Anti-Inflammatory Performance of Lactose-Modified Chitosan and Hyaluronic Acid Mixtures in an In Vitro Macrophage-Mediated Inflammation Osteoarthritis Model


 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Hyaluronic Acid (HA) and Lactose Modified Chitosan (CTL) Standard Solutions
2.2. Cell and Tissue Culture
2.2.1. Human Chondrocytes
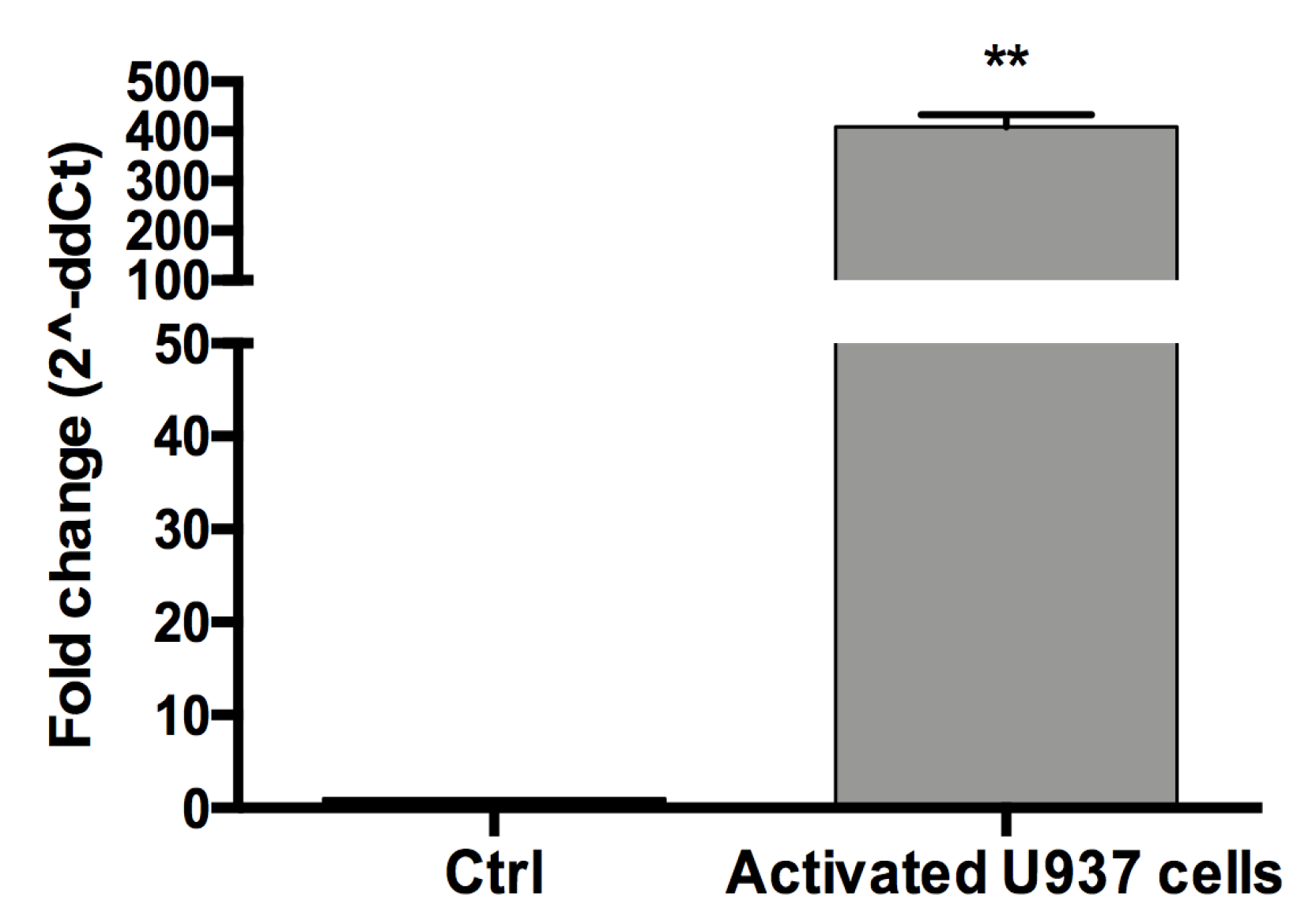
2.2.2. U937 Monocytes and Their Activation to Macrophages
2.3. Evaluation of the Effects of HA and CTL on Human Chondrocyte Viability
2.4. Analysis of Anti-Inflammatory and Anti-Oxidative Effects Induced by HA and CTL
2.4.1. RNA Isolation and Quantitative qPCR Analysis
2.4.2. Western Blotting Analysis
2.4.3. Detection of Intracellular ROS Generation
2.5. Statistical Analysis
3. Results
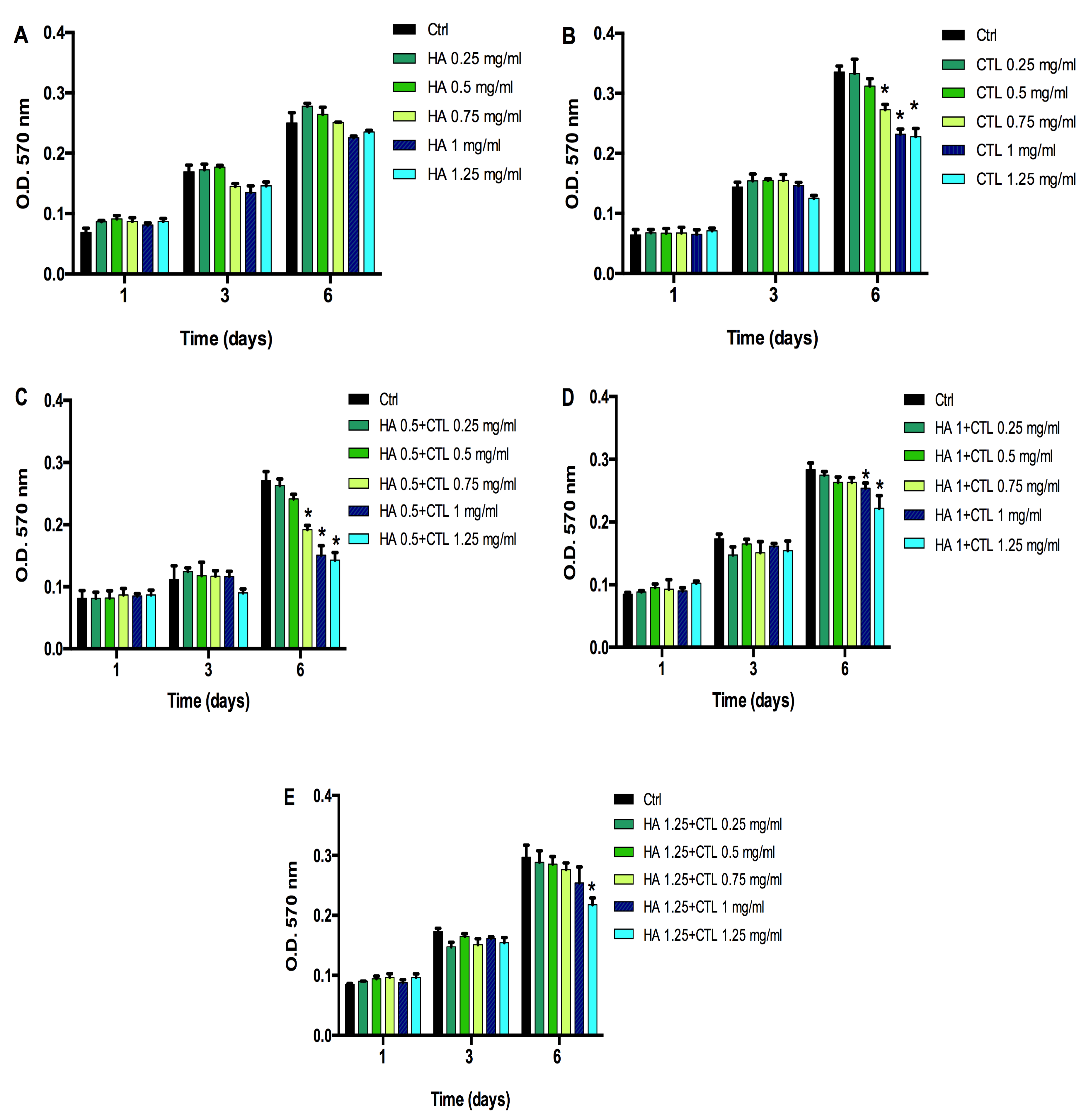
3.1. CTL Induces Changes in Chondrocytes Viability That Are Reduced by the Presence of HA
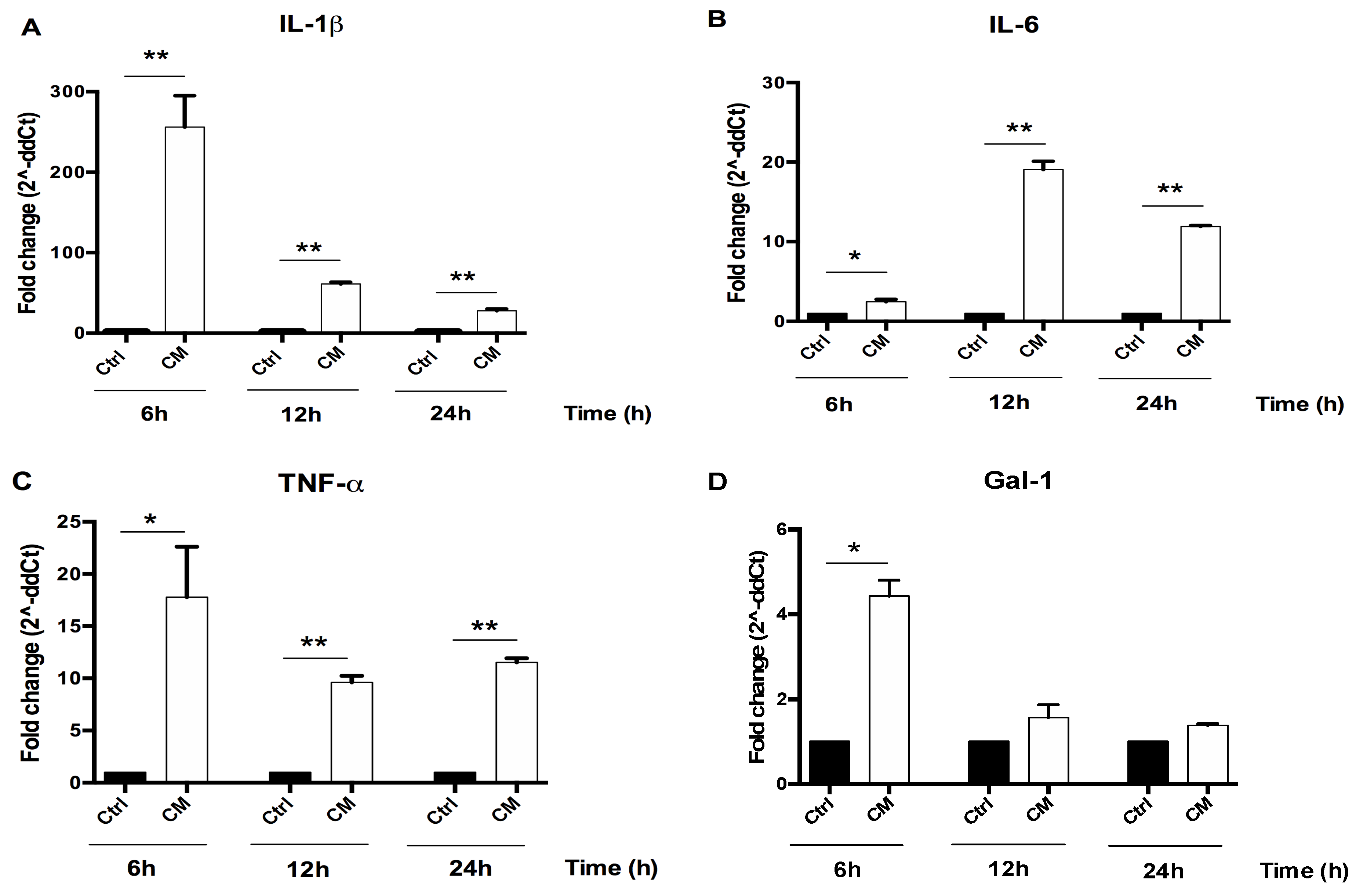
3.2. The Conditioned Medium (CM) of Activated U937 Cells Regulated Gene Expression of Human Chondrocytes
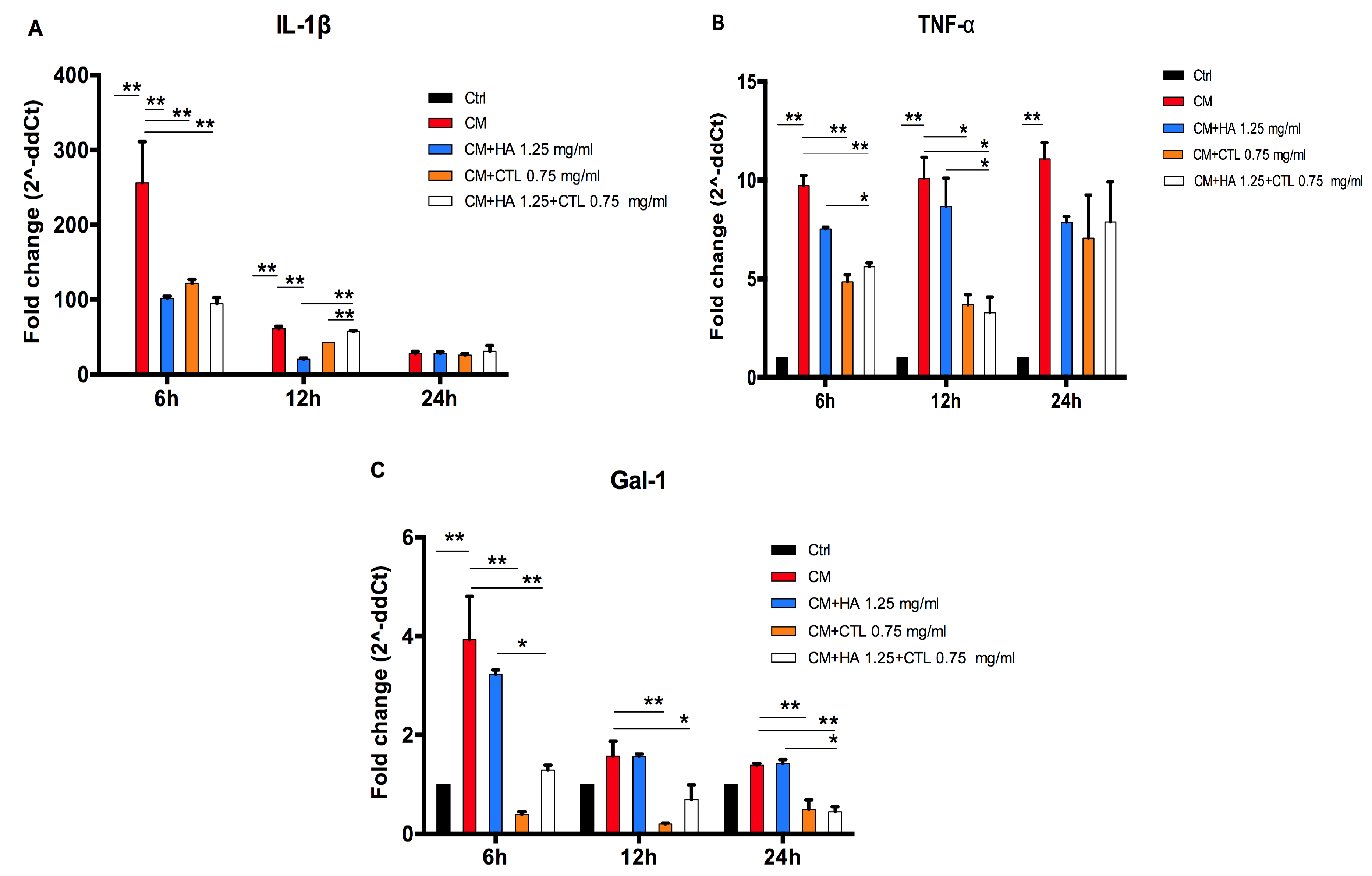
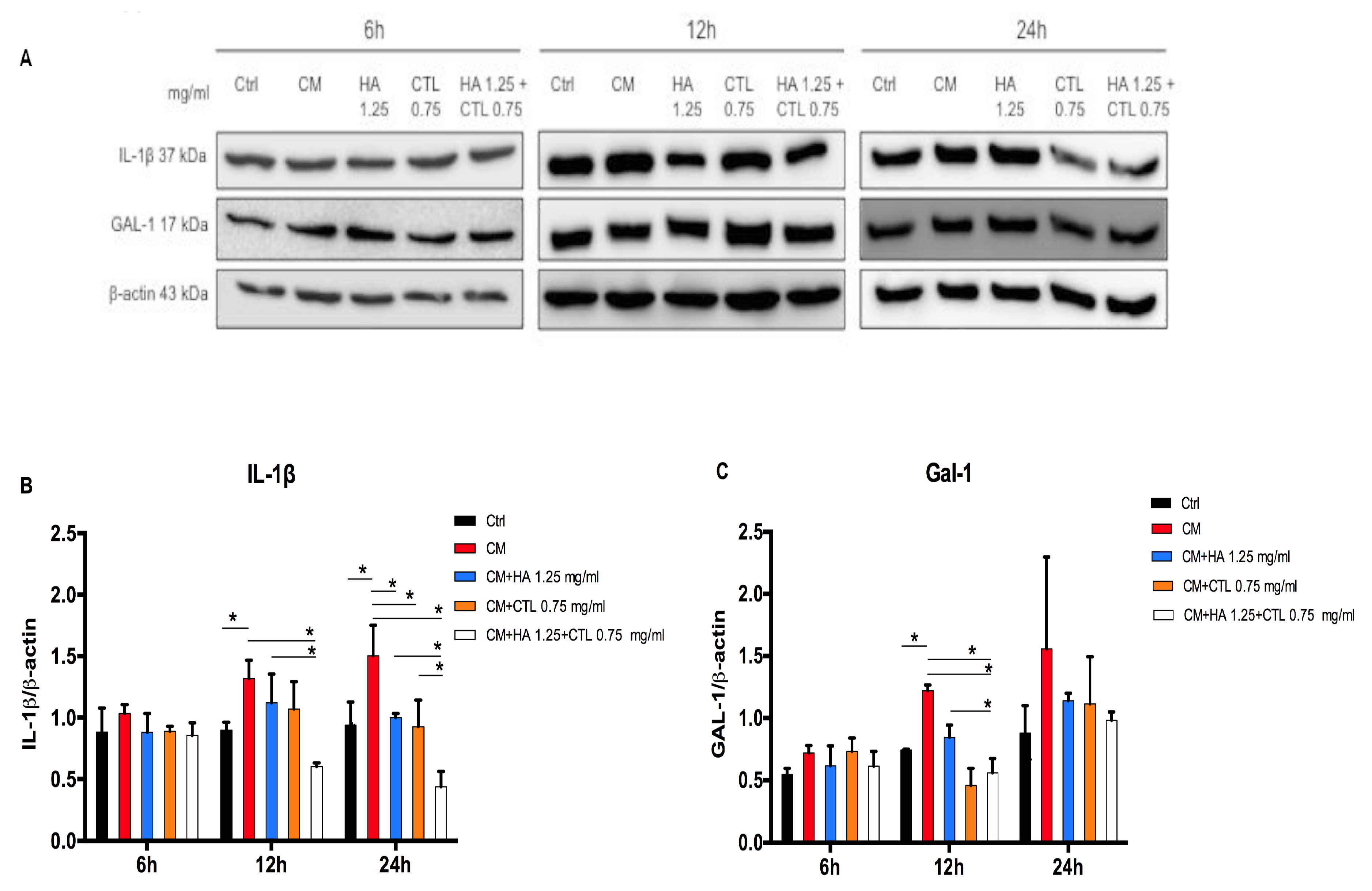
3.3. HA, CTL and HA-CTL Mixtures Reduce Chondrocyte Expression of Pro-Inflammatory Molecules
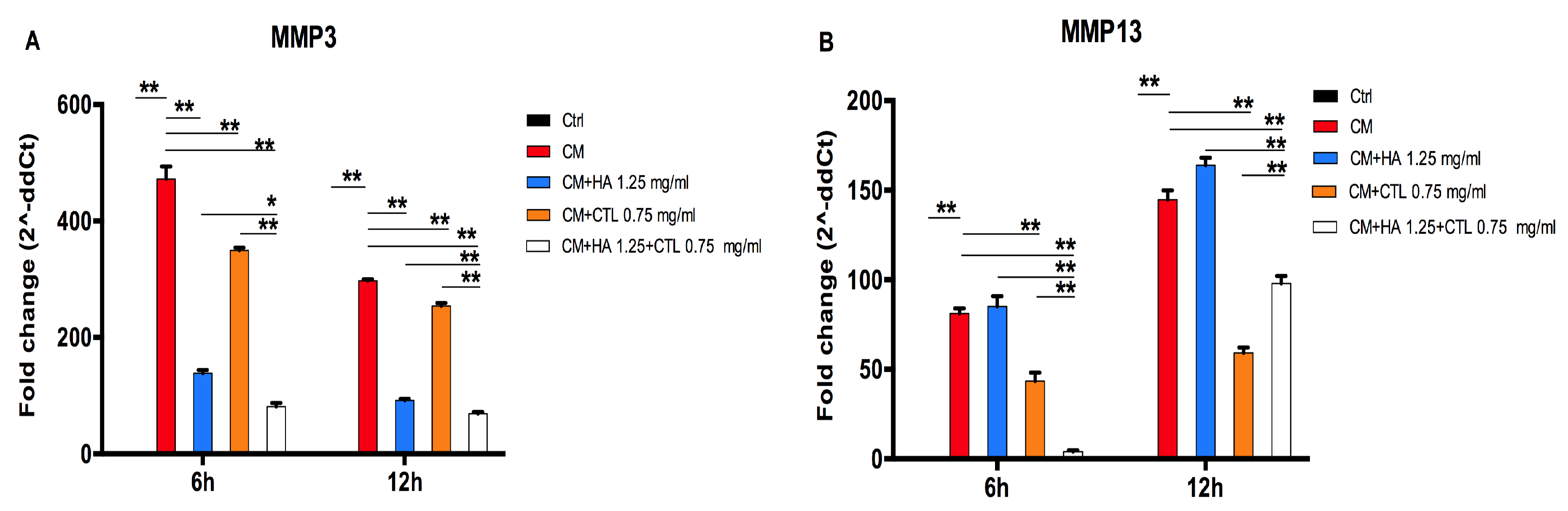
3.4. HA-CTL Mixture Down Regulated the Expression of MMPs
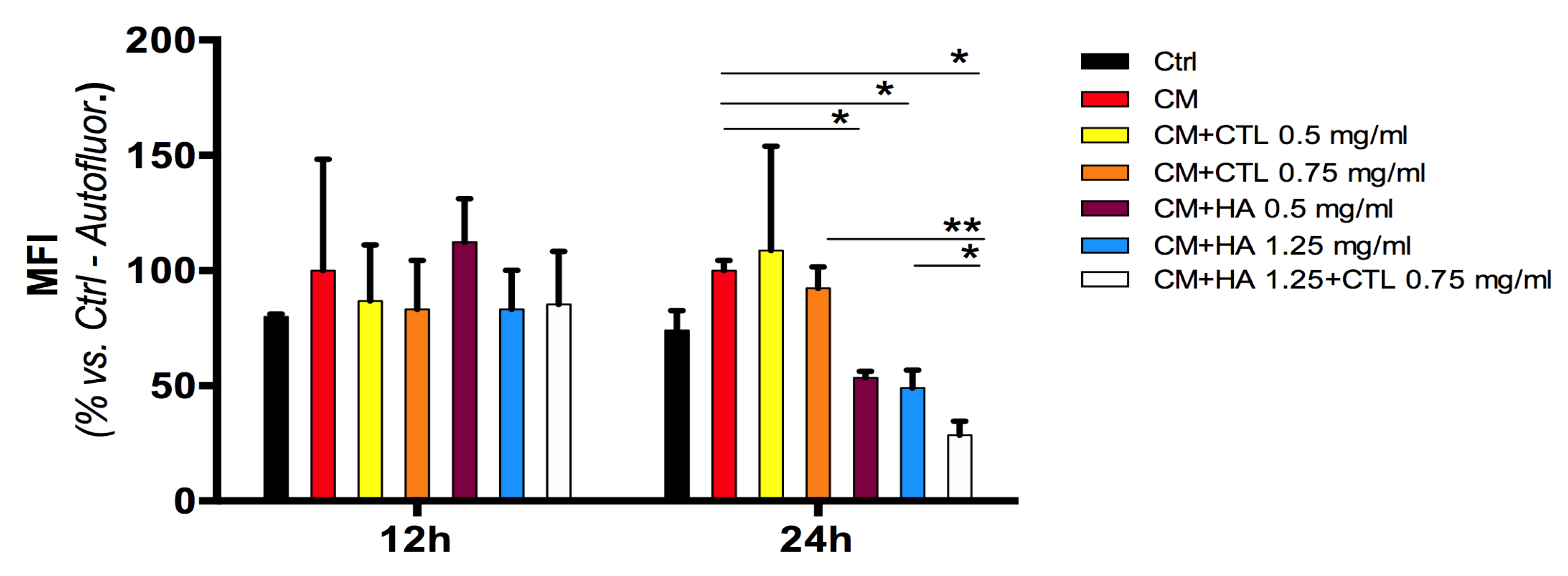
3.5. HA-CTL Mixture Induced a Reduction of ROS Formation in Human Chondrocytes Exposed to CM of U937 Activated Aonocytes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Loeser, R.F.; Goldring, S.R.; Scanzello, C.R.; Goldring, M.B. Osteoarthritis: A disease of the joint as an organ. Arthritis Rheum. 2012, 64, 1697–1707. [Google Scholar] [CrossRef] [Green Version]
- Glyn-Jones, S.; Palmer, A.J.; Agricola, R.; Price, A.J.; Vincent, T.L.; Weinans, H.; Carr, A.J. Osteoarthritis. Lancet 2015, 386, 376–387. [Google Scholar] [CrossRef]
