Hepatitis B Virus Entry into Cells


Abstract
:1. An Introduction to Virus Entry
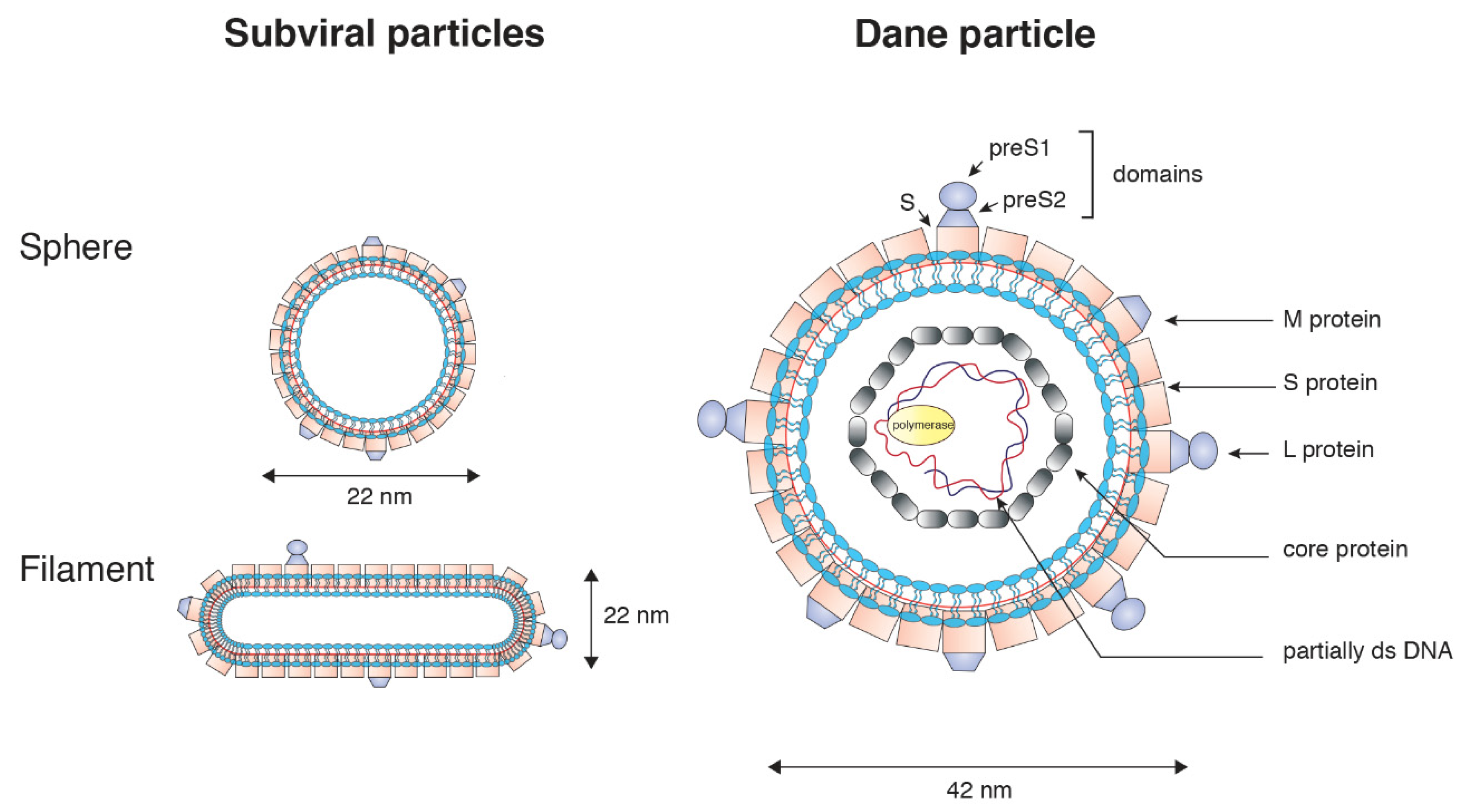
2. General Features of the Hepatitis B Virus
3. Mechanisms of HBV Entry into Cells
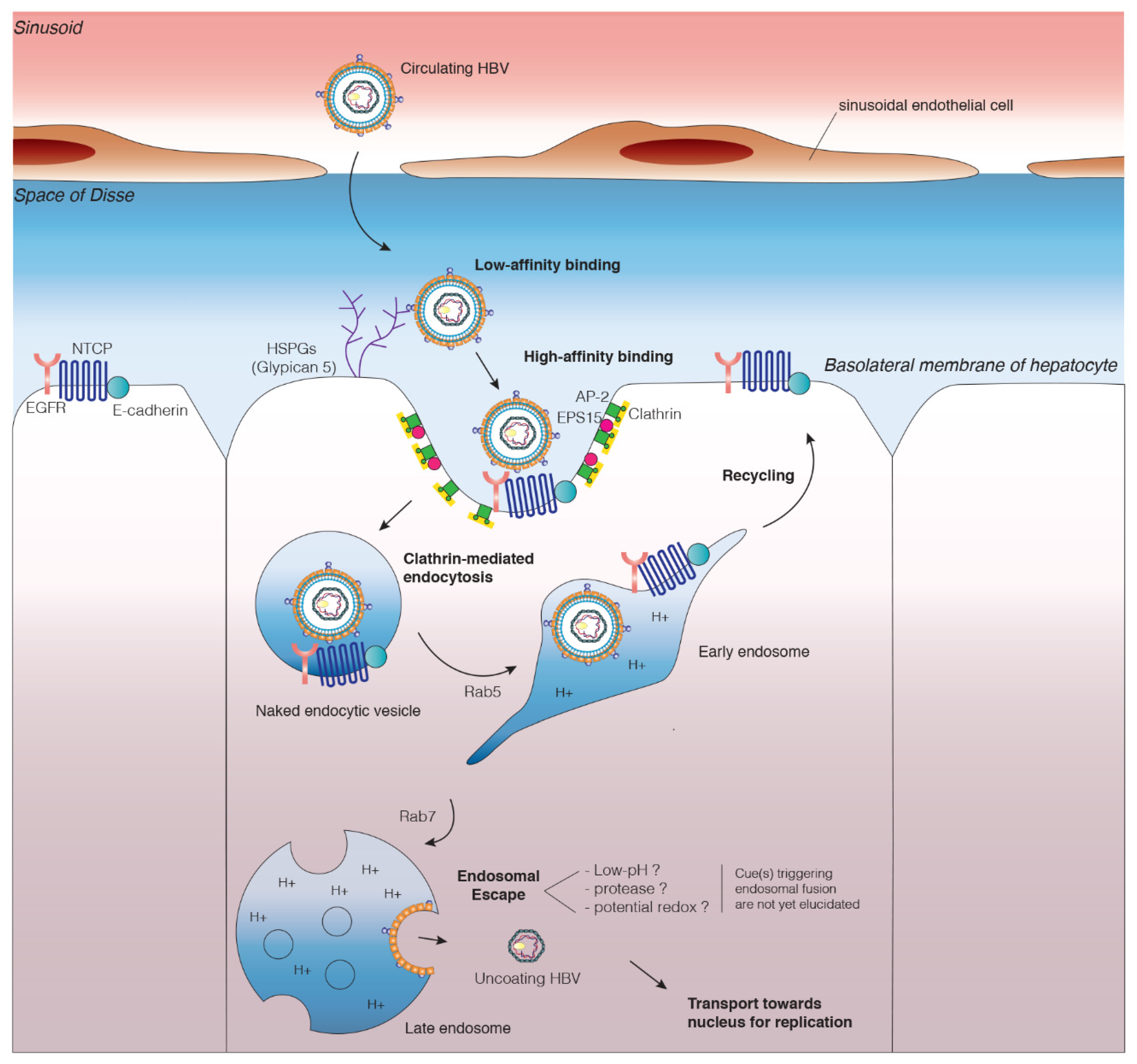
3.1. Attachment of HBV Particles to the Cell Surface
3.1.1. Low-Affinity Binding to Heparin Sulfate Proteoglycans
3.1.2. High-Affinity Binding to the NTCP Receptor
3.2. Viral Entry
3.2.1. Host Cell Factors Involved in HBV Internalization
3.2.2. Endocytosis Mediated by Receptor Binding
3.3. Viral Escape from Endosomes
4. Inhibition of Entry as a Pipeline for HBV Treatment
5. Conclusion and Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Doms, R.W.; Moore, J.P. HIV-1 Membrane Fusion. J. Cell Biol. 2000, 151, F9–F14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wittels, M.; Spear, P.G. Penetration of cells by herpes simplex virus does not require a low pH-dependent endocytic pathway. Virus Res. 1991, 18, 271–290. [Google Scholar] [CrossRef]
- Singh, M.; Jadhav, H.R.; Bhatt, T. Dynamin Functions and Ligands: Classical Mechanisms Behind. Mol. Pharmacol. 2017, 91, 123–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henley, J.R.; Krueger, E.W.A.; Oswald, B.J.; McNiven, M.A. Dynamin-mediated Internalization of Caveolae. J. Cell Biol. 1998, 141, 85–99. [Google Scholar] [CrossRef] [Green Version]
- McMahon, H.T.; Boucrot, E. Molecular mechanism and physiological functions of clathrin-mediated endocytosis. Nat. Rev. Mol. Cell Biol. 2011, 12, 517–533. [Google Scholar] [CrossRef]
- Nabi, I.R.; Le, P.U. Caveolae/raft-dependent endocytosis. J. Cell Biol. 2003, 161, 673–677. [Google Scholar] [CrossRef] [PubMed]
- Mayor, S.; Pagano, R.E. Pathways of clathrin-independent endocytosis. Nat. Rev. Mol. Cell Biol. 2007, 8, 603–612. [Google Scholar] [CrossRef]
- Matlin, K.S.; Reggio, H.; Helenius, A.; Simons, K. Infectious entry pathway of influenza virus in a canine kidney cell line. J. Cell Biol. 1981, 91, 601–613. [Google Scholar] [CrossRef] [Green Version]
- Rust, M.J.; Lakadamyali, M.; Zhang, F.; Zhuang, X. Assembly of endocytic machinery around individual influenza viruses during viral entry. Nat. Struct. Mol. Biol. 2004, 11, 567–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanchard, E.; Belouzard, S.; Goueslain, L.; Wakita, T.; Dubuisson, J.; Wychowski, C.; Rouillé, Y. Hepatitis C virus entry depends on clathrin-mediated endocytosis. J. Virol. 2006, 80, 6964–6972. [Google Scholar] [CrossRef] [Green Version]
- Meertens, L.; Bertaux, C.; Dragic, T. Hepatitis C Virus Entry Requires a Critical Postinternalization Step and Delivery to Early Endosomes via Clathrin-Coated Vesicles. J. Virol. 2006, 80, 11571–11578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acosta, E.G.; Castilla, V.; Damonte, E.B. Functional entry of dengue virus into Aedes albopictus mosquito cells is dependent on clathrin-mediated endocytosis. J. Gen. Virol. 2008, 89, 474–484. [Google Scholar] [CrossRef] [PubMed]
- Piccini, L.E.; Castilla, V.; Damonte, E.B. Dengue-3 Virus Entry into Vero Cells: Role of Clathrin-Mediated Endocytosis in the Outcome of Infection. PLoS ONE 2015, 10, e0140824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matlin, K.S.; Reggio, H.; Helenius, A.; Simons, K. Pathway of vesicular stomatitis virus entry leading to infection. J. Mol. Biol. 1982, 156, 609–631. [Google Scholar] [CrossRef]
- Johannsdottir, H.K.; Mancini, R.; Kartenbeck, J.; Amato, L.; Helenius, A. Host Cell Factors and Functions Involved in Vesicular Stomatitis Virus Entry. J. Virol. 2009, 83, 440–453. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.-C.; Chen, C.-C.; Chang, W.-C.; Tao, M.-H.; Huang, C. Entry of hepatitis B virus into immortalized human primary hepatocytes by clathrin-dependent endocytosis. J. Virol. 2012, 86, 9443–9453. [Google Scholar] [CrossRef] [Green Version]
- Umetsu, T.; Inoue, J.; Kogure, T.; Kakazu, E.; Ninomiya, M.; Iwata, T.; Takai, S.; Nakamura, T.; Sano, A.; Shimosegawa, T. Inhibitory effect of silibinin on hepatitis B virus entry. Biochem. Biophys. Rep. 2018, 14, 20–25. [Google Scholar] [CrossRef]
