Mitochondrial DNA Haplotypes Influence Energy Metabolism across Chicken Transmitochondrial Cybrids


Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells Preparation and Culture Conditions
2.2. Generation of Cybrids
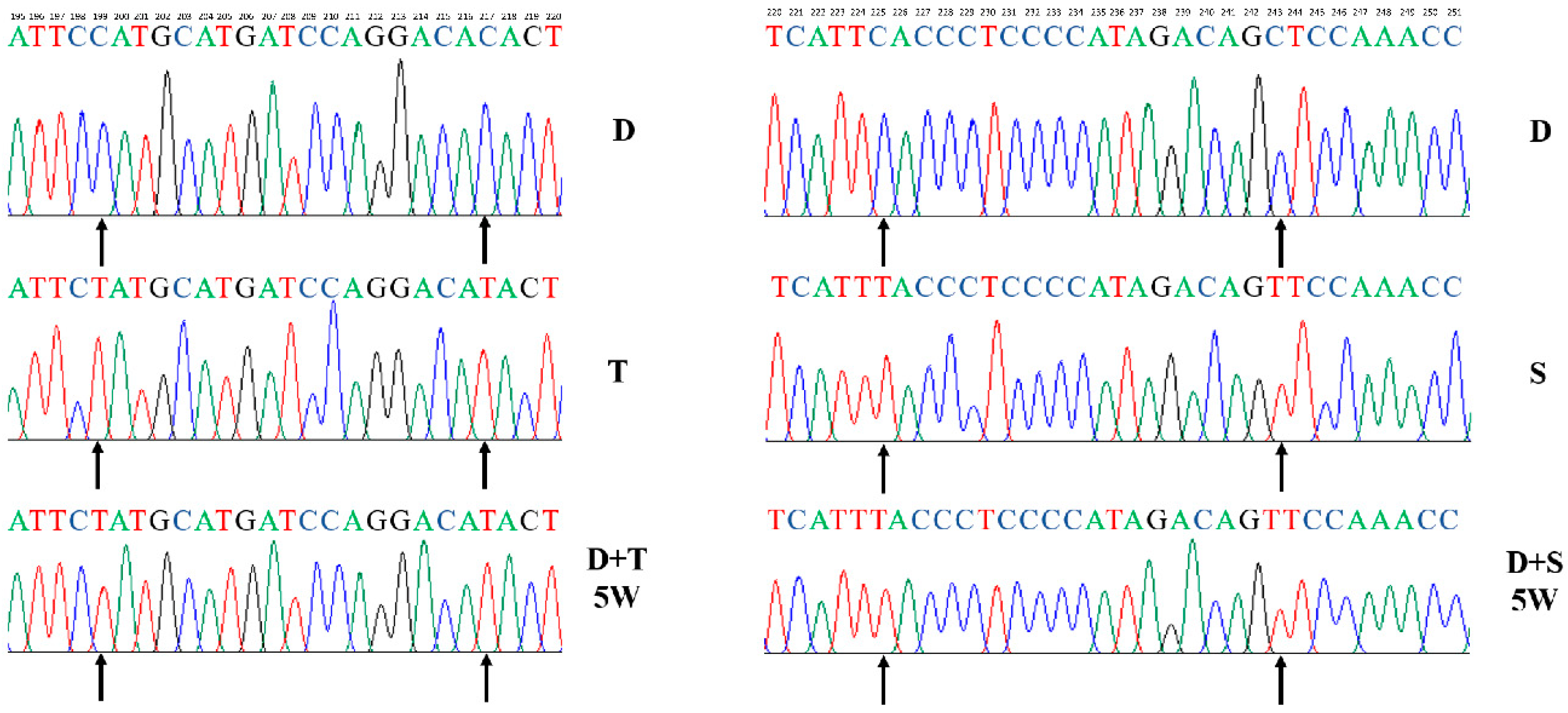
2.3. Mitogenome Sequencing
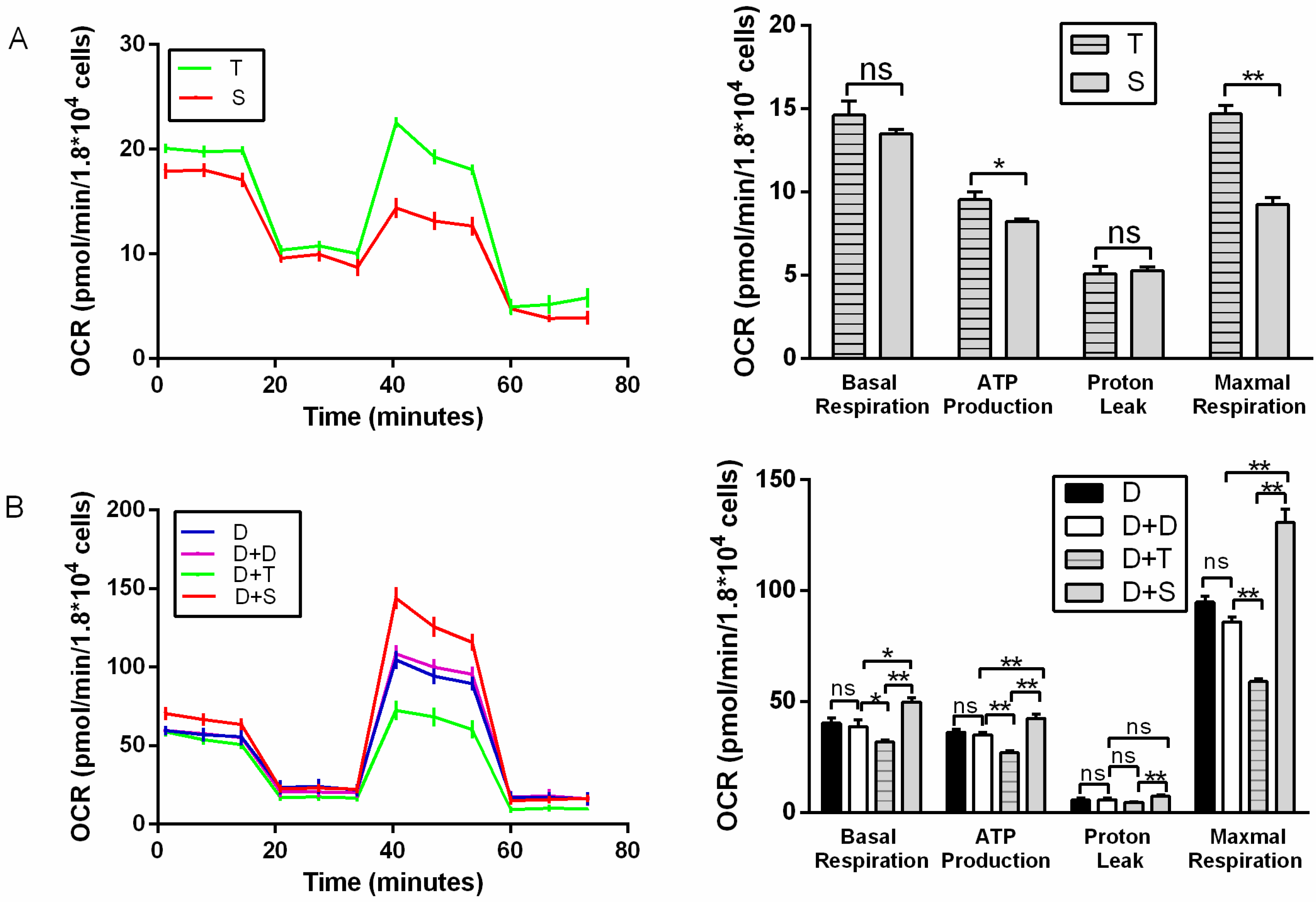
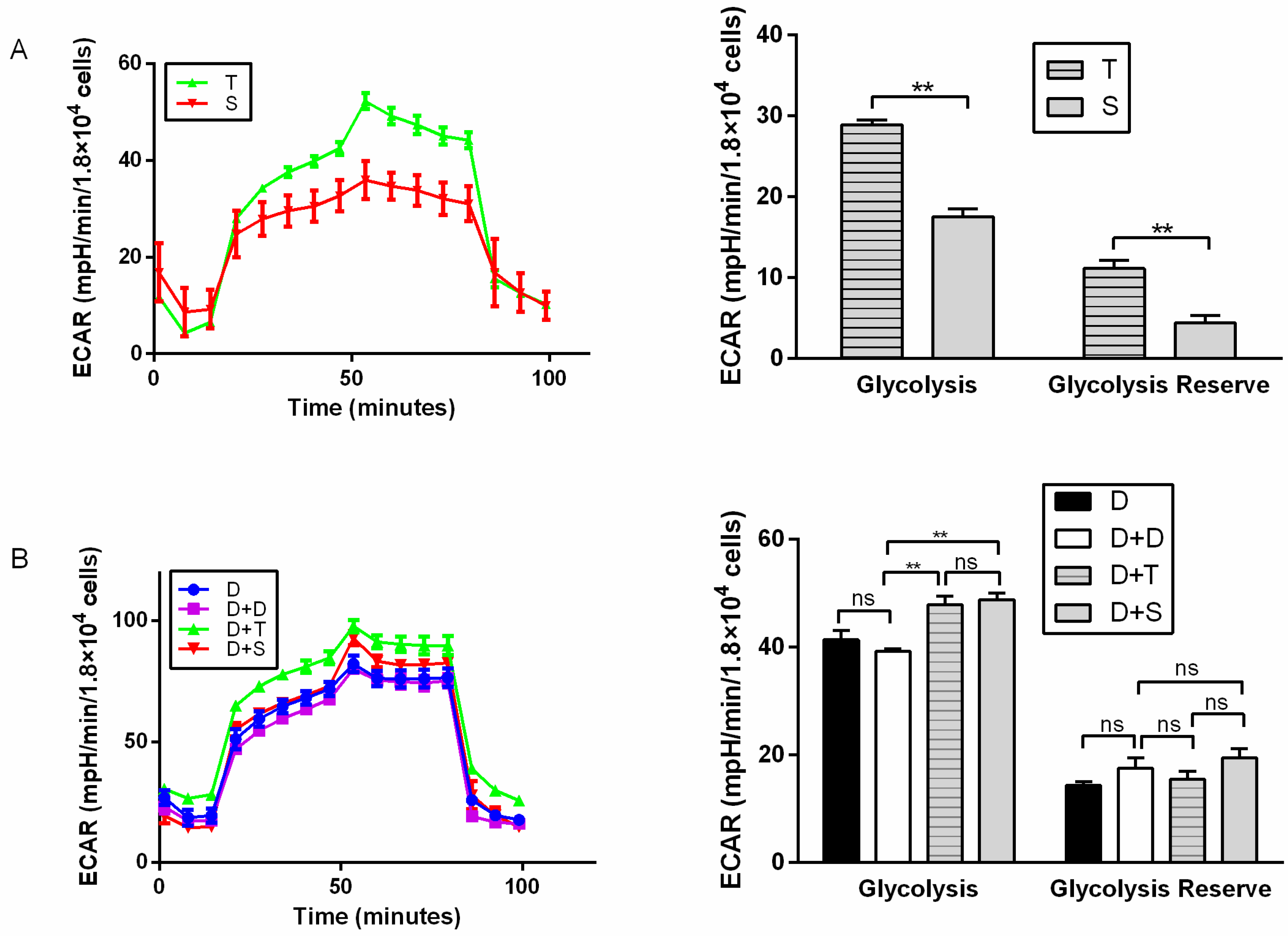
2.4. Metabolic Assays
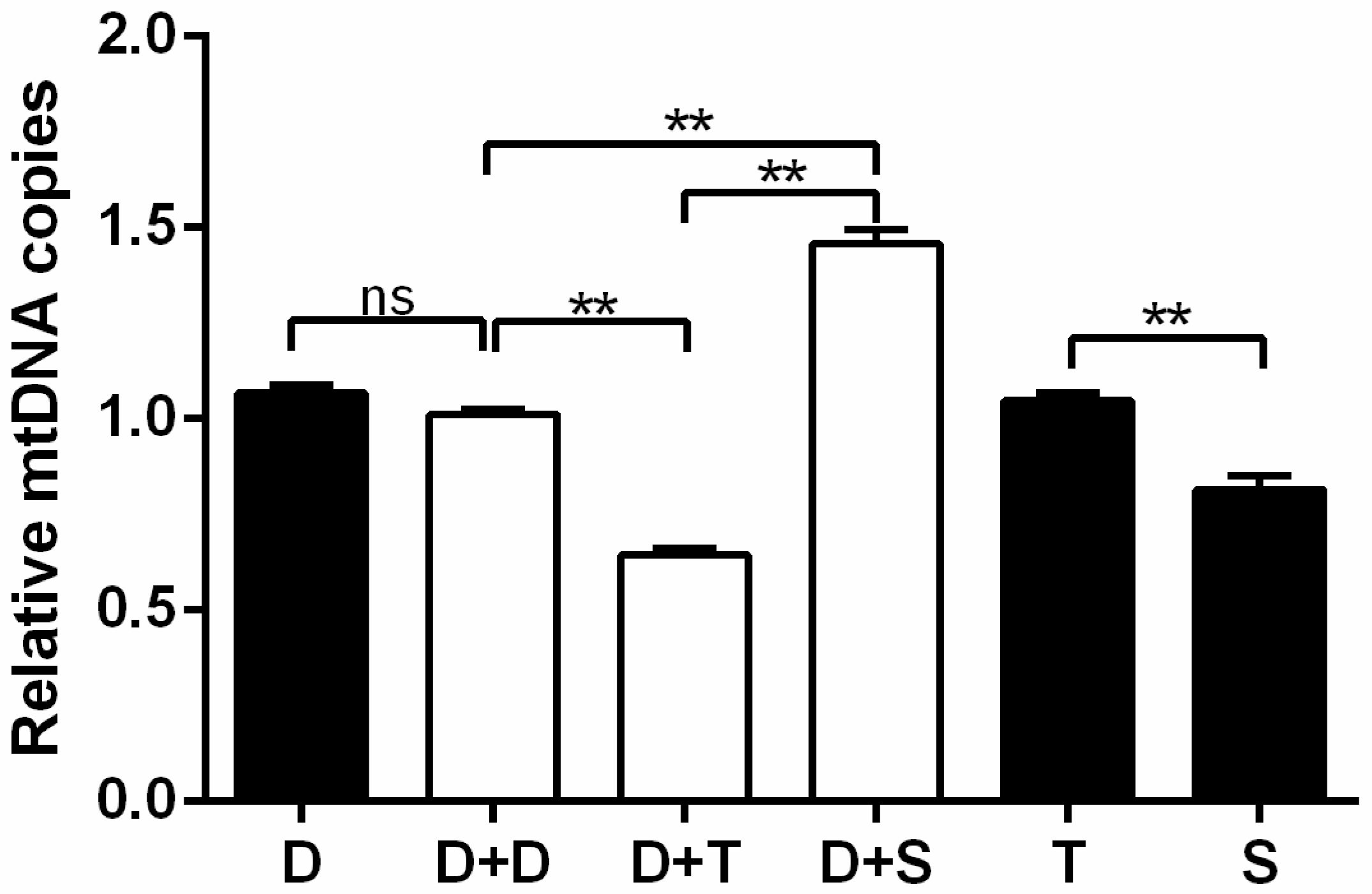
2.5. Total DNA Isolation and Relative mtDNA Copies Detecting
2.6. Total RNA Extraction and qPCR
2.7. Statistical Analysis
3. Results
3.1. Generation of Cybrids
3.2. Mitogenome Sequencing
3.3. Metabolic Assays
3.4. Relative mtDNA Copies Detecting
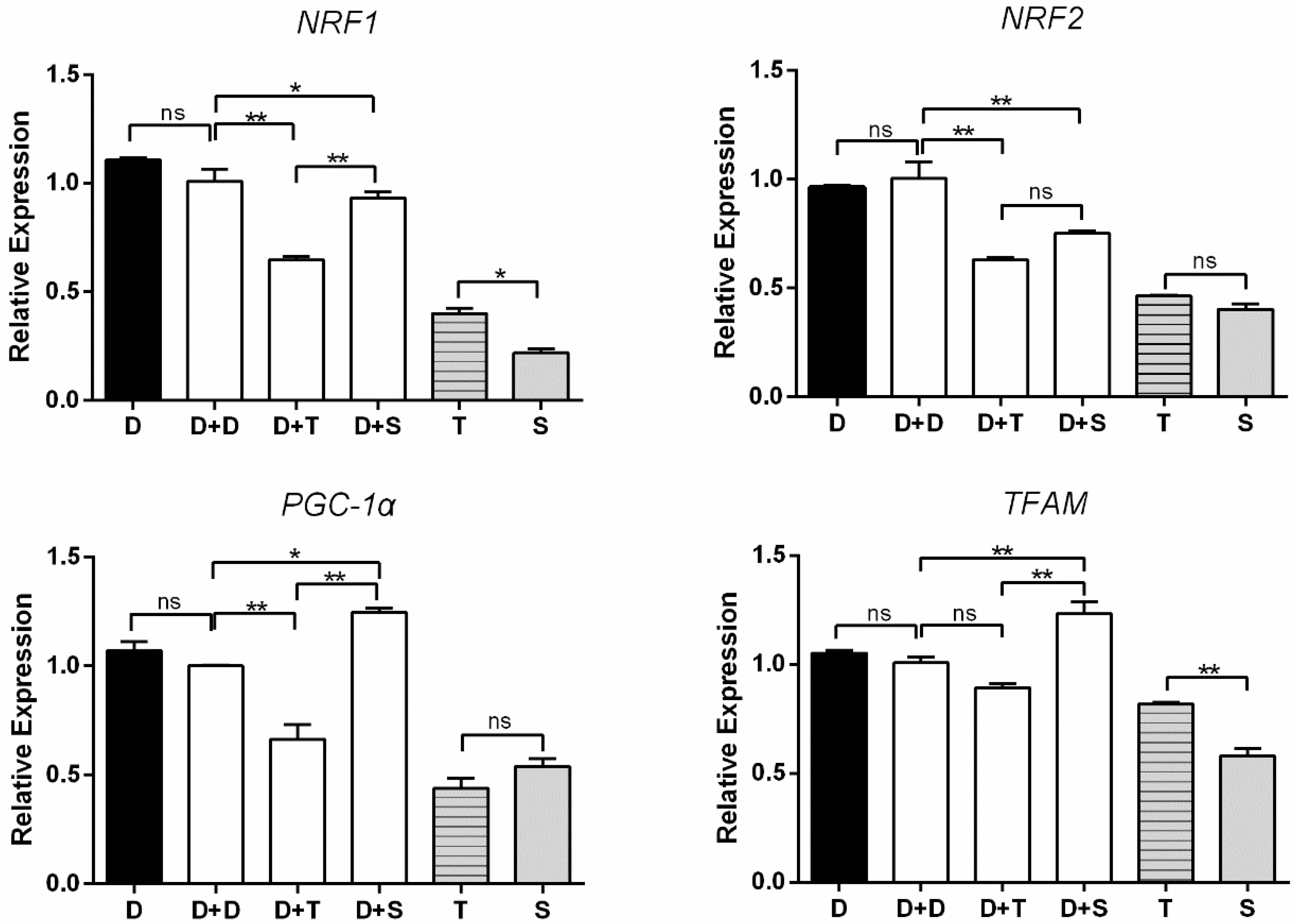
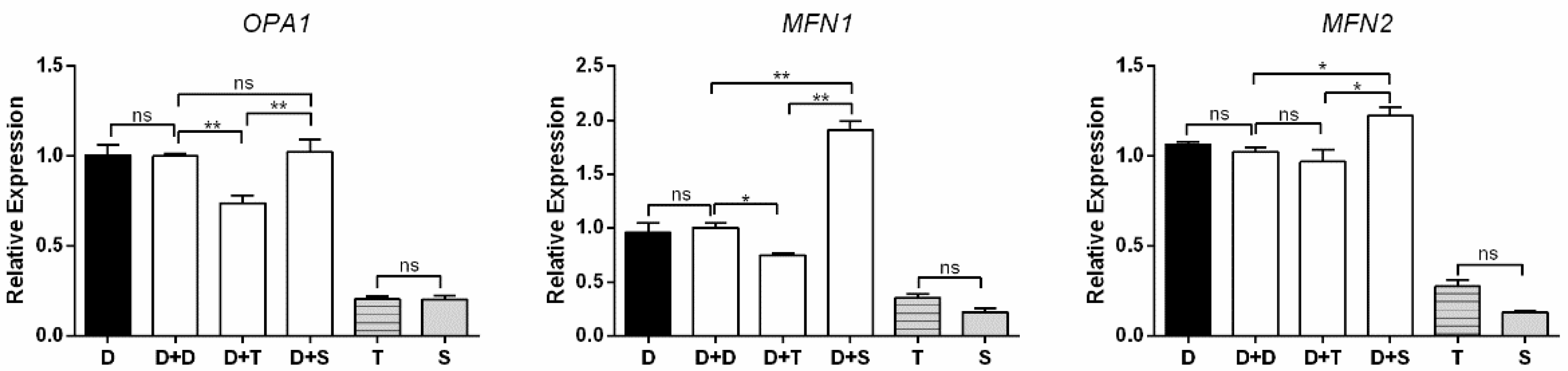
3.5. Expression Levels of Genes Involved in Mito-Biogenesis and Mito-Fusion
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chinnery, P.F.; Hudson, G. Mitochondrial genetics. Brit. Med. Bull. 2013, 106, 135–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Giezen, M.; Tovar, J. Degenerate mitochondria. EMBO Rep. 2005, 6, 525–530. