TRPV4 Inhibition and CRISPR-Cas9 Knockout Reduce Inflammation Induced by Hyperphysiological Stretching in Human Annulus Fibrosus Cells

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Human AF Cell Isolation and Culture
2.2. Generation of CRISPR-Cas9 TRPV4 Knockout (KO) Cells
2.3. Cyclic Stretching
2.4. RNA Extraction and RT-qPCR
2.5. ELISA of Conditioned Medium
2.6. Western Blot
2.7. Immunocytochemistry
2.8. Statistical Analysis
3. Results
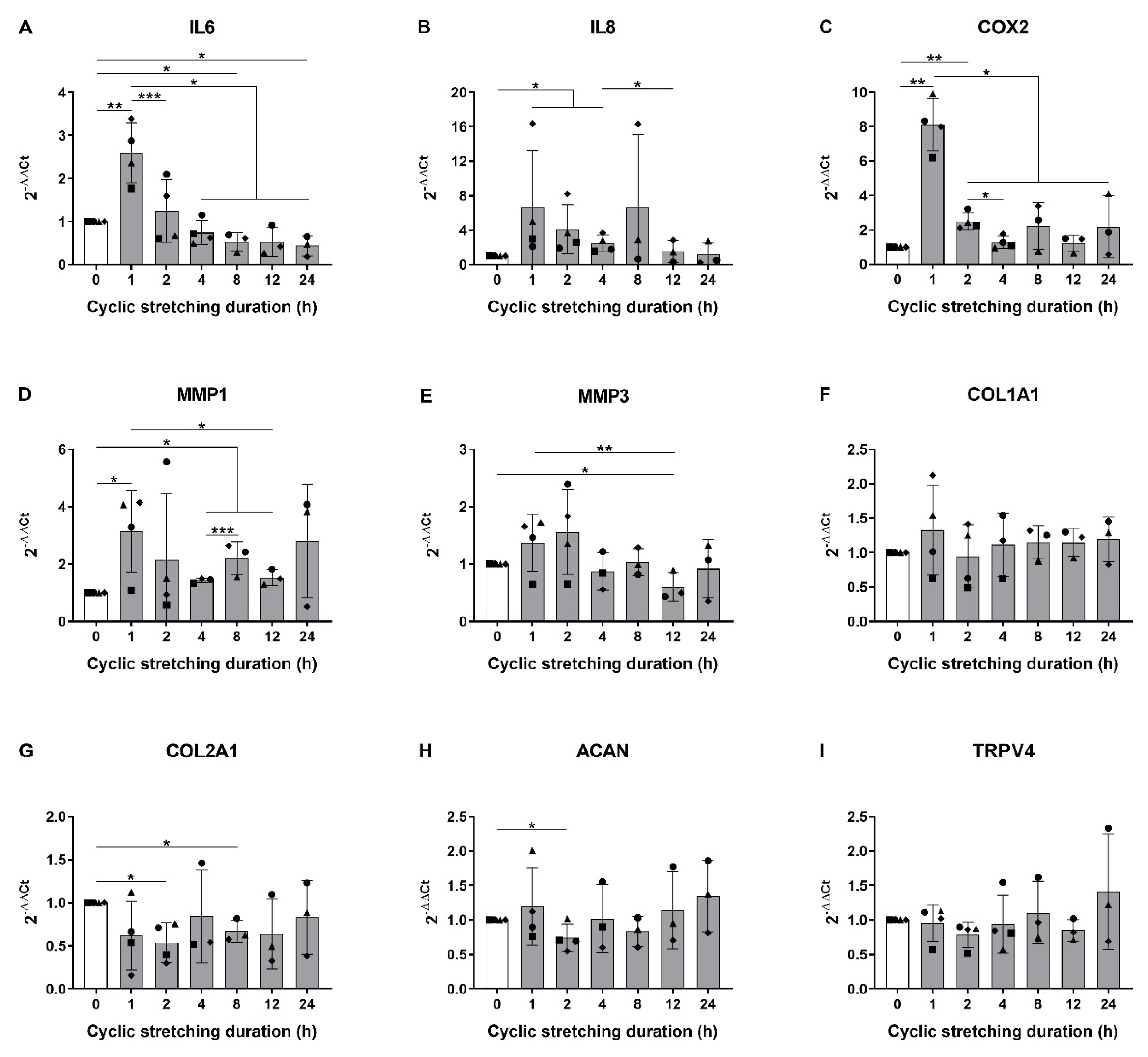
3.1. Hyperphysiological Cyclic Stretching Induces Gene Expression of Pro-Inflammatory Mediators in Human AF Cells
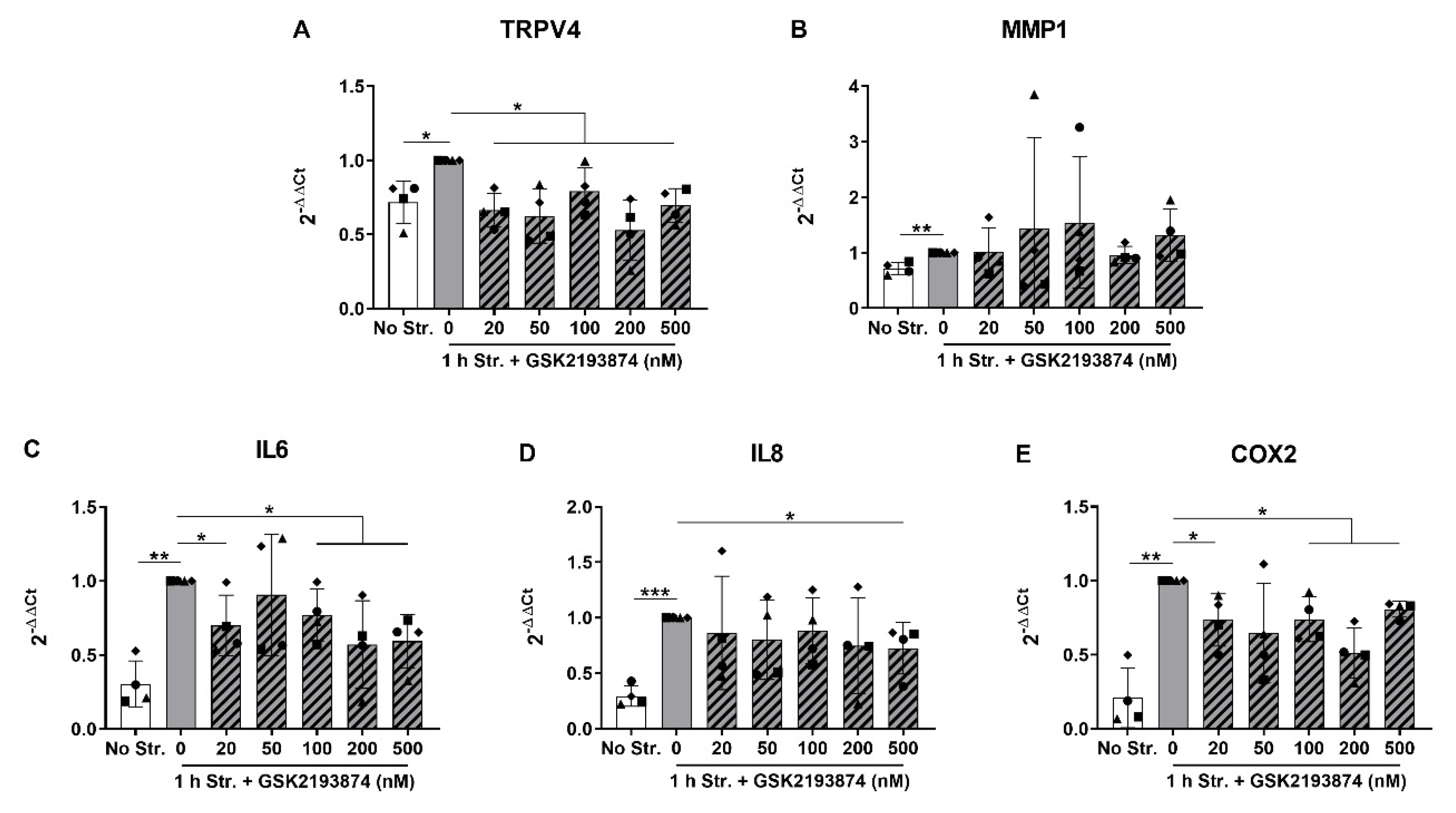
3.2. Pharmacological Inhibition of TRPV4 Reduces Stretch-Induced Gene Expression of Pro-Inflammatory Mediators
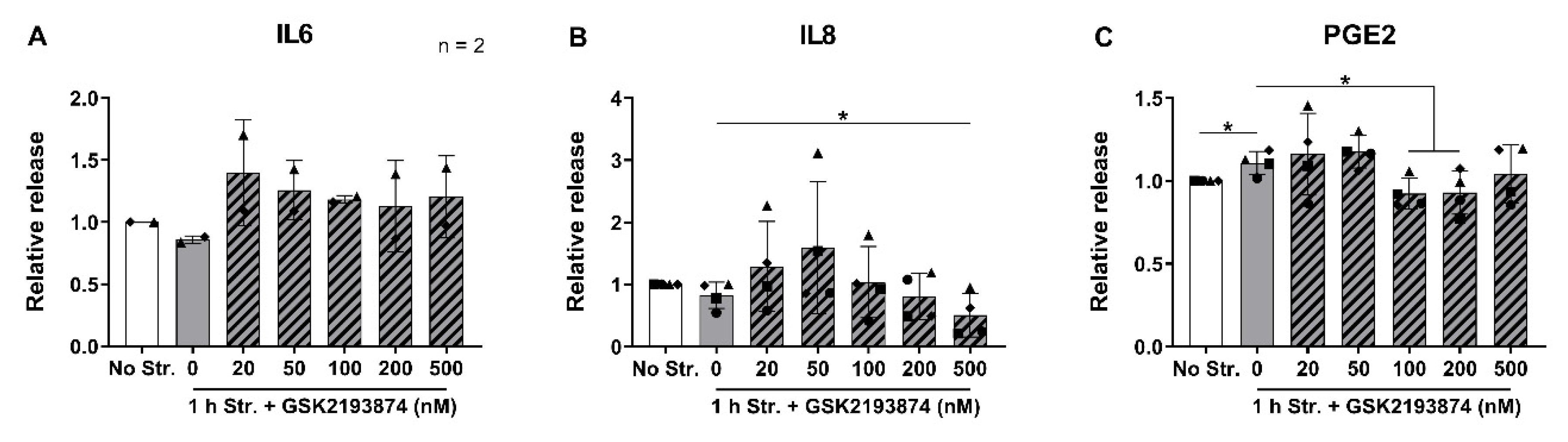
3.3. Pharmacological Inhibition of TRPV4 Downregulates the Release of IL8 and PGE2
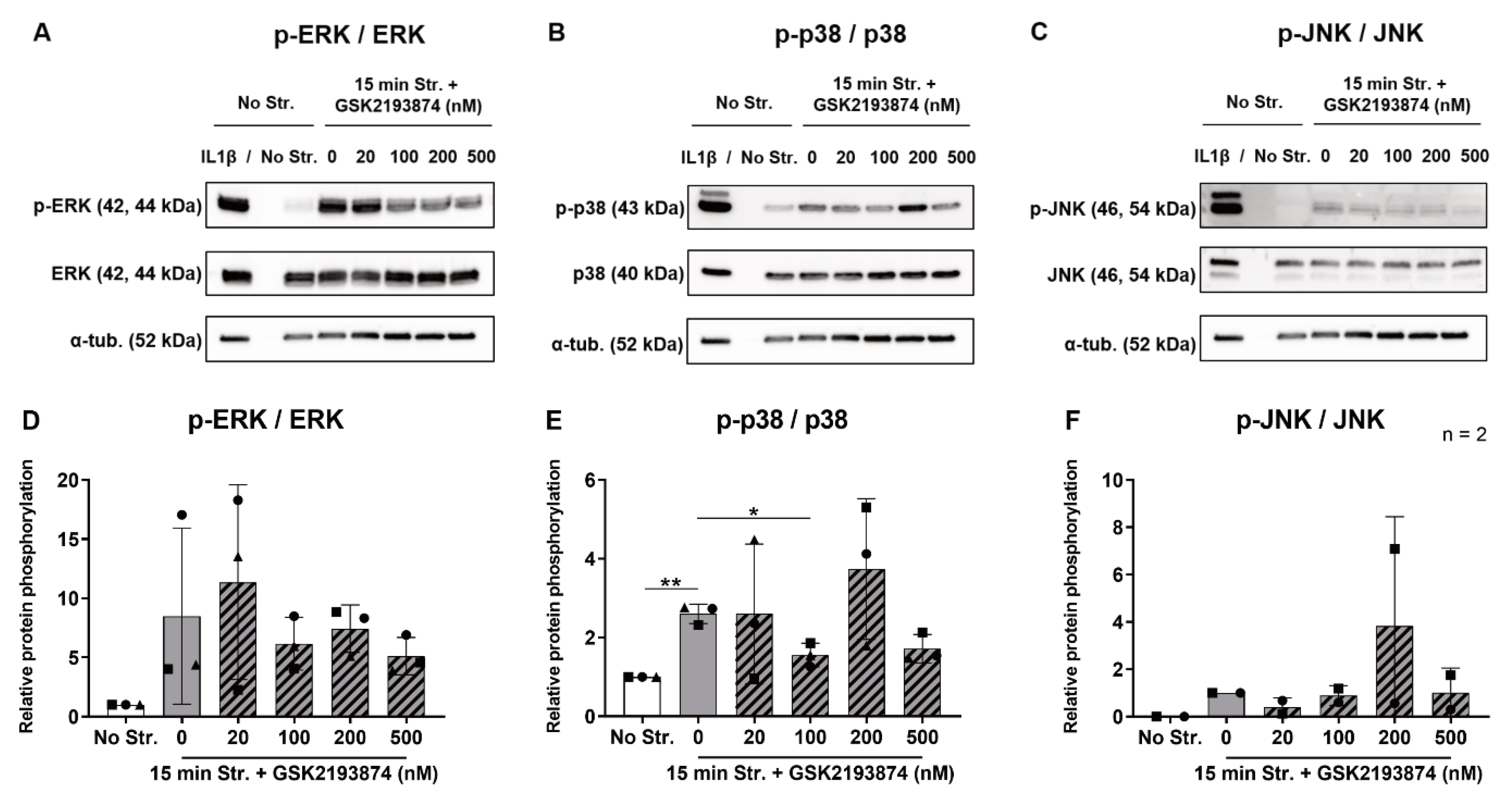
3.4. Pharmacological Inhibition of TRPV4 Reduces Stretch-Induced p38 Phosphorylation
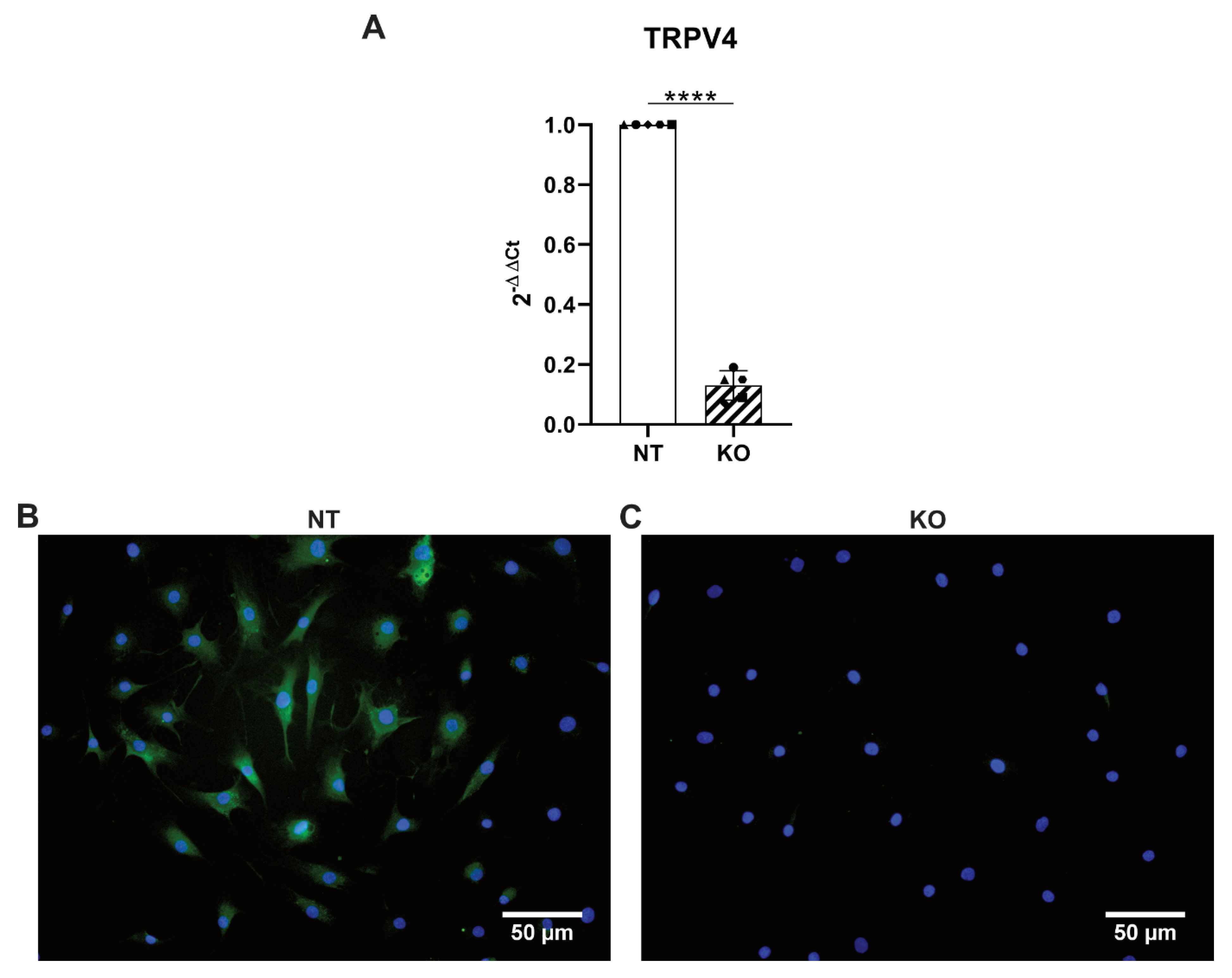
3.5. CRISPR-Cas9 Knocks Out TRPV4 in Human Primary AF Cells
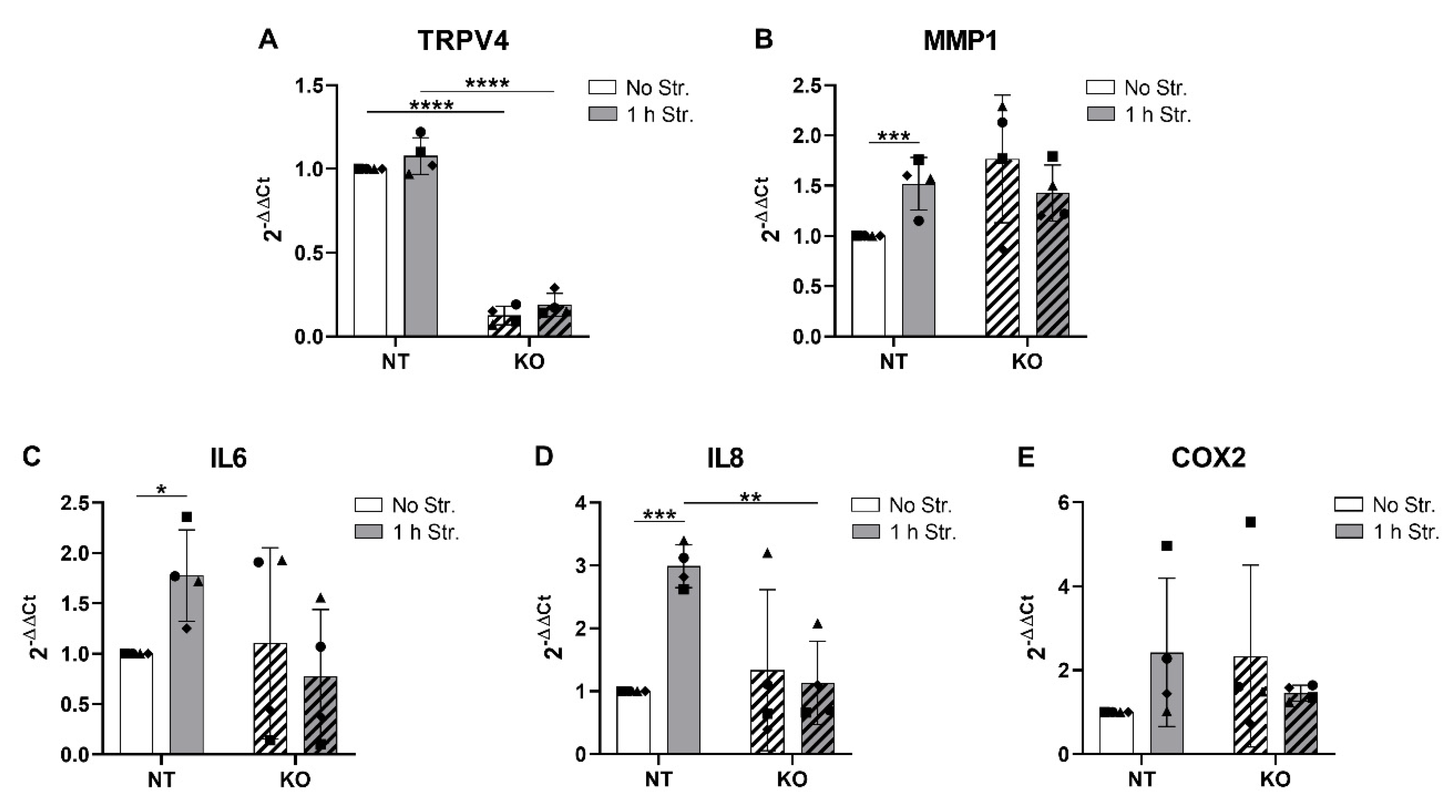
3.6. CRISPR-Cas9 KO of TRPV4 Prevents Stretch-Induced Gene Expression of IL8
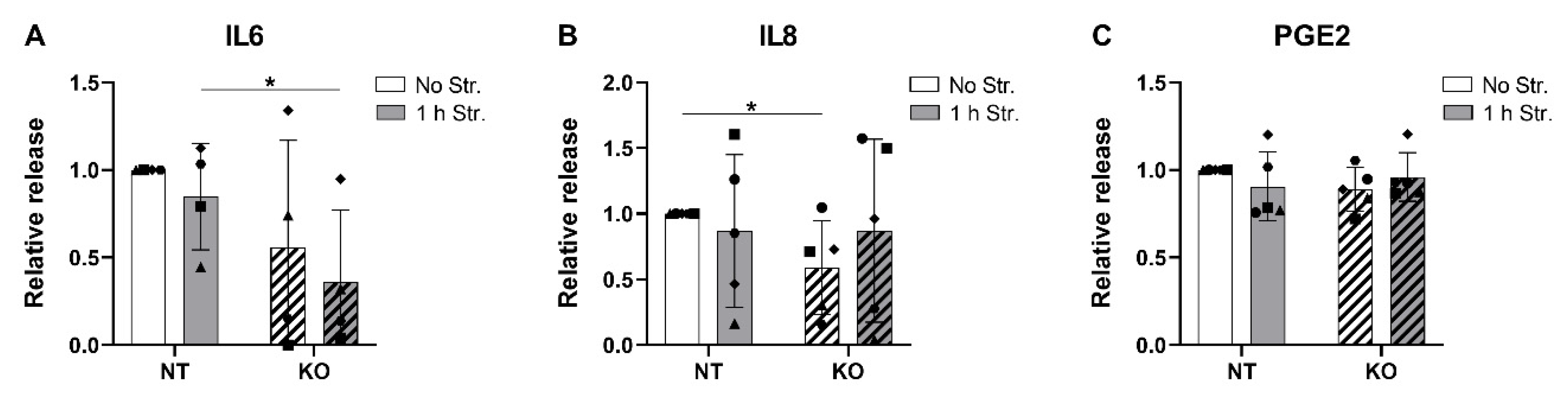
3.7. CRISPR-Cas9 KO of TRPV4 Downregulates the Release of IL6 and IL8
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ingber, D.E. Mechanobiology and diseases of mechanotransduction. Ann. Med. 2003, 35, 564–577. [Google Scholar] [CrossRef] [PubMed]
- Bao, S. Mechanical stress. In The Handbook of Clinical Neurology; Elsevier: Amsterdam, The Netherlands, 2015; Volume 131, pp. 367–396. [Google Scholar]
- Guo, X.E.; Hung, C.T.; Sandell, L.J.; Silva, M.J. Musculoskeletal mechanobiology: A new era for MechanoMedicine. J. Orthop. Res. 2018, 36, 531–532. [Google Scholar] [CrossRef] [Green Version]
- Fearing, B.V.; Hernandez, P.A.; Setton, L.A.; Chahine, N. Mechanotransduction and cell biomechanics of the intervertebral disc. JOR Spine 2018, 1, e1026. [Google Scholar] [CrossRef] [PubMed]