- Herrscher, C.; Pastor, F.; Burlaud-Gaillard, J.; Dumans, A.; Seigneuret, F.; Moreau, A.; Patient, R.; Eymieux, S.; de Rocquigny, H.; Hourioux, C.; et al. Hepatitis B virus entry into HepG2-NTCP cells requires clathrin-mediated endocytosis. Cell. Microbiol. 2020, e13205. [Google Scholar] [CrossRef]
- Anderson, H.A.; Chen, Y.; Norkin, L.C. Bound simian virus 40 translocates to caveolin-enriched membrane domains, and its entry is inhibited by drugs that selectively disrupt caveolae. Mol. Biol. Cell 1996, 7, 1825–1834. [Google Scholar] [CrossRef] [Green Version]
- Pelkmans, L.; Kartenbeck, J.; Helenius, A. Caveolar endocytosis of simian virus 40 reveals a new two-step vesicular-transport pathway to the ER. Nat. Cell Biol. 2001, 3, 473–483. [Google Scholar] [CrossRef]
- Macovei, A.; Radulescu, C.; Lazar, C.; Petrescu, S.; Durantel, D.; Dwek, R.A.; Zitzmann, N.; Nichita, N.B. Hepatitis B virus requires intact caveolin-1 function for productive infection in HepaRG cells. J. Virol. 2010, 84, 243–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saeed, M.F.; Kolokoltsov, A.A.; Albrecht, T.; Davey, R.A. Cellular entry of ebola virus involves uptake by a macropinocytosis-like mechanism and subsequent trafficking through early and late endosomes. PLoS Pathog. 2010, 6, e1001110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kondratowicz, A.S.; Hunt, C.L.; Davey, R.A.; Cherry, S.; Maury, W.J. AMP-activated protein kinase is required for the macropinocytic internalization of ebolavirus. J. Virol. 2013, 87, 746–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mercer, J.; Helenius, A. Vaccinia Virus Uses Macropinocytosis and Apoptotic Mimicry to Enter Host Cells. Science 2008, 320, 531–535. [Google Scholar] [CrossRef]
- Huang, C.-Y.; Lu, T.-Y.; Bair, C.-H.; Chang, Y.-S.; Jwo, J.-K.; Chang, W. A Novel Cellular Protein, VPEF, Facilitates Vaccinia Virus Penetration into HeLa Cells through Fluid Phase Endocytosis. J. Virol. 2008, 82, 7988–7999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amstutz, B.; Gastaldelli, M.; Kälin, S.; Imelli, N.; Boucke, K.; Wandeler, E.; Mercer, J.; Hemmi, S.; Greber, U.F. Subversion of CtBP1-controlled macropinocytosis by human adenovirus serotype 3. EMBO J. 2008, 27, 956–969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez-San Martin, C.; Lopez, T.; Arias, C.F.; Lopez, S. Characterization of Rotavirus Cell Entry. J. Virol. 2004, 78, 2310–2318. [Google Scholar] [CrossRef] [Green Version]
- Arias, C.F.; Silva-Ayala, D.; López, S. Rotavirus Entry: A Deep Journey into the Cell with Several Exits. J. Virol. 2015, 89, 890–893. [Google Scholar] [CrossRef] [Green Version]
- Nonnenmacher, M.; Weber, T. Adeno-Associated Virus 2 Infection Requires Endocytosis through the CLIC/GEEC Pathway. Cell Host Microbe 2011, 10, 563–576. [Google Scholar] [CrossRef] [Green Version]
- Heikkila, O.; Susi, P.; Tevaluoto, T.; Harma, H.; Marjomaki, V.; Hyypia, T.; Kiljunen, S. Internalization of Coxsackievirus A9 Is Mediated by 2-Microglobulin, Dynamin, and Arf6 but Not by Caveolin-1 or Clathrin. J. Virol. 2010, 84, 3666–3681. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.-L.; Liu, Y.-G.; Zhou, Y.-T.; Zhao, P.; Ren, H.; Xiao, M.; Zhu, Y.-Z.; Qi, Z.-T. Endophilin-A2-mediated endocytic pathway is critical for enterovirus 71 entry into caco-2 cells. Emerg. Microbes Infect. 2019, 8, 773–786. [Google Scholar] [CrossRef] [PubMed]
- White, J.M.; Whittaker, G.R. Fusion of Enveloped Viruses in Endosomes. Traffic Cph. Den. 2016, 17, 593–614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamontagne, R.J.; Bagga, S.; Bouchard, M.J. Hepatitis B virus molecular biology and pathogenesis. Hepatoma Res. 2016, 2, 163–186. [Google Scholar] [CrossRef] [PubMed]
- Glebe, D.; Urban, S. Viral and cellular determinants involved in hepadnaviral entry. World J. Gastroenterol. 2007, 13, 22–38. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Liu, K. Complete and Incomplete Hepatitis B Virus Particles: Formation, Function, and Application. Viruses 2017, 9, 56. [Google Scholar] [CrossRef] [Green Version]