- Maldonado, M.; Nam, J. The role of changes in extracellular matrix of cartilage in the presence of inflammation on the pathology of osteoarthritis. Biomed. Res. Int. 2013, 2013, 284873. [Google Scholar] [CrossRef] [Green Version]
- Kraus, V.B.; McDaniel, G.; Huebner, J.L.; Stabler, T.V.; Pieper, C.F.; Shipes, S.W.; Petry, N.A.; Low, P.S.; Shen, J.; McNearney, T.A.; et al. Direct in vivo evidence of activated macrophages in human osteoarthritis. Osteoarthr. Cartil. 2016, 24, 1613–1621. [Google Scholar] [CrossRef] [Green Version]
- Cuellar, V.G.; Cuellar, J.M.; Golish, S.R.; Yeomans, D.C.; Scuderi, G.J. Cytokine profiling in acute anterior cruciate ligament injury. Arthrosc. J. Arthrosc. Relat. Surg. 2010, 26, 1296–1301. [Google Scholar] [CrossRef]
- Toegel, S.; Bieder, D.; Andre, S.; Kayser, K.; Walzer, S.M.; Hobusch, G.; Windhager, R.; Gabius, H.J. Human osteoarthritic knee cartilage: Fingerprinting of adhesion/growth-regulatory galectins in vitro and in situ indicates differential upregulation in severe degeneration. Histochem. Cell Biol. 2014, 142, 373–388. [Google Scholar] [CrossRef] [PubMed]
- Osorio, J. Osteoarthritis: Galectin-1 damages cartilage via inflammation. Nat. Rev. Rheumatol. 2016, 12, 132. [Google Scholar] [CrossRef] [PubMed]
- Weinmann, D.; Kenn, M.; Schmidt, S.; Schmidt, K.; Walzer, S.M.; Kubista, B.; Windhager, R.; Schreiner, W.; Toegel, S.; Gabius, H.J. Galectin-8 induces functional disease markers in human osteoarthritis and cooperates with galectins-1 and -3. Cell. Mol. Life Sci. 2018, 75, 4187–4205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greenberg, D.D.; Stoker, A.; Kane, S.; Cockrell, M.; Cook, J.L. Biochemical effects of two different hyaluronic acid products in a co-culture model of osteoarthritis. Osteoarthr. Cartil. 2006, 14, 814–822. [Google Scholar] [CrossRef] [Green Version]