- Chan, S.C.W.; Ferguson, S.J.; Gantenbein-Ritter, B. The effects of dynamic loading on the intervertebral disc. Eur. Spine J. 2011, 20, 1796–1812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, M.A.; Roughley, P.J. What is intervertebral disc degeneration, and what causes it? Spine 2006, 31, 2151–2161. [Google Scholar] [CrossRef] [Green Version]
- Vergroesen, P.-P.; Kingma, I.; Emanuel, K.; Hoogendoorn, R.; Welting, T.; Van Royen, B.J.; Van Dieën, J.H.; Smit, T.H. Mechanics and biology in intervertebral disc degeneration: A vicious circle. Osteoarthr. Cart. 2015, 23, 1057–1070. [Google Scholar] [CrossRef] [Green Version]
- Likhitpanichkul, M.; Torre, O.M.; Gruen, J.; Walter, B.A.; Hecht, A.C.; Iatridis, J.C. Do mechanical strain and TNF-alpha interact to amplify pro-inflammatory cytokine production in human annulus fibrosus cells? J. Biomech. 2016, 49, 1214–1220. [Google Scholar] [CrossRef] [Green Version]
- Tisherman, R.; Coelho, P.; Phillibert, D.; Wang, D.; Dong, Q.; Vo, N.; Kang, J.; Sowa, J. NF-kappaB signaling pathway in controlling intervertebral disk cell response to inflammatory and mechanical stressors. Phys. Ther. 2016, 96, 704–711. [Google Scholar] [CrossRef] [Green Version]
- Balagué, F.; Mannion, A.F.; Pellise, F.; Cedraschi, C. Non-specific low back pain. Lancet 2012, 379, 482–491. [Google Scholar] [CrossRef] [Green Version]
- Airaksinen, O.; Brox, J.I.; Cedraschi, C.; Hildebrandt, J.; Klaber-Moffett, J.; Kovacs, F.; Mannion, A.F.; Reis, S.; Staal, J.B.; Ursin, H.; et al. Chapter 4. European guidelines for the management of chronic nonspecific low back pain. Eur. Spine J. 2006, 15, S192–S300. [Google Scholar] [CrossRef] [Green Version]
- Torre, O.M.; Mroz, V.; Bartelstein, M.K.; Huang, A.H.; Iatridis, J.C. Annulus fibrosus cell phenotypes in homeostasis and injury: Implications for regenerative strategies. Ann. N. Y. Acad. Sci. 2019, 1442, 61–78. [Google Scholar] [CrossRef] [PubMed]
- Shengjie, W.; Li, J.; Tian, J.; Yu, Z.; Gao, K.; Shao, J.; Li, A.; Xing, S.; Dong, Y.; Li, Z. High amplitude and low frequency cyclic mechanical strain promotes degeneration of human nucleus pulposus cells via the NF-κB p65 pathway. J. Cell. Physiol. 2018, 233, 7206–7216. [Google Scholar]