- Patient, R.; Hourioux, C.; Roingeard, P. Morphogenesis of hepatitis B virus and its subviral envelope particles. Cell. Microbiol. 2009, 11, 1561–1570. [Google Scholar] [CrossRef] [Green Version]
- Gavilanes, F.; Gonzalez-Ros, J.M.; Peterson, D.L. Structure of hepatitis B surface antigen. Characterization of the lipid components and their association with the viral proteins. J. Biol. Chem. 1982, 257, 7770–7777. [Google Scholar]
- Bruss, V. Hepatitis B virus morphogenesis. World J. Gastroenterol. 2007, 13, 65. [Google Scholar] [CrossRef] [Green Version]
- Schweitzer, A.; Horn, J.; Mikolajczyk, R.T.; Krause, G.; Ott, J.J. Estimations of worldwide prevalence of chronic hepatitis B virus infection: A systematic review of data published between 1965 and 2013. Lancet Lond. Engl. 2015, 386, 1546–1555. [Google Scholar] [CrossRef]
- Gish, R.; Jia, J.-D.; Locarnini, S.; Zoulim, F. Selection of chronic hepatitis B therapy with high barrier to resistance. Lancet Infect. Dis. 2012, 12, 341–353. [Google Scholar] [CrossRef]
- Sells, M.A.; Chen, M.L.; Acs, G. Production of hepatitis B virus particles in Hep G2 cells transfected with cloned hepatitis B virus DNA. Proc. Natl. Acad. Sci. USA 1987, 84, 1005–1009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sureau, C.; Romet-Lemonne, J.L.; Mullins, J.I.; Essex, M. Production of hepatitis B virus by a differentiated human hepatoma cell line after transfection with cloned circular HBV DNA. Cell 1986, 47, 37–47. [Google Scholar] [CrossRef]
- Hu, J.; Lin, Y.-Y.; Chen, P.-J.; Watashi, K.; Wakita, T. Cell and Animal Models for Studying Hepatitis B Virus Infection and Drug Development. Gastroenterology 2019, 156, 338–354. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Zhong, G.; Xu, G.; He, W.; Jing, Z.; Gao, Z.; Huang, Y.; Qi, Y.; Peng, B.; Wang, H.; et al. Sodium taurocholate cotransporting polypeptide is a functional receptor for human hepatitis B and D virus. eLife 2012, 1, e00049. [Google Scholar] [CrossRef] [PubMed]
- Ni, Y.; Lempp, F.A.; Mehrle, S.; Nkongolo, S.; Kaufman, C.; Fälth, M.; Stindt, J.; Königer, C.; Nassal, M.; Kubitz, R.; et al. Hepatitis B and D viruses exploit sodium taurocholate co-transporting polypeptide for species-specific entry into hepatocytes. Gastroenterology 2014, 146, 1070–1083. [Google Scholar] [CrossRef]
- Sarrazin, S.; Lamanna, W.C.; Esko, J.D. Heparan Sulfate Proteoglycans. Cold Spring Harb. Perspect. Biol. 2011, 3, a004952. [Google Scholar] [CrossRef] [Green Version]
- WuDunn, D.; Spear, P.G. Initial interaction of herpes simplex virus with cells is binding to heparan sulfate. J. Virol. 1989, 63, 52–58. [Google Scholar] [CrossRef] [Green Version]
- Giroglou, T.; Florin, L.; Schäfer, F.; Streeck, R.E.; Sapp, M. Human papillomavirus infection requires cell surface heparan sulfate. J. Virol. 2001, 75, 1565–1570. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Maguire, T.; Hileman, R.E.; Fromm, J.R.; Esko, J.D.; Linhardt, R.J.; Marks, R.M. Dengue virus infectivity depends on envelope protein binding to target cell heparan sulfate. Nat. Med. 1997, 3, 866–871. [Google Scholar] [CrossRef]
- Leistner, C.M.; Gruen-Bernhard, S.; Glebe, D. Role of glycosaminoglycans for binding and infection of hepatitis B virus. Cell. Microbiol. 2008, 10, 122–133. [Google Scholar] [CrossRef]
- Schulze, A.; Gripon, P.; Urban, S. Hepatitis B virus infection initiates with a large surface protein-dependent binding to heparan sulfate proteoglycans. Hepatol. Baltim. Md. 2007, 46, 1759–1768. [Google Scholar] [CrossRef] [PubMed]
- Ying, C.; Van Pelt, J.F.; Van Lommel, A.; Van Ranst, M.; Leyssen, P.; De Clercq, E.; Neyts, J. Sulphated and sulphonated polymers inhibit the initial interaction of hepatitis B virus with hepatocytes. Antivir. Chem. Chemother. 2002, 13, 157–164. [Google Scholar] [CrossRef]
- Lindblom, A.; Fransson, L.A. Endothelial heparan sulphate: Compositional analysis and comparison of chains from different proteoglycan populations. Glycoconj. J. 1990, 7, 545–562. [Google Scholar] [CrossRef] [PubMed]
- Verrier, E.R.; Colpitts, C.C.; Bach, C.; Heydmann, L.; Weiss, A.; Renaud, M.; Durand, S.C.; Habersetzer, F.; Durantel, D.; Abou-Jaoudé, G.; et al. A targeted functional RNA interference screen uncovers glypican 5 as an entry factor for hepatitis B and D viruses. Hepatol. Baltim. Md. 2016, 63, 35–48. [Google Scholar] [CrossRef]