- Grishko, V.; Xu, M.; Ho, R.; Mates, A.; Watson, S.; Kim, J.T.; Wilson, G.L.; Pearsall, A.W.T. Effects of hyaluronic acid on mitochondrial function and mitochondria-driven apoptosis following oxidative stress in human chondrocytes. J. Biol. Chem. 2009, 284, 9132–9139. [Google Scholar] [CrossRef] [Green Version]
- Brun, P.; Panfilo, S.; Daga Gordini, D.; Cortivo, R.; Abatangelo, G. The effect of hyaluronan on CD44-mediated survival of normal and hydroxyl radical-damaged chondrocytes. Osteoarthr. Cartil. 2003, 11, 208–216. [Google Scholar] [CrossRef] [Green Version]
- Campo, G.M.; Avenoso, A.; Campo, S.; D’Ascola, A.; Traina, P.; Calatroni, A. Differential effect of molecular size HA in mouse chondrocytes stimulated with PMA. Biochim. Biophys. Acta 2009, 1790, 1353–1367. [Google Scholar] [CrossRef]
- Vitanzo, P.C., Jr.; Sennett, B.J. Hyaluronans: Is clinical effectiveness dependent on molecular weight? Am. J. Orthop. (Belle Meadn. J.) 2006, 35, 421–428. [Google Scholar]
- Abate, M.; Salini, V. Efficacy and safety study on a new compound associating low and high molecular weight hyaluronic acid in the treatment of hip osteoarthritis. Int. J. Immunopathol. Pharmacol. 2017, 30, 89–93. [Google Scholar] [CrossRef]
- Brun, P.; Zavan, B.; Vindigni, V.; Schiavinato, A.; Pozzuoli, A.; Iacobellis, C.; Abatangelo, G. In vitro response of osteoarthritic chondrocytes and fibroblast-like synoviocytes to a 500-730 kDa hyaluronan amide derivative. J. Biomed. Mater. Res. Part B Appl. Biomater. 2012, 100, 2073–2081. [Google Scholar] [CrossRef]
- Comblain, F.; Rocasalbas, G.; Gauthier, S.; Henrotin, Y. Chitosan: A promising polymer for cartilage repair and viscosupplementation. Bio-Med. Mater. Eng. 2017, 28, S209–S215. [Google Scholar] [CrossRef]
- Kaderli, S.; Boulocher, C.; Pillet, E.; Watrelot-Virieux, D.; Rougemont, A.L.; Roger, T.; Viguier, E.; Gurny, R.; Scapozza, L.; Jordan, O. A novel biocompatible hyaluronic acid-chitosan hybrid hydrogel for osteoarthrosis therapy. Int. J. Pharm. 2015, 483, 158–168. [Google Scholar] [CrossRef]
