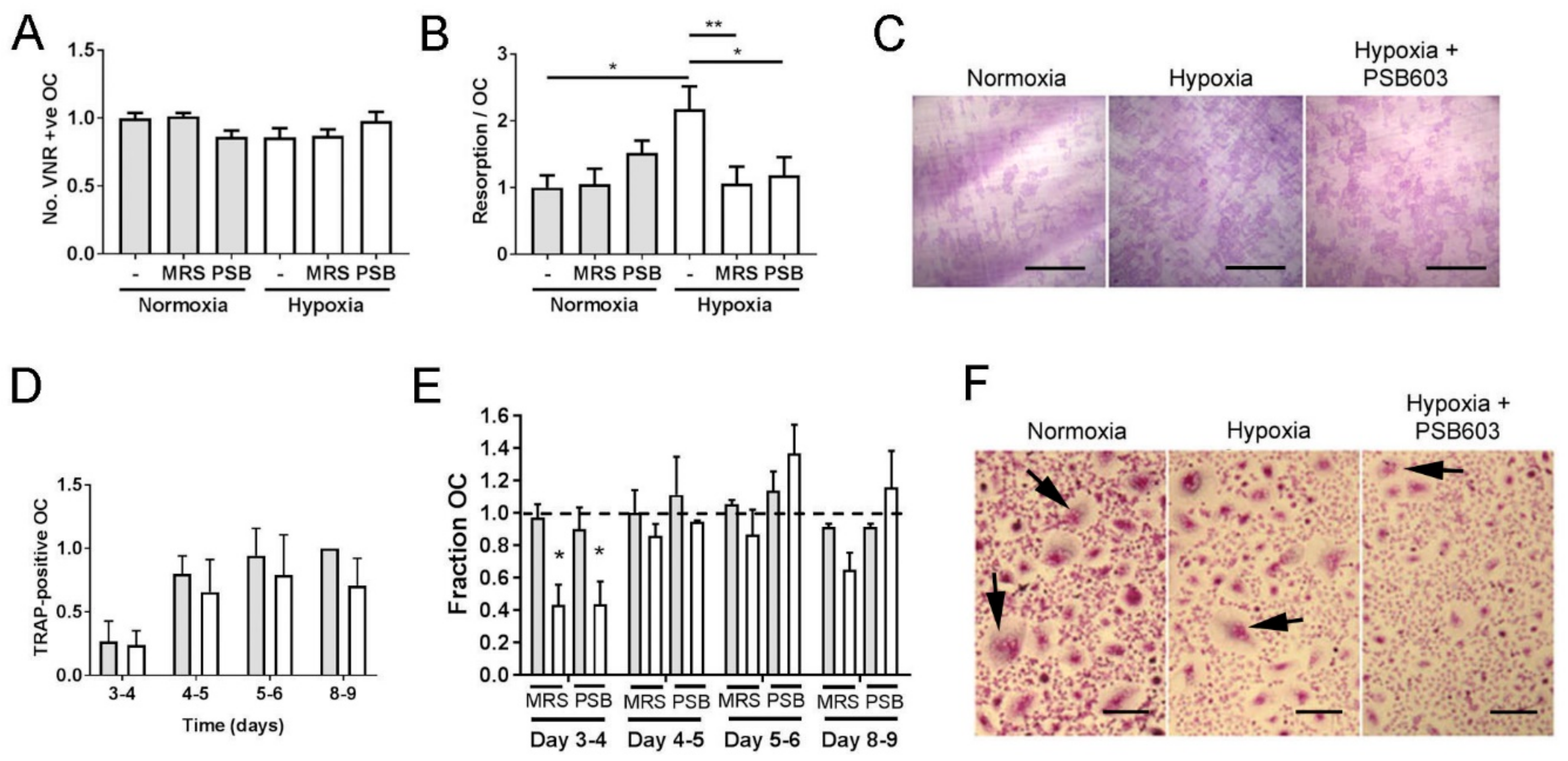
3.2. The A2B Receptor Drives the Hypoxic Increase in Osteoclast Bone Resorption
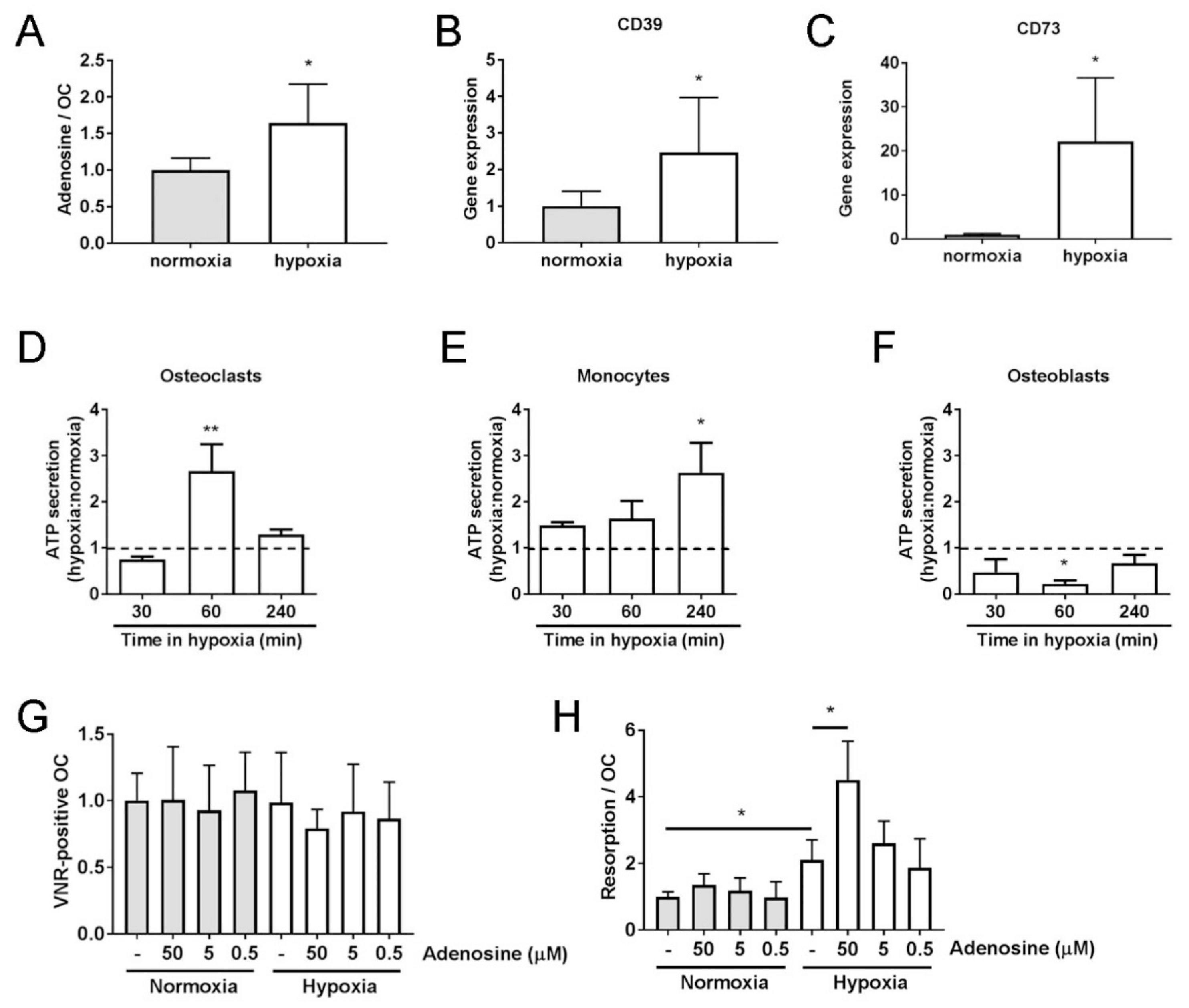
3.3. Hypoxic Osteoclasts Secrete ATP to Drive Increased Adenosine Concentrations
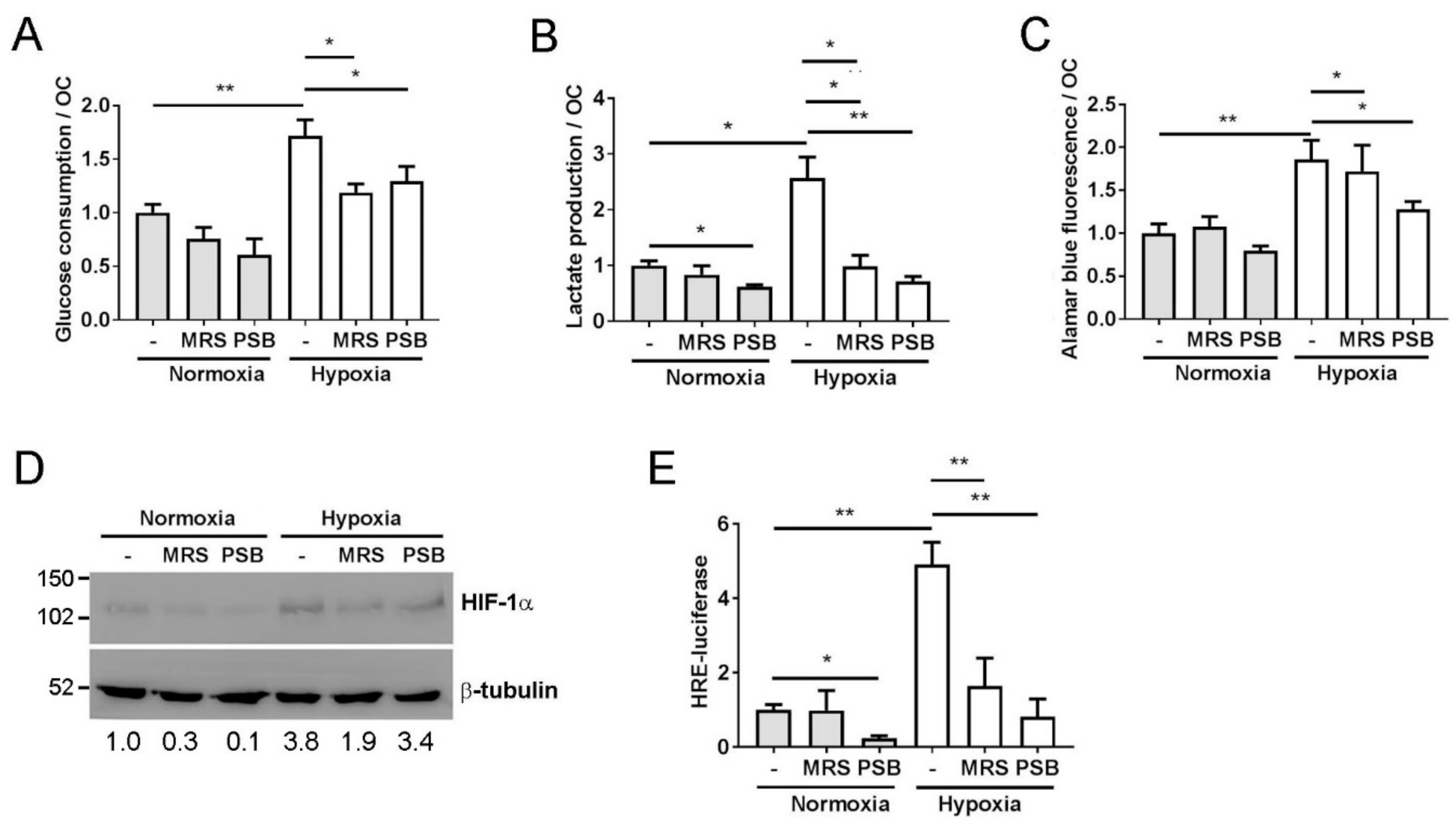
3.4. The A2B Receptor Activates Glycolytic Pathways
4. Discussion
5. Conclusions
Supplementary Materials
Funding
Acknowledgments
Conflicts of Interest
References
- Kaarela, K. Prognostic Factors and Diagnostic Criteria in Early Rheumatoid Arthritis. Scand. J. Rheumatol. Suppl. 1985, 57, 1–54. [Google Scholar] [CrossRef] [PubMed]
- Harrison, B.J.; Hutchinson, C.E.; Adams, J.; Bruce, I.N.; Herrick, A.L. Assessing Periarticular Bone Mineral Density in Patients with Early Psoriatic Arthritis or Rheumatoid Arthritis. Ann. Rheum. Dis. 2002, 61, 1007–1011. [Google Scholar] [CrossRef] [PubMed]
- Gough, A.; Sambrook, P.; Devlin, J.; Huissoon, A.; Njeh, C.; Robbins, S.; Nguyen, T.; Emery, P. Osteoclastic Activation Is the Principal Mechanism Leading to Secondary Osteoporosis in Rheumatoid Arthritis. J. Rheumatol. 1998, 25, 1282–1289. [Google Scholar] [PubMed]
- Fujikawa, Y.; Sabokbar, A.; Neale, S.; Athanasou, N.A. Human Osteoclast Formation and Bone Resorption by Monocytes and Synovial Macrophages in Rheumatoid Arthritis. Ann. Rheum. Dis. 1996, 55, 816–822. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, H.; Shima, N.; Nakagawa, N.; Yamaguchi, K.; Kinosaki, M.; Mochizuki, S.; Tomoyasu, A.; Yano, K.; Goto, M.; et al. Osteoclast Differentiation Factor Is a Ligand for Osteoprotegerin/Osteoclastogenesis-Inhibitory Factor and Is Identical to Trance/Rankl. Proc. Natl. Acad. Sci. USA 1998, 95, 3597–3602. [Google Scholar] [CrossRef] [PubMed]