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Pesini, E.; Mishmar, D.; Brandon, M.; Procaccio, V.; Wallace, D.C. Effects of purifying and adaptive selection on regional variation in human mtDNA. Science 2004, 303, 223–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goto, Y.; Nonaka, I.; Horai, S. A mutation in the tRNA (Leu) (UUR) gene associated with the MELAS subgroup of mitochondrial encephalomyopathies. Nature 1990, 348, 651–653. [Google Scholar] [CrossRef] [PubMed]
- Bratic, A.; Larsson, N.G. The role of mitochondria in aging. J. Clin. Invest. 2013, 123, 951–957. [Google Scholar] [CrossRef] [Green Version]
- Fernandez-Moreno, M.; Soto-Hermida, A.; Vazquez-Mosquera, M.E.; Cortes-Pereira, E.; Pertega, S.; Relano, S.; Oreiro-Villar, N.; Fernandez-Lopez, C.; Blanco, F.J.; Rego-Perez, I. A replication study and meta-analysis of mitochondrial DNA variants in the radiographic progression of knee osteoarthritis. Rheumatology 2017, 56, 263–270. [Google Scholar] [CrossRef] [Green Version]
- Schutz, M.M.; Freeman, A.E.; Lindberg, G.L.; Koehler, C.M.; Beitz, D.C. The Effect of Mitochondrial-DNA on Milk-Production and Health of Dairy-Cattle. Livest. Prod. Sci. 1994, 37, 283–295. [Google Scholar] [CrossRef]