- Seitz, S.; Iancu, C.; Volz, T.; Mier, W.; Dandri, M.; Urban, S.; Bartenschlager, R. A Slow Maturation Process Renders Hepatitis B Virus Infectious. Cell Host Microbe 2016, 20, 25–35. [Google Scholar] [CrossRef] [Green Version]
- Sureau, C.; Salisse, J. A conformational heparan sulfate binding site essential to infectivity overlaps with the conserved hepatitis B virus a-determinant. Hepatol. Baltim. Md. 2013, 57, 985–994. [Google Scholar] [CrossRef]
- Blanchet, M.; Sureau, C. Infectivity Determinants of the Hepatitis B Virus Pre-S Domain Are Confined to the N-Terminal 75 Amino Acid Residues. J. Virol. 2007, 81, 5841–5849. [Google Scholar] [CrossRef] [Green Version]
- Chouteau, P.; Le Seyec, J.; Cannie, I.; Nassal, M.; Guguen-Guillouzo, C.; Gripon, P. A short N-proximal region in the large envelope protein harbors a determinant that contributes to the species specificity of human hepatitis B virus. J. Virol. 2001, 75, 11565–11572. [Google Scholar] [CrossRef] [Green Version]
- Le Duff, Y.; Blanchet, M.; Sureau, C. The pre-S1 and antigenic loop infectivity determinants of the hepatitis B virus envelope proteins are functionally independent. J. Virol. 2009, 83, 12443–12451. [Google Scholar] [CrossRef] [Green Version]
- Le Seyec, J.; Chouteau, P.; Cannie, I.; Guguen-Guillouzo, C.; Gripon, P. Infection process of the hepatitis B virus depends on the presence of a defined sequence in the pre-S1 domain. J. Virol. 1999, 73, 2052–2057. [Google Scholar] [CrossRef] [Green Version]
- Barrera, A.; Guerra, B.; Notvall, L.; Lanford, R.E. Mapping of the hepatitis B virus pre-S1 domain involved in receptor recognition. J. Virol. 2005, 79, 9786–9798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engelke, M.; Mills, K.; Seitz, S.; Simon, P.; Gripon, P.; Schnölzer, M.; Urban, S. Characterization of a hepatitis B and hepatitis delta virus receptor binding site. Hepatol. Baltim. Md. 2006, 43, 750–760. [Google Scholar] [CrossRef]
- Glebe, D.; Urban, S.; Knoop, E.V.; Cag, N.; Krass, P.; Grün, S.; Bulavaite, A.; Sasnauskas, K.; Gerlich, W.H. Mapping of the hepatitis B virus attachment site by use of infection-inhibiting preS1 lipopeptides and tupaia hepatocytes. Gastroenterology 2005, 129, 234–245. [Google Scholar] [CrossRef]
- Murayama, A.; Yamada, N.; Osaki, Y.; Shiina, M.; Aly, H.H.; Iwamoto, M.; Tsukuda, S.; Watashi, K.; Matsuda, M.; Suzuki, R.; et al. N-terminal PreS1 Sequence Regulates Efficient Infection of Cell Culture-generated Hepatitis B Virus. Hepatology 2020, hep.31308. [Google Scholar] [CrossRef] [PubMed]
- Treichel, U.; Meyer zum Buschenfelde, K.-H.; Stockert, R.J.; Poralla, T.; Gerken, G. The Asialoglycoprotein Receptor Mediates Hepatic Binding and Uptake of Natural Hepatitis B Virus Particles Derived From Viraemic Carriers. J. Gen. Virol. 1994, 75, 3021–3029. [Google Scholar] [CrossRef] [PubMed]
- Franco, A.; Paroli, M.; Testa, U.; Benvenuto, R.; Peschle, C.; Balsano, F.; Barnaba, V. Transferrin receptor mediates uptake and presentation of hepatitis B envelope antigen by T lymphocytes. J. Exp. Med. 1992, 175, 1195–1205. [Google Scholar] [CrossRef] [Green Version]
- Neurath, A.R.; Strick, N.; Sproul, P. Search for hepatitis B virus cell receptors reveals binding sites for interleukin 6 on the virus envelope protein. J. Exp. Med. 1992, 175, 461–469. [Google Scholar] [CrossRef] [Green Version]
- Pontisso, P.; Petit, M.A.; Bankowski, M.J.; Peeples, M.E. Human liver plasma membranes contain receptors for the hepatitis B virus pre-S1 region and, via polymerized human serum albumin, for the pre-S2 region. J. Virol. 1989, 63, 1981–1988. [Google Scholar] [CrossRef] [Green Version]
- Anwer, M.S.; Stieger, B. Sodium-dependent bile salt transporters of the SLC10A transporter family: More than solute transporters. Pflüg. Arch. Eur. J. Physiol. 2014, 466, 77–89. [Google Scholar] [CrossRef] [Green Version]