- Knowles, H.J. Hypoxic Regulation of Osteoclast Differentiation and Bone Resorption Activity. Hypoxia 2015, 3, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Hulley, P.A.; Bishop, T.; Vernet, A.; Schneider, J.E.; Edwards, J.R.; Athanasou, N.A.; Knowles, H.J. Hypoxia-Inducible Factor 1-Alpha Does Not Regulate Osteoclastogenesis but Enhances Bone Resorption Activity Via Prolyl-4-Hydroxylase 2. J. Pathol. 2017, 242, 322–333. [Google Scholar] [CrossRef] [PubMed]
- Orriss, I.R. The Role of Purinergic Signalling in the Musculoskeletal System. Auton. Neurosci. 2015, 191, 124–134. [Google Scholar] [CrossRef]
- Kara, F.M.; Chitu, V.; Sloane, J.; Axelrod, M.; Fredholm, B.B.; Stanley, E.R.; Cronstei, B.N. Adenosine A1 Receptors (A1rs) Play a Critical Role in Osteoclast Formation and Function. FASEB J. 2010, 24, 2325–2333. [Google Scholar] [CrossRef]
- Kara, F.M.; Doty, S.B.; Boskey, A.; Goldring, S.; Zaidi, M.; Fredholm, B.B.; Cronstein, B.N. Adenosine a(1) Receptors Regulate Bone Resorption in Mice: Adenosine a(1) Receptor Blockade or Deletion Increases Bone Density and Prevents Ovariectomy-Induced Bone Loss in Adenosine a(1) Receptor-Knockout Mice. Arthritis Rheum. 2010, 62, 534–541. [Google Scholar] [CrossRef]
- Pellegatti, P.; Falzoni, S.; Donvito, G.; Lemaire, I.; Di Virgilio, F. P2x7 Receptor Drives Osteoclast Fusion by Increasing the Extracellular Adenosine Concentration. FASEB J. 2011, 25, 1264–1274. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Cronstein, B.N. Adenosine A1 Receptor Regulates Osteoclast Formation by Altering Traf6/Tak1 Signaling. Purinergic Signal. 2012, 8, 327–337. [Google Scholar] [CrossRef] [PubMed]
- Mediero, A.; Frenkel, S.R.; Wilder, T.; He, W.; Mazumder, A.; Cronstein, B.N. Adenosine A2a Receptor Activation Prevents Wear Particle-Induced Osteolysis. Sci. Transl. Med. 2012. [Google Scholar] [CrossRef] [PubMed]