- Sowa, G.; Coelho, P.; Vo, N.; Bedison, R.; Chiao, A.; Davies, C.; Studer, R.; Kang, J. Determination of annulus fibrosus cell response to tensile strain as a function of duration, magnitude, and frequency. J. Orthop. Res. 2011, 29, 1275–1283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyamoto, H.; Doita, M.; Nishida, K.; Yamamoto, T.; Sumi, M.; Kurosaka, M. Effects of cyclic mechanical stress on the production of inflammatory agents by nucleus pulposus and anulus fibrosus derived cells in vitro. Spine 2006, 31, 4–9. [Google Scholar] [CrossRef] [PubMed]
- Gawri, R.; Rosenzweig, D.H.; Krock, E.; A Ouellet, J.; Stone, L.S.; Quinn, T.M.; Haglund, L. High mechanical strain of primary intervertebral disc cells promotes secretion of inflammatory factors associated with disc degeneration and pain. Arthritis Res. Ther. 2014, 16, R21. [Google Scholar] [CrossRef] [Green Version]
- Pratsinis, H.; Papadopoulou, A.; Neidlinger-Wilke, C.; Brayda-Bruno, M.; Wilke, H.-J.; Kletsas, D.; Information, P.E.K.F.C. Cyclic tensile stress of human annulus fibrosus cells induces MAPK activation: Involvement in proinflammatory gene expression. Osteoarthr. Cartil. 2016, 24, 679–687. [Google Scholar] [CrossRef] [Green Version]
- Moran, M.M.; McAlexander, M.A.; Bíró, T.; Szallasi, A. Transient receptor potential channels as therapeutic targets. Nat. Rev. Drug Discov. 2011, 10, 601–620. [Google Scholar] [CrossRef]
- Krupkova, O.; Zvick, J.; Wuertz-Kozak, K. The role of transient receptor potential channels in joint diseases. Eur. Cells Mater. 2017, 34, 180–201. [Google Scholar] [CrossRef]
- McNulty, A.L.; Leddy, H.A.; Liedtke, W.B.; Guilak, F. TRPV4 as a therapeutic target for joint diseases. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2015, 388, 437–450. [Google Scholar] [CrossRef] [Green Version]
- O’Conor, C.J.; Leddy, H.A.; Benefield, H.C.; Liedtke, W.B.; Guilak, F. TRPV4-mediated mechanotransduction regulates the metabolic response of chondrocytes to dynamic loading. Proc. Natl. Acad. Sci. USA 2014, 111, 1316–1321. [Google Scholar] [CrossRef] [Green Version]