- Kullak-Ublick, G.; Glasa, J.; Boker, C.; Oswald, M.; Grutzner, U.; Hagenbuch, B.; Stieger, B.; Meier, P.; Beuers, U.; Kramer, W.; et al. Chlorambucil-taurocholate is transported by bile acid carriers expressed in human hepatocellular carcinomas. Gastroenterology 1997, 113, 1295–1305. [Google Scholar] [CrossRef]
- Xia, Y.; Carpentier, A.; Cheng, X.; Block, P.D.; Zhao, Y.; Zhang, Z.; Protzer, U.; Liang, T.J. Human stem cell-derived hepatocytes as a model for hepatitis B virus infection, spreading and virus-host interactions. J. Hepatol. 2017, 66, 494–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.-Z.; Qian, X.-J.; Zhao, P.; Qi, Z.-T. How hepatitis C virus invades hepatocytes: The mystery of viral entry. World J. Gastroenterol. WJG 2014, 20, 3457–3467. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Qu, B.; Zhang, F.; Zhang, C.; Deng, W.; Dao Thi, V.L.; Xia, Y. Stem Cell-Derived Hepatocyte-Like Cells as Model for Viral Hepatitis Research. Stem Cells Int. 2019, 2019, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwamoto, M.; Saso, W.; Sugiyama, R.; Ishii, K.; Ohki, M.; Nagamori, S.; Suzuki, R.; Aizaki, H.; Ryo, A.; Yun, J.-H.; et al. Epidermal growth factor receptor is a host-entry cofactor triggering hepatitis B virus internalization. Proc. Natl. Acad. Sci. USA 2019, 116, 8487–8492. [Google Scholar] [CrossRef] [Green Version]
- Zhao, P.; Yang, X.; Qi, S.; Liu, H.; Jiang, H.; Hoppmann, S.; Cao, Q.; Chua, M.-S.; So, S.K.; Cheng, Z. Molecular imaging of hepatocellular carcinoma xenografts with epidermal growth factor receptor targeted affibody probes. BioMed Res. Int. 2013, 2013, 759057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Q.; Zhang, F.; Duan, L.; Wang, B.; Ye, Y.; Li, P.; Li, D.; Yang, S.; Zhou, L.; Chen, W. E-cadherin Plays a Role in Hepatitis B Virus Entry Through Affecting Glycosylated Sodium-Taurocholate Cotransporting Polypeptide Distribution. Front. Cell. Infect. Microbiol. 2020, 10, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulze, A.; Mills, K.; Weiss, T.S.; Urban, S. Hepatocyte polarization is essential for the productive entry of the hepatitis B virus. Hepatology 2012, 55, 373–383. [Google Scholar] [CrossRef]
- Mee, C.J.; Grove, J.; Harris, H.J.; Hu, K.; Balfe, P.; McKeating, J.A. Effect of Cell Polarization on Hepatitis C Virus Entry. J. Virol. 2008, 82, 461–470. [Google Scholar] [CrossRef] [Green Version]
- Mee, C.J.; Harris, H.J.; Farquhar, M.J.; Wilson, G.; Reynolds, G.; Davis, C.; van IJzendoorn, S.C.D.; Balfe, P.; McKeating, J.A. Polarization Restricts Hepatitis C Virus Entry into HepG2 Hepatoma Cells. J. Virol. 2009, 83, 6211–6221. [Google Scholar] [CrossRef] [Green Version]
- Belouzard, S.; Danneels, A.; Fénéant, L.; Séron, K.; Rouillé, Y.; Dubuisson, J. Entry and Release of Hepatitis C Virus in Polarized Human Hepatocytes. J. Virol. 2017, 91, e00478-17. [Google Scholar] [CrossRef] [Green Version]
- Bremer, C.M.; Bung, C.; Kott, N.; Hardt, M.; Glebe, D. Hepatitis B virus infection is dependent on cholesterol in the viral envelope. Cell. Microbiol. 2009, 11, 249–260. [Google Scholar] [CrossRef] [PubMed]
- Iwamoto, M.; Saso, W.; Nishioka, K.; Ohashi, H.; Sugiyama, R.; Ryo, A.; Ohki, M.; Yun, J.-H.; Park, S.-Y.; Ohshima, T.; et al. The machinery for endocytosis of epidermal growth factor receptor coordinates the transport of incoming hepatitis B virus to the endosomal network. J. Biol. Chem. 2020, 295, 800–807. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, B.; McNiven, M.A. Importance of Endocytic Pathways in Liver Function and Disease. Compr. Physiol. 2014, 4, 1403–1417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stross, C.; Kluge, S.; Weissenberger, K.; Winands, E.; Häussinger, D.; Kubitz, R. A dileucine motif is involved in plasma membrane expression and endocytosis of rat sodium taurocholate cotransporting polypeptide (Ntcp). Am. J. Physiol. Gastrointest. Liver Physiol. 2013, 305, G722–G730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oess, S.; Hildt, E. Novel cell permeable motif derived from the PreS2-domain of hepatitis-B virus surface antigens. Gene Ther. 2000, 7, 750–758. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Crespo, I.; Gomez-Gutierrez, J.; Nieto, M.; Peterson, D.L.; Gavilanes, F. Prediction of a putative fusion peptide in the S protein of hepatitis B virus. J. Gen. Virol. 1994, 75, 637–639. [Google Scholar] [CrossRef]