- Tsai, T.S.; Rajasekar, S.; St. John, J.C. The relationship between mitochondrial DNA haplotype and the reproductive capacity of domestic pigs (Sus scrofa domesticus). BMC Genet. 2016, 17, 67. [Google Scholar] [CrossRef] [Green Version]
- Lu, W.W.; Hou, L.L.; Zhang, W.W.; Zhang, P.F.; Chen, W.; Kang, X.T.; Huang, Y.Q. Study on heteroplasmic variation and the effect of chicken mitochondrial ND2. Mitochondrial DNA Part A 2016, 27, 2303–2309. [Google Scholar]
- Li, S.; Aggrey, S.E.; Zadworny, D.; Fairfull, W.; Kuhnlein, U. Evidence for a genetic variation in the mitochondrial genome affecting traits in White Leghorn chickens. J. Hered. 1998, 89, 222–226. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.Y.; He, Y.; Li, J.Y.; Bao, H.G.; Wu, C. Association of a missense nucleotide polymorphism in the MT-ND2 gene with mitochondrial reactive oxygen species production in the Tibet chicken embryo incubated in normoxia or simulated hypoxia. Anim. Genet. 2013, 44, 472–475. [Google Scholar] [CrossRef] [PubMed]
- Paliwal, S.; Fiumera, A.C.; Fiumera, H.L. Mitochondrial-Nuclear Epistasis Contributes to Phenotypic Variation and Coadaptation in Natural Isolates of Saccharomyces cerevisiae. Genetics 2014, 198, 1251–1265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- James, A.C.; Ballard, J.W. Mitochondrial genotype affects fitness in Drosophila simulans. Genetics 2003, 164, 187–194. [Google Scholar] [PubMed]
- Latorre-Pellicer, A.; Moreno-Loshuertos, R.; Lechuga-Vieco, A.V.; Sanchez-Cabo, F.; Torroja, C.; Acin-Perez, R.; Calvo, E.; Aix, E.; Gonzalez-Guerra, A.; Logan, A.; et al. Mitochondrial and nuclear DNA matching shapes metabolism and healthy ageing. Nature 2016, 535, 561–565. [Google Scholar] [CrossRef] [PubMed]
- Trounce, I.; Wallace, D.C. Production of transmitochondrial mouse cell lines by cybrid rescue of rhodamine-6G pre-treated L-cells. Somat. Cell Mol. Genet. 1996, 22, 81–85. [Google Scholar] [CrossRef]
- Wang, J.K.; Xiang, H.; Liu, L.Q.; Kong, M.H.; Yin, T.; Zhao, X.B. Mitochondrial haplotypes influence metabolic traits across bovine interand intra-species cybrids. Sci. Rep. 2017, 7, 4179. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.S.; Li, Y.; Peng, M.S.; Zhong, L.; Wang, Z.J.; Li, Q.Y.; Tu, X.L.; Dong, Y.; Zhu, C.L.; Wang, L.; et al. Genomic Analyses Reveal Potential Independent Adaptation to High Altitude in Tibetan Chickens. Mol. Biol. Evol. 2015, 32, 1880–1889. [Google Scholar] [CrossRef]