- O’Conor, C.J.; Ramalingam, S.; Zelenski, N.A.; Benefield, H.C.; Rigo, I.; Little, D.; Wu, C.-L.; Chen, D.; Liedtke, W.; McNulty, A.L.; et al. Cartilage-specific knockout of the mechanosensory ion channel TRPV4 decreases age-related osteoarthritis. Sci. Rep. 2016, 6, 29053. [Google Scholar]
- Walter, B.; Purmessur, D.; Moon, A.; Occhiogrosso, J.; Laudier, D.; Hecht, A.; Iatridis, J. Reduced tissue osmolarity increases TRPV4 expression and pro-inflammatory cytokines in intervertebral disc cells. Eur. Cell Mater. 2016, 32, 123–136. [Google Scholar] [CrossRef] [PubMed]
- Franco-Obregon, A.; Cambria, E.; Greutert, H.; Wernas, T.; Hitzl, W.; Egli, M.; Sekiguchi, M.; Boos, N.; Hausmann, O.; Ferguson, S.J.; et al. TRPC6 in simulated microgravity of intervertebral disc cells. Eur. Spine J. 2018, 27, 2621–2630. [Google Scholar] [CrossRef] [Green Version]
- Sadowska, A.; Touli, E.; Hitzl, W.; Greutert, H.; Ferguson, S.J.; Wuertz-Kozak, K.; Hausmann, O. Inflammaging in cervical and lumbar degenerated intervertebral discs: Analysis of proinflammatory cytokine and TRP channel expression. Eur. Spine J. 2018, 27, 564–577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadowska, A.; Hitzl, W.; Karol, A.; Jaszczuk, P.; Cherif, H.; Haglund, L.; Hausmann, O.N.; Wuertz, K. Differential regulation of TRP channel gene and protein expression by intervertebral disc degeneration and back pain. Sci. Rep. 2019, 9, 18889. [Google Scholar] [CrossRef] [PubMed]
- Kameda, T.; Zvick, J.; Vuk, M.; Sadowska, A.; Tam, W.K.; Leung, V.Y.-L.; Bölcskei, K.; Helyes, Z.; Applegate, L.A.; Hausmann, O.; et al. Expression and activity of TRPA1 and TRPV1 in the intervertebral disc: Association with inflammation and matrix remodeling. Int. J. Mol. Sci. 2019, 20, 1767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naito, Y.; Hino, K.; Bono, H.; Ui-Tei, K. CRISPRdirect: Software for designing CRISPR/Cas guide RNA with reduced off-target sites. Bioinformatics 2015, 31, 1120–1123. [Google Scholar] [CrossRef]
- Morgens, D.W.; Wainberg, M.; Boyle, E.; Ursu, O.; Araya, C.L.; Tsui, C.K.; Haney, M.S.; Hess, G.T.; Han, K.; Jeng, E.; et al. Genome-scale measurement of off-target activity using Cas9 toxicity in high-throughput screens. Nat. Commun. 2017, 8, 15178. [Google Scholar] [CrossRef]