- Mediero, A.; Kara, F.M.; Wilder, T.B.; Cronstein, N. Adenosine a(2a) Receptor Ligation Inhibits Osteoclast Formation. Am. J. Pathol. 2012, 180, 775–786. [Google Scholar] [CrossRef] [PubMed]
- Hajjawi, M.O.; Patel, J.J.; Corcelli, M.; Arnett, T.R.; Orriss, I.R. Lack of Effect of Adenosine on the Function of Rodent Osteoblasts and Osteoclasts in Vitro. Purinergic Signal. 2016, 12, 247–258. [Google Scholar] [CrossRef] [PubMed]
- Morrison, L.; Turin, M.S.; King, B.F.; Burnstock, G.; Arnett, T.R. Atp Is a Potent Stimulator of the Activation and Formation of Rodent Osteoclasts. J. Physiol. 1998, 511, 495–500. [Google Scholar] [CrossRef] [PubMed]
- Orriss, I.R.; Knight, G.E.; Utting, J.C.; Taylor, S.E.; Burnstock, G.; Arnett, T.R. Hypoxia Stimulates Vesicular Atp Release from Rat Osteoblasts. J. Cell Physiol. 2009, 220, 155–162. [Google Scholar] [CrossRef]
- Strazzulla, L.C.; Cronstein, B.N. Regulation of Bone and Cartilage by Adenosine Signaling. Purinergic Signal. 2016, 12, 583–593. [Google Scholar] [CrossRef]
- Knowles, H.J.; Cleton-Jansen, A.M.; Korsching, E.; Athanasou, N.A. Hypoxia-Inducible Factor Regulates Osteoclast-Mediated Bone Resorption: Role of Angiopoietin-Like 4. FASEB J. 2010, 24, 4648–4659. [Google Scholar] [CrossRef]
- Sun, Y.; Huang, P. Huang. Adenosine A2b Receptor: From Cell Biology to Human Diseases. Front. Chem. 2016, 4, 37. [Google Scholar] [CrossRef]
- Teramachi, J.; Kukita, A.; Li, Y.J.; Ushijima, Y.; Ohkuma, H.; Wada, N.; Watanabe, T.; Nakamura, S.; Kukita, T. Adenosine Abolishes Mtx-Induced Suppression of Osteoclastogenesis and Inflammatory Bone Destruction in Adjuvant-Induced Arthritis. Lab. Invest. 2011, 91, 719–731. [Google Scholar] [CrossRef] [PubMed]