- Liu, S.-Q.; Qiu, B.; Chen, L.-y.; Peng, H.; Du, Y.-M. The effects of carboxymethylated chitosan on metalloproteinase-1, −3 and tissue inhibitor of metalloproteinase-1 gene expression in cartilage of experimental osteoarthritis. Rheumatol. Int. 2005, 26, 52–57. [Google Scholar] [CrossRef]
- Rieger, R.; Boulocher, C.; Kaderli, S.; Hoc, T. Chitosan in viscosupplementation: In vivo effect on rabbit subchondral bone. BMC Musculoskelet. Disord. 2017, 18, 350. [Google Scholar] [CrossRef] [Green Version]
- Patchornik, S.; Ram, E.; Ben Shalom, N.; Nevo, Z.; Robinson, D. Chitosan-hyaluronate hybrid gel intraarticular injection delays osteoarthritis progression and reduces pain in a rat meniscectomy model as compared to saline and hyaluronate treatment. Adv. Orthop. 2012, 2012, 979152. [Google Scholar] [CrossRef] [Green Version]
- Oprenyeszk, F.; Chausson, M.; Maquet, V.; Dubuc, J.E.; Henrotin, Y. Protective effect of a new biomaterial against the development of experimental osteoarthritis lesions in rabbit: A pilot study evaluating the intra-articular injection of alginate-chitosan beads dispersed in an hydrogel. Osteoarthr. Cartil. 2013, 21, 1099–1107. [Google Scholar] [CrossRef] [Green Version]
- Donati, I.; Stredanska, S.; Silvestrini, G.; Vetere, A.; Marcon, P.; Marsich, E.; Mozetic, P.; Gamini, A.; Paoletti, S.; Vittur, F. The aggregation of pig articular chondrocyte and synthesis of extracellular matrix by a lactose-modified chitosan. Biomaterials 2005, 26, 987–998. [Google Scholar] [CrossRef]
- Marcon, P.; Marsich, E.; Vetere, A.; Mozetic, P.; Campa, C.; Donati, I.; Vittur, F.; Gamini, A.; Paoletti, S. The role of Galectin-1 in the interaction between chondrocytes and a lactose-modified chitosan. Biomaterials 2005, 26, 4975–4984. [Google Scholar] [CrossRef] [PubMed]
- Salamanna, F.; Veronesi, F.; Frizziero, A.; Fini, M. Role and translational implication of galectins in arthritis pathophysiology and treatment: A systematic literature review. J. Cell. Physiol. 2019, 234, 1588–1605. [Google Scholar] [CrossRef] [PubMed]