- Sanjana, N.E.; Shalem, O.; Zhang, F. Improved vectors and genome-wide libraries for CRISPR screening. Nat. Methods 2014, 11, 783–784. [Google Scholar] [CrossRef] [Green Version]
- Stewart, S.A.; Dykxhoorn, D.M.; Palliser, D.; Mizuno, H.; Yu, E.Y.; An, N.S.; Sabatini, D.M.; Chen, I.S.; Hahn, W.C.; Sharp, P.A.; et al. Lentivirus-delivered stable gene silencing by RNAi in primary cells. RNA 2003, 9, 493–501. [Google Scholar] [CrossRef] [Green Version]
- Vincent, T.L. Mechanoflammation in osteoarthritis pathogenesis. Semin. Arthritis Rheum. 2019, 49, S36–S38. [Google Scholar] [CrossRef] [PubMed]
- Krupkova, O.; Cambria, E.; Bešše, L.; Besse, A.; Bowles, R.; Wuertz-Kozak, K. The potential of CRISPR/Cas9 genome editing for the study and treatment of intervertebral disc pathologies. JOR Spine 2018, 1, e1003. [Google Scholar] [CrossRef] [PubMed]
- Stumpo, D.J.; Lai, W.S.; Blackshear, P.J. Inflammation: Cytokines and RNA-based regulation. Wiley Interdiscip. Rev. RNA 2010, 1, 60–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazumder, B.; Li, X.; Barik, S. Translation control: A multifaceted regulator of inflammatory response. J. Immunol. 2010, 184, 3311–3319. [Google Scholar] [CrossRef] [Green Version]
- Nayak, P.S.; Wang, Y.; Najrana, T.; Priolo, L.M.; Rios, M.; Shaw, S.K.; Sanchez-Esteban, J. Mechanotransduction via TRPV4 regulates inflammation and differentiation in fetal mouse distal lung epithelial cells. Respir. Res. 2015, 16, 60. [Google Scholar] [CrossRef] [Green Version]
- Ohsaki, A.; Tanuma, S.I.; Tsukimoto, M. TRPV4 channel-regulated ATP release contributes to gamma-irradiation-induced production of IL-6 and IL-8 in epidermal keratinocytes. Biol. Pharm. Bull. 2018, 41, 1620–1626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, Y.; Pan, Y.; Guo, S.; Sun, L.; Zhang, C.; Wang, L. The roles of mechanosensitive ion channels and associated downstream MAPK signaling pathways in PDLC mechanotransduction. Mol. Med. Rep. 2020, 21, 2113–2122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phan, M.N.; Leddy, H.A.; Votta, B.J.; Kumar, S.; Levy, D.S.; Lipshutz, D.B.; Lee, S.H.; Liedtke, W.; Guilak, F. Functional characterization of TRPV4 as an osmotically sensitive ion channel in porcine articular chondrocytes. Arthritis Rheum. 