- Ye, T.S.; Du, Z.H.; Li, Z.H.; Xie, W.X.; Huang, K.T.; Chen, Y.; Chen, Z.Y.; Hu, H.; Wang, J.L.; Fang, J.Q. Repeated Electroacupuncture Persistently Elevates Adenosine and Ameliorates Collagen-Induced Arthritis in Rats. Evid. Based Complement. Alternat. Med. 2016. [Google Scholar] [CrossRef] [PubMed]
- Stamp, L.K.; Hazlett, J.; Roberts, R.L.; Frampton, C.; Highton, J.; Hessian, P.A. Adenosine Receptor Expression in Rheumatoid Synovium: A Basis for Methotrexate Action. Arthritis Res. Ther. 2012, 14, R138. [Google Scholar] [CrossRef]
- Houlder, E.L.; Millier, M.J.; Highton, J.; Gwynne-Jones, D.; Stamp, L.K.; Hessian, P.A. Expression of the Genes Facilitating Methotrexate Action within Subcutaneous Rheumatoid Nodules. Clin. Exp. Rheumatol. 2017, 35, 943–947. [Google Scholar] [PubMed]
- Jones, D.T.; Harris, A.L. Identification of Novel Small-Molecule Inhibitors of Hypoxia-Inducible Factor-1 Transactivation and DNA Binding. Mol. Cancer Ther. 2006, 5, 2193–2202. [Google Scholar] [CrossRef] [PubMed]
- Knowles, H.J.; Athanasou, N.A. Acute Hypoxia and Osteoclast Activity: A Balance between Enhanced Resorption and Increased Apoptosis. J. Pathol. 2009, 218, 256–264. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Yu, R.; Zhao, J.; Sun, L.; Jian, L.; Li, C.; Liu, X. Constant Hypoxia Inhibits Osteoclast Differentiation and Bone Resorption by Regulating Phosphorylation of Jnk and Ikappabalpha. Inflamm. Res. 2019, 68, 157–166. [Google Scholar] [CrossRef]
- Brandao-Burch, A.; Key, J.; Patel, M.L.; Arnett, T.R.; Orriss, I.R. The P2x7 Receptor Is an Important Regulator of Extracellular Atp Levels. Front. Endocrinol. 2012. [Google Scholar] [CrossRef]
- Sitaraman, S.V.; Wang, L.; Wong, M.; Bruewer, M.; Hobert, M.; Yun, C.H.; Merlin, D.; Madara, J.L. The Adenosine 2b Receptor Is Recruited to the Plasma Membrane and Associates with E3karp and Ezrin Upon Agonist Stimulation. J. Biol. Chem. 2002, 277, 33188–33195. [Google Scholar] [CrossRef]