- Delgado, C.L.; Núñez, E.; Yélamos, B.; Gómez-Gutiérrez, J.; Peterson, D.L.; Gavilanes, F. Study of the putative fusion regions of the preS domain of hepatitis B virus. Biochim. Biophys. Acta BBA Biomembr. 2015, 1848, 895–906. [Google Scholar] [CrossRef] [Green Version]
- Somiya, M.; Sasaki, Y.; Matsuzaki, T.; Liu, Q.; Iijima, M.; Yoshimoto, N.; Niimi, T.; Maturana, A.D.; Kuroda, S. Intracellular trafficking of bio-nanocapsule–liposome complex: Identification of fusogenic activity in the pre-S1 region of hepatitis B virus surface antigen L protein. J. Control. Release 2015, 212, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Funk, A.; Mhamdi, M.; Hohenberg, H.; Will, H.; Sirma, H. pH-independent entry and sequential endosomal sorting are major determinants of hepadnaviral infection in primary hepatocytes. Hepatology 2006, 44, 685–693. [Google Scholar] [CrossRef]
- Rigg, R.J.; Schaller, H. Duck hepatitis B virus infection of hepatocytes is not dependent on low pH. J. Virol. 1992, 66, 2829–2836. [Google Scholar] [CrossRef] [Green Version]
- Yoshimori, T.; Yamamoto, A.; Moriyama, Y.; Futai, M.; Tashiro, Y. Bafilomycin A1, a specific inhibitor of vacuolar-type H(+)-ATPase, inhibits acidification and protein degradation in lysosomes of cultured cells. J. Biol. Chem. 1991, 266, 17707–17712. [Google Scholar] [PubMed]
- Chojnacki, J.; Anderson, D.A.; Grgacic, E.V.L. A Hydrophobic Domain in the Large Envelope Protein Is Essential for Fusion of Duck Hepatitis B Virus at the Late Endosome. J. Virol. 2005, 79, 14945–14955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watashi, K.; Sluder, A.; Daito, T.; Matsunaga, S.; Ryo, A.; Nagamori, S.; Iwamoto, M.; Nakajima, S.; Tsukuda, S.; Borroto-Esoda, K.; et al. Cyclosporin A and its analogs inhibit hepatitis B virus entry into cultured hepatocytes through targeting a membrane transporter, sodium taurocholate cotransporting polypeptide (NTCP): Watashi et al. Hepatology 2014, 59, 1726–1737. [Google Scholar] [CrossRef] [PubMed]
- Macovei, A.; Petrareanu, C.; Lazar, C.; Florian, P.; Branza-Nichita, N. Regulation of Hepatitis B Virus Infection by Rab5, Rab7, and the Endolysosomal Compartment. J. Virol. 2013, 87, 6415–6427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Somsel Rodman, J.; Wandinger-Ness, A. Rab GTPases coordinate endocytosis. J. Cell Sci. 2000, 113, 183–192. [Google Scholar]
- Urban, S.; Bartenschlager, R.; Kubitz, R.; Zoulim, F. Strategies to inhibit entry of HBV and HDV into hepatocytes. Gastroenterology 2014, 147, 48–64. [Google Scholar] [CrossRef]
- Hong, H.J.; Ryu, C.J.; Hur, H.; Kim, S.; Oh, H.K.; Oh, M.S.; Park, S.Y. In vivo neutralization of hepatitis B virus infection by an anti-preS1 humanized antibody in chimpanzees. Virology 2004, 318, 134–141. [Google Scholar] [CrossRef] [Green Version]
- Wi, J.; Jeong, M.S.; Hong, H.J. Construction and Characterization of an Anti-Hepatitis B Virus preS1 Humanized Antibody that Binds to the Essential Receptor Binding Site. J. Microbiol. Biotechnol. 2017, 27, 1336–1344. [Google Scholar] [CrossRef] [Green Version]
- Eke, A.C.; Eleje, G.U.; Eke, U.A.; Xia, Y.; Liu, J. Hepatitis B immunoglobulin during pregnancy for prevention of mother-to-child transmission of hepatitis B virus. Cochrane Database Syst. Rev. 2017. [Google Scholar] [CrossRef]
- Katz, L.H.; Paul, M.; Guy, D.G.; Tur-Kaspa, R. Prevention of recurrent hepatitis B virus infection after liver transplantation: Hepatitis B immunoglobulin, antiviral drugs, or both? Systematic review and meta-analysis: Post liver transplant hepatitis B prophylaxis. Transpl. Infect. Dis. 2010, 12, 292–308. [Google Scholar] [CrossRef]