2009, 60, 3028–3037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qu, Y.-J.; Zhang, X.; Fan, Z.-Z.; Huai, J.; Teng, Y.-B.; Zhang, Y.; Yue, S.-W. Effect of TRPV4-p38 MAPK pathway on neuropathic pain in rats with chronic compression of the dorsal root ganglion. BioMed Res. Int. 2016, 2016, 6978923. [Google Scholar] [CrossRef] [Green Version]
- Pizzute, T.; He, F.; Zhang, X.; Pei, M. Impact of Wnt signals on human intervertebral disc cell regeneration. J. Orthop. Res. 2018, 36, 3196–3207. [Google Scholar] [CrossRef] [Green Version]
- Farhang, N.; Ginley-Hidinger, M.; Berrett, K.C.; Gertz, J.; Lawrence, B.; Bowles, R.D.; Bowles, R.D. Lentiviral CRISPR epigenome editing of inflammatory receptors as a gene therapy strategy for disc degeneration. Hum. Gene Ther. 2019, 30, 1161–1175. [Google Scholar] [CrossRef] [PubMed]
- Thorneloe, K.S.; Cheung, M.; Bao, W.; Alsaid, H.; Lenhard, S.; Jian, M.-Y.; Costell, M.; Maniscalco-Hauk, K.; Krawiec, J.A.; Olzinski, A.; et al. An orally active TRPV4 channel blocker prevents and resolves pulmonary edema induced by heart failure. Sci. Transl. Med. 2012, 4, 159ra148. [Google Scholar] [CrossRef] [PubMed]
- Dong, Q.; Li, J.; Wu, Q.; Zhao, N.; Qian, C.; Ding, D.; Wang, B.-B.; Chen, L.; Guo, K.-F.; Fu, D.; et al. Blockage of transient receptor potential vanilloid 4 alleviates myocardial ischemia/reperfusion injury in mice. Sci. Rep. 2017, 7, 42678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michalick, L.; Erfinanda, L.; Weichelt, U.; Van Der Giet, M.; Liedtke, W.; Kuebler, W.M. Transient receptor potential vanilloid 4 and serum glucocorticoid-regulated kinase 1 are critical mediators of lung injury in overventilated mice in vivo. Anesthesiology 2017, 126, 300–311. [Google Scholar] [CrossRef]