- Sun, Y.; Hu, W.; Yu, X.; Liu, Z.; Tarran, R.; Ravid, K.; Huang, P. Actinin-1 Binds to the C-Terminus of A2b Adenosine Receptor (A2bar) and Enhances A2bar Cell-Surface Expression. Biochem. J. 2016, 473, 2179–2186. [Google Scholar] [CrossRef]
- Hu, X.; Adebiyi, M.G.; Luo, J.; Sun, K.; Le, T.T.; Zhang, Y.; Wu, H.; Zhao, S.; Karmouty-Quintana, H.; et al. Sustained Elevated Adenosine Via Adora2b Promotes Chronic Pain through Neuro-Immune Interaction. Cell Rep. 2016, 16, 106–119. [Google Scholar] [CrossRef]
- Ning, C.; Wen, Y.; Zhang, Y.; Dai, W.; Wang, W.; Zhang, L.; Qi, A.; Grenz, H.K.; Eltzschig, M.R.; et al. Excess Adenosine A2b Receptor Signaling Contributes to Priapism through Hif-1alpha Mediated Reduction of Pde5 Gene Expression. FASEB J. 2014, 28, 2725–2735. [Google Scholar] [CrossRef]
- Eckle, T.; Hartmann, K.; Bonney, S.; Reithel, S.; Mittelbronn, M.; Walker, L.A.; Lowes, B.D.; Han, J.; Borchers, C.H.; Buttrick, P.M.; et al. Adora2b-Elicited Per2 Stabilization Promotes a Hif-Dependent Metabolic Switch Crucial for Myocardial Adaptation to Ischemia. Nat. Med. 2012, 18, 774–782. [Google Scholar] [CrossRef] [PubMed]
- Morten, K.J.; Badder, L.; Knowles, H.J. Differential Regulation of Hif-Mediated Pathways Increases Mitochondrial Metabolism and Atp Production in Hypoxic Osteoclasts. J. Pathol. 2013, 229, 755–764. [Google Scholar] [CrossRef]
- Arnett, T.R.; Orriss, I.R. Metabolic Properties of the Osteoclast. Bone 2018, 115, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Okabe, T.M.; Kumagai, Y.; Nakajima, S.; Shirotake, K.; Kodaira, M.; Oyama, M.U.; Ikeda, M. The Impact of Hif1alpha on the Per2 Circadian Rhythm in Renal Cancer Cell Lines. PLoS ONE 2014. [Google Scholar] [CrossRef] [PubMed]