- Krepstakies, M.; Lucifora, J.; Nagel, C.-H.; Zeisel, M.B.; Holstermann, B.; Hohenberg, H.; Kowalski, I.; Gutsmann, T.; Baumert, T.F.; Brandenburg, K.; et al. A New Class of Synthetic Peptide Inhibitors Blocks Attachment and Entry of Human Pathogenic Viruses. J. Infect. Dis. 2012, 205, 1654–1664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamas Longarela, O.; Schmidt, T.T.; Schöneweis, K.; Romeo, R.; Wedemeyer, H.; Urban, S.; Schulze, A. Proteoglycans Act as Cellular Hepatitis Delta Virus Attachment Receptors. PLoS ONE 2013, 8, e58340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petcu, D.J.; Aldrich, C.E.; Coates, L.; Taylor, J.M.; Mason, W.S. Suramin inhibits in vitro infection by duck hepatitis B virus, Rous sarcoma virus, and hepatitis delta virus. Virology 1988, 167, 385–392. [Google Scholar] [CrossRef]
- Xia, Y.; Cheng, X.; Blossey, C.K.; Wisskirchen, K.; Esser, K.; Protzer, U. Secreted Interferon-Inducible Factors Restrict Hepatitis B and C Virus Entry In Vitro. J. Immunol. Res. 2017, 2017, 1–12. [Google Scholar] [CrossRef]
- Allweiss, L.; Volz, T.; Giersch, K.; Kah, J.; Raffa, G.; Petersen, J.; Lohse, A.W.; Beninati, C.; Pollicino, T.; Urban, S.; et al. Proliferation of primary human hepatocytes and prevention of hepatitis B virus reinfection efficiently deplete nuclear cccDNA in vivo. Gut 2018, 67, 542–552. [Google Scholar] [CrossRef]
- Tu, T.; Urban, S. Virus entry and its inhibition to prevent and treat hepatitis B and hepatitis D virus infections. Curr. Opin. Virol. 2018, 30, 68–79. [Google Scholar] [CrossRef]
- Tsukuda, S.; Watashi, K.; Hojima, T.; Isogawa, M.; Iwamoto, M.; Omagari, K.; Suzuki, R.; Aizaki, H.; Kojima, S.; Sugiyama, M.; et al. A new class of hepatitis B and D virus entry inhibitors, proanthocyanidin and its analogs, that directly act on the viral large surface proteins. Hepatology 2017, 65, 1104–1116. [Google Scholar] [CrossRef] [Green Version]
- Yan, H.; Peng, B.; Liu, Y.; Xu, G.; He, W.; Ren, B.; Jing, Z.; Sui, J.; Li, W. Viral Entry of Hepatitis B and D Viruses and Bile Salts Transportation Share Common Molecular Determinants on Sodium Taurocholate Cotransporting Polypeptide. J. Virol. 2014, 88, 3273–3284. [Google Scholar] [CrossRef] [Green Version]
- Veloso Alves Pereira, I.; Buchmann, B.; Sandmann, L.; Sprinzl, K.; Schlaphoff, V.; Döhner, K.; Vondran, F.; Sarrazin, C.; Manns, M.P.; Pinto Marques Souza de Oliveira, C.; et al. Primary Biliary Acids Inhibit Hepatitis D Virus (HDV) Entry into Human Hepatoma Cells Expressing the Sodium-Taurocholate Cotransporting Polypeptide (NTCP). PLoS ONE 2015, 10, e0117152. [Google Scholar] [CrossRef] [Green Version]
- König, A.; Döring, B.; Mohr, C.; Geipel, A.; Geyer, J.; Glebe, D. Kinetics of the bile acid transporter and hepatitis B virus receptor Na+/taurocholate cotransporting polypeptide (NTCP) in hepatocytes. J. Hepatol. 2014, 61, 867–875. [Google Scholar] [CrossRef] [Green Version]
- Lucifora, J.; Esser, K.; Protzer, U. Ezetimibe blocks hepatitis B virus infection after virus uptake into hepatocytes. Antivir. Res. 2013, 97, 195–197. [Google Scholar] [CrossRef] [PubMed]
- Ko, C.; Park, W.-J.; Park, S.; Kim, S.; Windisch, M.P.; Ryu, W.-S. The FDA approved drug irbesartan inhibits HBV-infection in HepG2 cells stably expressing sodium taurocholate co-transporting polypeptide. Antivir. Ther. 2015, 20, 835–842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bogomolov, P.; Alexandrov, A.; Voronkova, N.; Macievich, M.; Kokina, K.; Petrachenkova, M.; Lehr, T.; Lempp, F.A.; Wedemeyer, H.; Haag, M.; et al. Treatment of chronic hepatitis D with the entry inhibitor myrcludex B: First results of a phase Ib/IIa study. J. Hepatol. 2016, 65, 490–498. [Google Scholar] [CrossRef] [PubMed]
- Volz, T.; Allweiss, L.; ḾBarek, M.B.; Warlich, M.; Lohse, A.W.; Pollok, J.M.; Alexandrov, A.; Urban, S.; Petersen, J.; Lütgehetmann, M.; et al. The entry inhibitor Myrcludex-B efficiently blocks intrahepatic virus spreading in humanized mice previously infected with hepatitis B virus. J. Hepatol. 2013, 58, 861–867. [Google Scholar] [CrossRef]
- Shimura, S.; Watashi, K.; Fukano, K.; Peel, M.; Sluder, A.; Kawai, F.; Iwamoto, M.; Tsukuda, S.; Takeuchi, J.S.; Miyake, T.; et al. Cyclosporin derivatives inhibit hepatitis B virus entry without interfering with NTCP transporter activity. J. Hepatol. 2017, 66, 685–692. [Google Scholar] [CrossRef]