- Goyal, N.; Skrdla, P.; Schroyer, R.; Kumar, S.; Fernando, D.; Oughton, A.; Norton, N.; Sprecher, D.L.; Cheriyan, J. Clinical pharmacokinetics, safety, and tolerability of a novel, first-in-class TRPV4 ion channel inhibitor, GSK2798745, in healthy and heart failure subjects. Am. J. Cardiovasc. Drugs 2019, 19, 335–342. [Google Scholar] [CrossRef] [Green Version]
- Setton, L.A.; Chen, J. Cell mechanics and mechanobiology in the intervertebral disc. Spine 2004, 29, 2710–2723. [Google Scholar] [CrossRef]
- Weber, K.T.; Alipui, D.O.; Sison, C.P.; Bloom, O.; Quraishi, S.; Overby, M.C.; Levine, M.; Chahine, N.O. Serum levels of the proinflammatory cytokine interleukin-6 vary based on diagnoses in individuals with lumbar intervertebral disc diseases. Arthritis Res. 2016, 18, 3. [Google Scholar] [CrossRef] [Green Version]
- Khan, A.N.; Jacobsen, H.E.; Khan, J.; Filippi, C.G.; Levine, M.; Lehman, R.A.; Riew, K.D.; Lenke, L.G.; Chahine, N. Inflammatory biomarkers of low back pain and disc degeneration: A review. Ann. N. Y. Acad. Sci. 2017, 1410, 68–84. [Google Scholar] [CrossRef]
- Wang, K.; Bao, J.-P.; Yang, S.; Hong, X.; Liu, L.; Xie, X.-H.; Wu, X.-T. A cohort study comparing the serum levels of pro- or anti-inflammatory cytokines in patients with lumbar radicular pain and healthy subjects. Eur. Spine J. 2016, 25, 1428–1434. [Google Scholar] [CrossRef]
- Kang, J.D.; Stefanovic-Racic, M.; McIntyre, L.A.; Georgescu, H.I.; Evans, C.H. Toward a biochemical understanding of human intervertebral disc degeneration and herniation. Contributions of nitric oxide, interleukins, prostaglandin E2, and matrix metalloproteinases. Spine 1997, 22, 1065–1073. [Google Scholar] [CrossRef]
- Miyamoto, H.; Saura, R.; Doita, M.; Kurosaka, M.; Mizuno, K. The role of cyclooxygenase-2 in lumbar disc herniation. Spine 2002, 27, 2477–2483. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.D.; Georgescu, H.I.; McIntyre-Larkin, L.; Stefanovic-Racic, M.; Evans, C.H. Herniated cervical intervertebral discs spontaneously produce matrix metalloproteinases, nitric oxide, interleukin-6, and prostaglandin E2. Spine 1995, 20, 2373–2378. [Google Scholar] [CrossRef] [PubMed]
- Wuertz, K.; Vo, N.; Kletsas, D.; Boos, N. Inflammatory and catabolic signalling in intervertebral discs: The roles of NF-kappaB and MAP kinases. Eur. Cell Mater. 2012, 23, 103–119, discussion 119–120. [Google Scholar] [CrossRef] [PubMed]