- Poth, J.M.; Brodsky, K.; Ehrentraut, H.; Grenz, A.; Eltzschig, H.K. Transcriptional Control of Adenosine Signaling by Hypoxia-Inducible Transcription Factors During Ischemic or Inflammatory Disease. J. Mol. Med. 2013, 91, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Eckle, T.; Kewley, E.M.; Brodsky, K.; Tak, E.; Bonney, S.; Gobel, M.; Anderson, D.; Glover, L.E.; Riegel, A.K.; Colgan, S.P.; et al. Eltzschig. Identification of Hypoxia-Inducible Factor Hif-1a as Transcriptional Regulator of the A2b Adenosine Receptor During Acute Lung Injury. J. Immunol. 2014, 192, 1249–1256. [Google Scholar] [CrossRef]
- Lan, J.; Lu, H.; Samanta, D.; Salman, S.; Lu, Y.; Semenza, G.L. Hypoxia-Inducible Factor 1-Dependent Expression of Adenosine Receptor 2b Promotes Breast Cancer Stem Cell Enrichment. Proc. Natl. Acad. Sci. USA 2018. [Google Scholar] [CrossRef]
- Yang, M.; Ma, C.; Liu, S.; Shao, Q.; Gao, W.; Song, B.; Sun, J.; Xie, Q.; Zhang, Y.; Feng, A.; et al. Hif-Dependent Induction of Adenosine Receptor A2b Skews Human Dendritic Cells to a Th2-Stimulating Phenotype under Hypoxia. Immunol. Cell Biol. 2010, 88, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.H.; Oh, J.H.; Lee, N.K. The Inactivation of Erk1/2, P38 and Nf-Kb Is Involved in the Down-Regulation of Osteoclastogenesis and Function by A2b Adenosine Receptor Stimulation. Mol. Cells 2017, 40, 752–760. [Google Scholar]
- Cui, D.; Trier, K.; Chen, X.; Zeng, J.; Yang, X.; Hu, J.; Ge, J. Distribution of Adenosine Receptors in Human Sclera Fibroblasts. Mol. Vis. 2008, 14, 523–529. [Google Scholar] [PubMed]
- Wan, W.J.; Cui, D.M.; Yang, X.; Hu, J.M.; Li, C.X.; Hu, S.L.; Trier, K.; Zeng, J.W. Expression of Adenosine Receptors in Human Retinal Pigment Epithelium Cells in Vitro. Chin. Med. J. 2011, 124, 1139–1144. [Google Scholar] [PubMed]