- Donati, I.; Borgogna, M.; Turello, E.; Cesàro, A.; Paoletti, S. Tuning Supramolecular Structuring at the Nanoscale Level: Nonstoichiometric Soluble Complexes in Dilute Mixed Solutions of Alginate and Lactose-Modified Chitosan (Chitlac). Biomacromolecules 2007, 8, 1471–1479. [Google Scholar] [CrossRef] [PubMed]
- Salamanna, F.; Giavaresi, G.; Parrilli, A.; Martini, L.; Nicoli Aldini, N.; Abatangelo, G.; Frizziero, A.; Fini, M. Effects of intra-articular hyaluronic acid associated to Chitlac (arty-duo(R)) in a rat knee osteoarthritis model. J. Orthop. Res. 2019, 37, 867–876. [Google Scholar] [CrossRef]
- Brun, P.; Abatangelo, G.; Radice, M.; Zacchi, V.; Guidolin, D.; Daga Gordini, D.; Cortivo, R. Chondrocyte aggregation and reorganization into three-dimensional scaffolds. J. Biomed. Mater. Res. 1999, 46, 337–346. [Google Scholar] [CrossRef]
- Denizot, F.; Lang, R. Rapid colorimetric assay for cell growth and survival. Modifications to the tetrazolium dye procedure giving improved sensitivity and reliability. J. Immunol. Methods 1986, 89, 271–277. [Google Scholar] [CrossRef]
- Marsich, E.; Borgogna, M.; Donati, I.; Mozetic, P.; Strand, B.L.; Salvador, S.G.; Vittur, F.; Paoletti, S. Alginate/lactose-modified chitosan hydrogels: A bioactive biomaterial for chondrocyte encapsulation. J. Biomed. Mater. Res. Part A 2008, 84, 364–376. [Google Scholar] [CrossRef]
- Marchev, A.S.; Dimitrova, P.A.; Burns, A.J.; Kostov, R.V.; Dinkova-Kostova, A.T.; Georgiev, M.I. Oxidative stress and chronic inflammation in osteoarthritis: Can NRF2 counteract these partners in crime? Ann. N. Y. Acad. Sci. 2017, 1401, 114–135. [Google Scholar] [CrossRef]
- Onodera, Y.; Teramura, T.; Takehara, T.; Fukuda, K. Hyaluronic acid regulates a key redox control factor Nrf2 via phosphorylation of Akt in bovine articular chondrocytes. FEBS Open Bio 2015, 5, 476–484. [Google Scholar] [CrossRef] [Green Version]