- Henckaerts, E.; Linden, R.M. Adeno-associated virus: A key to the human genome? Future Virol. 2010, 5, 555–574. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N° | Age | Sex | Diagnosis | Disc Level | Pfirrmann Grade |
|---|---|---|---|---|---|
| 1 | 74 | M | Herniation | L4/5 | 3 |
| 2 | 71 | M | Herniation | L4/5 | 3 |
| 3 | 76 | F | DDD | L5/S1 | 4 |
| 4 | 56 | F | DDD | C6/7 | 3 |
| 5 | 40 | F | Herniation | C5/6 | 2 |
| 6 | 52 | F | Herniation | L4/5 | 3 |
| 7 | 15 | M | DDD | L4/5 | 4 |
| 8 | 75 | F | DDD | L3/4 | 3 |
| 9 | 46 | M | Herniation | L4/5 | 4 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cambria, E.; Arlt, M.J.E.; Wandel, S.; Krupkova, O.; Hitzl, W.; Passini, F.S.; Hausmann, O.N.; Snedeker, J.G.; Ferguson, S.J.; Wuertz-Kozak, K. TRPV4 Inhibition and CRISPR-Cas9 Knockout Reduce Inflammation Induced by Hyperphysiological Stretching in Human Annulus Fibrosus Cells. Cells 2020, 9, 1736. https://doi.org/10.3390/cells9071736
Cambria E, Arlt MJE, Wandel S, Krupkova O, Hitzl W, Passini FS, Hausmann ON, Snedeker JG, Ferguson SJ, Wuertz-Kozak K. TRPV4 Inhibition and CRISPR-Cas9 Knockout Reduce Inflammation Induced by Hyperphysiological Stretching in Human Annulus Fibrosus Cells. Cells. 2020; 9(7):1736. https://doi.org/10.3390/cells9071736
Chicago/Turabian StyleCambria, Elena, Matthias J. E. Arlt, Sandra Wandel, Olga Krupkova, Wolfgang Hitzl, Fabian S. Passini, Oliver N. Hausmann, Jess G. Snedeker, Stephen J. Ferguson, and Karin Wuertz-Kozak. 2020. "TRPV4 Inhibition and CRISPR-Cas9 Knockout Reduce Inflammation Induced by Hyperphysiological Stretching in Human Annulus Fibrosus Cells" Cells 9, no. 7: 1736. https://doi.org/10.3390/cells9071736
APA StyleCambria, E., Arlt, M. J. E., Wandel, S., Krupkova, O., Hitzl, W., Passini, F. S., Hausmann, O. N., Snedeker, J. G., Ferguson, S. J., & Wuertz-Kozak, K. (2020). TRPV4 Inhibition and CRISPR-Cas9 Knockout Reduce Inflammation Induced by Hyperphysiological Stretching in Human Annulus Fibrosus Cells. Cells, 9(7), 1736. https://doi.org/10.3390/cells9071736