- Merrill, J.T.; Shen, C.; Schreibman, D.; Coffey, D.; Zakharenko, O.; Fisher, R.; Lahita, R.G.; Salmon, J.; Cronstein, B.N. Adenosine A1 Receptor Promotion of Multinucleated Giant Cell Formation by Human Monocytes: A Mechanism for Methotrexate-Induced Nodulosis in Rheumatoid Arthritis. Arthritis Rheum. 1997, 40, 1308–1315. [Google Scholar] [PubMed]
- Molck, C.; Ryall, J.; Failla, L.M.; Coates, J.L.; Pascussi, J.M.; Heath, J.K.; Stewart, G.; Hollande, F. The A2b Adenosine Receptor Antagonist Psb-603 Promotes Oxidative Phosphorylation and Ros Production in Colorectal Cancer Cells Via Adenosine Receptor-Independent Mechanism. Cancer Lett. 2016, 383, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Corciulo, C.; Wilder, T.; Cronstein, B.N. Adenosine A2b Receptors Play an Important Role in Bone Homeostasis. Purinergic Signal. 2016, 12, 537–547. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Mazumder, A.; Wilder, T.; Cronstein, B.N. Adenosine Regulates Bone Metabolism Via A1, A2a, and A2b Receptors in Bone Marrow Cells from Normal Humans and Patients with Multiple Myeloma. FASEB J. 2013, 27, 3446–3454. [Google Scholar] [CrossRef] [PubMed]
- Hinz, S.; Lacher, S.K.; Seibt, B.F.; Muller, C.F. Bay60-6583 Acts as a Partial Agonist at Adenosine A2b Receptors. J. Pharmacol. Exp. Ther. 2014, 349, 427–436. [Google Scholar] [CrossRef] [PubMed]
- Majmundar, A.J.; Wong, W.J.; Simon, M.C. Hypoxia-Inducible Factors and the Response to Hypoxic Stress. Mol. Cell 2010, 40, 294–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dammen, R.; Haugen, M.; Svejda, B.; Alaimo, D.; Brenna, O.; Pfragner, R.; Gustafsson, B.I.; Kidd, M. The Stimulatory Adenosine Receptor Adora2b Regulates Serotonin (5-Ht) Synthesis and Release in Oxygen-Depleted Ec Cells in Inflammatory Bowel Disease. PLoS ONE 2013. [Google Scholar] [CrossRef]