- Hudson, N.J.; Bottje, W.G.; Hawken, R.J.; Kong, B.; Okimoto, R.; Reverter, A. Mitochondrial metabolism: A driver of energy utilisation and product quality? Anim. Prod. Sci. 2017, 57, 2204–2215. [Google Scholar] [CrossRef]
- Himly, M.; Foster, D.N.; Bottoli, I.; Iacovoni, J.S.; Vogt, P.K. The DF-1 chicken fibroblast cell line: Transformation induced by diverse oncogenes and cell death resulting from infection by avian leukosis viruses. Virology 1998, 248, 295–304. [Google Scholar] [CrossRef] [Green Version]
- Bacman, S.R.; Moraes, C.T. Transmitochondrial technology in animal cells. Methods Cell Biol. 2007, 80, 503–524. [Google Scholar]
- Trounce, I.A.; Kim, Y.L.; Jun, A.S.; Wallace, D.C. Assessment of mitochondrial oxidative phosphorylation in patient muscle biopsies, lymphoblasts, and transmitochondrial cell lines. Methods Enzymol. 1996, 264, 484–509. [Google Scholar] [PubMed]
- Grantham, R. Amino-Acid Difference Formula to Help Explain Protein Evolution. Science 1974, 185, 862–864. [Google Scholar] [CrossRef] [PubMed]