- Avenoso, A.; D’Ascola, A.; Scuruchi, M.; Mandraffino, G.; Calatroni, A.; Saitta, A.; Campo, S.; Campo, G.M. Hyaluronan in the experimental injury of the cartilage: Biochemical action and protective effects. Inflamm. Res. 2018, 67, 5–20. [Google Scholar] [CrossRef]
- Knudson, C.B.; Knudson, W. Hyaluronan and CD44: Modulators of chondrocyte metabolism. Clin. Orthop. Relat. Res. 2004, 427, S152–S162. [Google Scholar] [CrossRef]
- Goldring, M.B.; Goldring, S.R. Osteoarthritis. J. Cell. Physiol. 2007, 213, 626–634. [Google Scholar] [CrossRef]
- Haraden, C.A.; Huebner, J.L.; Hsueh, M.F.; Li, Y.J.; Kraus, V.B. Synovial fluid biomarkers associated with osteoarthritis severity reflect macrophage and neutrophil related inflammation. Arthritis Res. Ther. 2019, 21, 146. [Google Scholar] [CrossRef] [Green Version]
- Santangelo, K.S.; Nuovo, G.J.; Bertone, A.L. In vivo reduction or blockade of interleukin-1β in primary osteoarthritis influences expression of mediators implicated in pathogenesis. Osteoarthr. Cartil. 2012, 20, 1610–1618. [Google Scholar] [CrossRef] [Green Version]
- Sachs, D.; Cunha, F.Q.; Poole, S.; Ferreira, S.H. Tumour necrosis factor-alpha, interleukin-1beta and interleukin-8 induce persistent mechanical nociceptor hypersensitivity. Pain 2002, 96, 89–97. [Google Scholar] [CrossRef]
- Toegel, S.; Weinmann, D.; Andre, S.; Walzer, S.M.; Bilban, M.; Schmidt, S.; Chiari, C.; Windhager, R.; Krall, C.; Bennani-Baiti, I.M.; et al. Galectin-1 Couples Glycobiology to Inflammation in Osteoarthritis through the Activation of an NF-kappaB-Regulated Gene Network. J. Immunol. 2016, 196, 1910–1921. [Google Scholar] [CrossRef] [Green Version]
- Axe, J.M.; Snyder-Mackler, L.; Axe, M.J. The role of viscosupplementation. Sports Med. Arthrosc. Rev. 2013, 21, 18–22. [Google Scholar] [CrossRef]