- Kaneko, M.; Watashi, K.; Kamisuki, S.; Matsunaga, H.; Iwamoto, M.; Kawai, F.; Ohashi, H.; Tsukuda, S.; Shimura, S.; Suzuki, R.; et al. A Novel Tricyclic Polyketide, Vanitaracin A, Specifically Inhibits the Entry of Hepatitis B and D Viruses by Targeting Sodium Taurocholate Cotransporting Polypeptide. J. Virol. 2015, 89, 11945–11953. [Google Scholar] [CrossRef] [Green Version]
- Tsukuda, S.; Watashi, K.; Iwamoto, M.; Suzuki, R.; Aizaki, H.; Okada, M.; Sugiyama, M.; Kojima, S.; Tanaka, Y.; Mizokami, M.; et al. Dysregulation of Retinoic Acid Receptor Diminishes Hepatocyte Permissiveness to Hepatitis B Virus Infection through Modulation of Sodium Taurocholate Cotransporting Polypeptide (NTCP) Expression. J. Biol. Chem. 2015, 290, 5673–5684. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Y.; Liu, C.; Tang, W.; Zhang, H.; Chen, X. Evans Blue Inhibits HBV Replication Through a Dual Antiviral Mechanism by Targeting Virus Binding and Capsid Assembly. Front. Microbiol. 2019, 10, 2638. [Google Scholar] [CrossRef]
- Donkers, J.M.; Zehnder, B.; van Westen, G.J.P.; Kwakkenbos, M.J.; IJzerman, A.P.; Oude Elferink, R.P.J.; Beuers, U.; Urban, S.; van de Graaf, S.F.J. Reduced hepatitis B and D viral entry using clinically applied drugs as novel inhibitors of the bile acid transporter NTCP. Sci. Rep. 2017, 7, 15307. [Google Scholar] [CrossRef] [Green Version]
- Donkers, J.M.; Appelman, M.D.; van de Graaf, S.F.J. Mechanistic insights into the inhibition of NTCP by myrcludex B. JHEP Rep. 2019, 1, 278–285. [Google Scholar] [CrossRef] [Green Version]
- Si, J.; Yu, C.; Guo, Y.; Bian, Z.; Meng, R.; Yang, L.; Chen, Y.; Jin, J.; Liu, J.; Guo, Z.; et al. Chronic hepatitis B virus infection and total and cause-specific mortality: A prospective cohort study of 0.5 million people. BMJ Open 2019, 9, e027696. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Viruses | References |
|---|---|
| Clathrin-Mediated Endocytosis | |
| influenza A virus | [8,9] |
| hepatitis C virus | [10,11] |
| dengue virus | [12,13] |
| vesicular stomatitis virus hepatitis B virus * | [14,15] [16,17,18] |
| Caveolae/Lipid Raft- Mediated Endocytosis | |
| simian virus 40 hepatitis B virus * | [19,20] [21] |
| Macropinocytosis | |
| ebola virus | [22,23] |
| vaccinia virus | [24,25] |
| adenovirus 3 | [26] |
| Other pathways | |
| rotavirus—IL-2 pathway | [27,28] |
| adenovirus 2—CLIC-GEEC pathway | [29] |
| coxsackievirus A9—Arf6 pathway | [30] |
| enterovirus 71—endophilin pathway | [31] |
| Class | Substance | Target | Status | References |
|---|---|---|---|---|
| Attachment inhibitors | Heparin | S, M, L | FDA-approved | [51] |
| Suramin | S, M, L | FDA-approved | [103] | |
| Proanthocyanidin | PreS1 | Preclinical | [107] | |
| SALP | HSPGs | Preclinical | [101] | |
| NTCP substrate inhibitors | Taurocholic acid, UDCA, TUDCA, GUDCA | NTCP | / | [108,109,110] |
| Ezetimib | NTCP/NPC1L1 | FDA-approved | [111] | |
| Irbesartan | NTCP | FDA-approved | [112] | |
| NTCP inhibitors | Myrcludex B | NTCP/Interferes slightly with bile acid uptake | Phase III | [113,114] |
| CsA | NTCP/Interferes strongly with bile acid uptake | FDA-approved | [93] | |
| SCY450 | NTCP/No interference with bile acid uptake | Preclinical | [115] | |
| SCY995 | NTCP/No interference with bile acid uptake | Preclinical | [115] | |
| Vanitaracin A | NTCP/Interferes strongly with bile acid uptake | Preclinical | [116] | |
| Ro41-5253 | NTCP/Antagonist of retinoic acid receptor which regulates NTCP expression | Preclinical | [117] | |
| Evans blue | NTCP | FDA-approved | [118] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Herrscher, C.; Roingeard, P.; Blanchard, E. Hepatitis B Virus Entry into Cells. Cells 2020, 9, 1486. https://doi.org/10.3390/cells9061486
Herrscher C, Roingeard P, Blanchard E. Hepatitis B Virus Entry into Cells. Cells. 2020; 9(6):1486. https://doi.org/10.3390/cells9061486
Chicago/Turabian StyleHerrscher, Charline, Philippe Roingeard, and Emmanuelle Blanchard. 2020. "Hepatitis B Virus Entry into Cells" Cells 9, no. 6: 1486. https://doi.org/10.3390/cells9061486
APA StyleHerrscher, C., Roingeard, P., & Blanchard, E. (2020). Hepatitis B Virus Entry into Cells. Cells, 9(6), 1486. https://doi.org/10.3390/cells9061486