- Li, W.H.; Wu, C.I.; Luo, C.C. Nonrandomness of Point Mutation as Reflected in Nucleotide Substitutions in Pseudogenes and Its Evolutionary Implications. J. Mol. Evol. 1984, 21, 58–71. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, W.; Jiang, L.; Bhasin, S.; Khan, S.M.; Swerdlow, R.H. DNA extraction procedures meaningfully influence qPCR-based mtDNA copy number determination. Mitochondrion 2009, 9, 261–265. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2 (T) (-Delta Delta C) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Fernandez-Moreno, M.; Soto-Hermida, A.; Vazquez-Mosquera, M.E.; Cortes-Pereira, E.; Relano, S.; Hermida-Gomez, T.; Pertega, S.; Oreiro-Villar, N.; Fernandez-Lopez, C.; Garesse, R.; et al. Mitochondrial DNA haplogroups influence the risk of incident knee osteoarthritis in OAI and CHECK cohorts. A meta-analysis and functional study. Ann. Rheum. Dis. 2017, 76, 1114–1122. [Google Scholar] [CrossRef]
- Kenney, M.C.; Chwa, M.; Atilano, S.R.; Falatoonzadeh, P.; Ramirez, C.; Malik, D.; Tarek, M.; Caceres-del-Carpio, J.; Nesburn, A.B.; Boyer, D.S.; et al. Inherited mitochondrial DNA variants can affect complement, inflammation and apoptosis pathways: Insights into mitochondrial-nuclear interactions. Hum. Mol. Genet. 2014, 23, 3537–3551. [Google Scholar] [CrossRef] [Green Version]