- Zhang, F.J.; Gao, S.G.; Cheng, L.; Tian, J.; Xu, W.S.; Luo, W.; Song, Y.; Yang, Y.; Lei, G.H. The effect of hyaluronic acid on osteopontin and CD44 mRNA of fibroblast-like synoviocytes in patients with osteoarthritis of the knee. Rheumatol. Int. 2013, 33, 79–83. [Google Scholar] [CrossRef]
- Funato, S.; Yasuhara, R.; Yoshimura, K.; Miyamoto, Y.; Kaneko, K.; Suzawa, T.; Chikazu, D.; Mishima, K.; Baba, K.; Kamijo, R. Extracellular matrix loss in chondrocytes after exposure to interleukin-1beta in NADPH oxidase-dependent manner. Cell Tissue Res. 2017, 368, 135–144. [Google Scholar] [CrossRef] [PubMed]
- McDermott, B.T.; Peffers, M.J.; McDonagh, B.; Tew, S.R. Translational regulation contributes to the secretory response of chondrocytic cells following exposure to interleukin-1beta. J. Biol. Chem. 2019, 294, 13027–13039. [Google Scholar] [CrossRef] [Green Version]
- Weinmann, D.; Schlangen, K.; Andre, S.; Schmidt, S.; Walzer, S.M.; Kubista, B.; Windhager, R.; Toegel, S.; Gabius, H.J. Galectin-3 Induces a Pro-degradative/inflammatory Gene Signature in Human Chondrocytes, Teaming Up with Galectin-1 in Osteoarthritis Pathogenesis. Sci. Rep. 2016, 6, 39112. [Google Scholar] [CrossRef]
- Fernandes, J.C.; Martel-Pelletier, J.; Pelletier, J.P. The role of cytokines in osteoarthritis pathophysiology. Biorheology 2002, 39, 237–246. [Google Scholar]
- Li, H.; Wang, D.; Yuan, Y.; Min, J. New insights on the MMP-13 regulatory network in the pathogenesis of early osteoarthritis. Arthritis Res. Ther. 2017, 19, 248. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Sampson, E.R.; Jin, H.; Li, J.; Ke, Q.H.; Im, H.J.; Chen, D. MMP13 is a critical target gene during the progression of osteoarthritis. Arthritis Res. Ther. 2013, 15, R5. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene (Accession Number) | Name | Primer Sequences |
|---|---|---|
| PPIA (NM_021130.5) | Peptidylprolyl Isomerase A | Fw 5′-GGGCTTTAGGCTGTAGGTCAA-3′ Rv 5′-AACCAAAGCTAGGGAGAGGC-3′ |
| IL-1β (NM_000576.3) | Interleukin 1 beta | Fw 5′-GAATCTCCGACCACCACTACAG-3′ Rv 5′-TGATCGTACAGGTGCATCGTG-3′ |