- Kasama, H.; Sakamoto, Y.; Kasamatsu, A.; Okamoto, A.; Koyama, T.; Minakawa, Y.; Ogawara, K.; Yokoe, H.; Shiiba, M.; Tanzawa, H.; et al. Adenosine A2b Receptor Promotes Progression of Human Oral Cancer. BMC Cancer 2015. [Google Scholar] [CrossRef] [PubMed]
- Merighi, S.; Borea, P.A.; Stefanelli, A.; Bencivenni, S.; Castillo, C.A.; Varani, K.; Gessi, S. A2a and A2b Adenosine Receptors Affect Hif-1alpha Signaling in Activated Primary Microglial Cells. Glia 2015, 63, 1933–1952. [Google Scholar] [CrossRef] [PubMed]
- Quarona, V.; Ferri, V.; Chillemi, A.; Bolzoni, M.; Mancini, C.; Zaccarello, G.; Roato, I.; Morandi, F.; Marimpietri, D.; Faccani, G.; et al. Unraveling the Contribution of Ectoenzymes to Myeloma Life and Survival in the Bone Marrow Niche. Ann. N. Y. Acad. Sci. 2015, 1335, 10–22. [Google Scholar] [CrossRef] [PubMed]




© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Knowles, H.J. The Adenosine A2B Receptor Drives Osteoclast-Mediated Bone Resorption in Hypoxic Microenvironments. Cells 2019, 8, 624. https://doi.org/10.3390/cells8060624
Knowles HJ. The Adenosine A2B Receptor Drives Osteoclast-Mediated Bone Resorption in Hypoxic Microenvironments. Cells. 2019; 8(6):624. https://doi.org/10.3390/cells8060624
Chicago/Turabian StyleKnowles, Helen J. 2019. "The Adenosine A2B Receptor Drives Osteoclast-Mediated Bone Resorption in Hypoxic Microenvironments" Cells 8, no. 6: 624. https://doi.org/10.3390/cells8060624
APA StyleKnowles, H. J. (2019). The Adenosine A2B Receptor Drives Osteoclast-Mediated Bone Resorption in Hypoxic Microenvironments. Cells, 8(6), 624. https://doi.org/10.3390/cells8060624