- Song, M.; Han, W.; Bao, H.; Liu, C.; Wu, C.; Zhao, C. Physiological Adaptability of Tibet Chicken Embryo Liver to Hypoxia and its Protein Profile Analysis. Asian J. Anim. Vet. Adv. 2010, 5, 547–556. [Google Scholar] [CrossRef] [Green Version]
- Schon, E.A. Mitochondrial genetics and disease. Trends Biochem. Sci. 2000, 25, 555–560. [Google Scholar] [CrossRef]
- Burton, R.S.; Pereira, R.J.; Barreto, F.S. Cytonuclear Genomic Interactions and Hybrid Breakdown. Annu. Rev. Ecol. Evol. Syst. 2013, 44, 281–302. [Google Scholar] [CrossRef]
- Chou, J.Y.; Leu, J.Y. The Red Queen in mitochondria: Cyto-nuclear co-evolution, hybrid breakdown and human disease. Front. Genet. 2015, 6, 187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gan, L.; Yan, J.; Liu, Z.J.; Feng, M.; Sun, C. Adiponectin Prevents Reduction of Lipid-Induced Mitochondrial Biogenesis via AMPK/ACC2 Pathway in Chicken Adipocyte. J. Cell Biochem. 2015, 116, 1090–1100. [Google Scholar] [CrossRef]