| TNF-α (NM_000594.4) | Tumour necrosis factor alpha | Fw 5′-AAGCCTGTAGCCCATGTTGT-3′ Rv 5′-GGACCTGGGAGTAGATGAGGT-3′ |
| GAL-1 (NM_002305.4) | Galectin 1 | Fw 5′-TCTCGGGTGGAGTCTTCTGA-3′ Rv 5′-GTTCAGCACGAAGCTCTTAGC-3′ |
| MMP-3 (J03209.1) | Matrix metalloproteinase 3 | Fw 5′-TCACTCACAGACCTGACTCG-3′ Rv 5′-AAAGCAGGATCACAGTTGGC-3′ |
| MMP-13 (AY741163.1) | Matrix metalloproteinase 13 | Fw 5′-AACGCCAGACAAATGTGACC-3′ Rv 5′-AGGTCATGAGAAGGGTGCTC-3′ |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tarricone, E.; Mattiuzzo, E.; Belluzzi, E.; Elia, R.; Benetti, A.; Venerando, R.; Vindigni, V.; Ruggieri, P.; Brun, P. Anti-Inflammatory Performance of Lactose-Modified Chitosan and Hyaluronic Acid Mixtures in an In Vitro Macrophage-Mediated Inflammation Osteoarthritis Model. Cells 2020, 9, 1328. https://doi.org/10.3390/cells9061328
Tarricone E, Mattiuzzo E, Belluzzi E, Elia R, Benetti A, Venerando R, Vindigni V, Ruggieri P, Brun P. Anti-Inflammatory Performance of Lactose-Modified Chitosan and Hyaluronic Acid Mixtures in an In Vitro Macrophage-Mediated Inflammation Osteoarthritis Model. Cells. 2020; 9(6):1328. https://doi.org/10.3390/cells9061328
Chicago/Turabian StyleTarricone, Elena, Elena Mattiuzzo, Elisa Belluzzi, Rossella Elia, Andrea Benetti, Rina Venerando, Vincenzo Vindigni, Pietro Ruggieri, and Paola Brun. 2020. "Anti-Inflammatory Performance of Lactose-Modified Chitosan and Hyaluronic Acid Mixtures in an In Vitro Macrophage-Mediated Inflammation Osteoarthritis Model" Cells 9, no. 6: 1328. https://doi.org/10.3390/cells9061328
APA StyleTarricone, E., Mattiuzzo, E., Belluzzi, E., Elia, R., Benetti, A., Venerando, R., Vindigni, V., Ruggieri, P., & Brun, P. (2020). Anti-Inflammatory Performance of Lactose-Modified Chitosan and Hyaluronic Acid Mixtures in an In Vitro Macrophage-Mediated Inflammation Osteoarthritis Model. Cells, 9(6), 1328. https://doi.org/10.3390/cells9061328