- Reznick, R.M.; Shulman, G.I. The role of AMP-activated protein kinase in mitochondrial biogenesis. J. Physiol. 2006, 574, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Cogliati, S.; Frezza, C.; Soriano, M.E.; Varanita, T.; Quintana-Cabrera, R.; Corrado, M.; Cipolat, S.; Costa, V.; Casarin, A.; Gomes, L.C.; et al. Mitochondrial cristae shape determines respiratory chain supercomplexes assembly and respiratory efficiency. Cell 2013, 155, 160–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, P.; Carelli, V.; Manfredi, G.; Chan, D.C. Proteolytic cleavage of Opa1 stimulates mitochondrial inner membrane fusion and couples fusion to oxidative phosphorylation. Cell Metab. 2014, 19, 630–641. [Google Scholar] [CrossRef] [Green Version]
- Bellizzi, D.; D’Aquila, P.; Giordano, M.; Montesanto, A.; Passarino, G. Global DNA methylation levels are modulated by mitochondrial DNA variants. Epigenomics 2012, 4, 17–27. [Google Scholar] [CrossRef] [Green Version]
- Havird, J.C.; Sloan, D.B. The Roles of Mutation, Selection, and Expression in Determining Relative Rates of Evolution in Mitochondrial versus Nuclear Genomes. Mol. Biol. Evol. 2016, 33, 3042–3053. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Comparison | SNPs/InDels | AA Change | Grantham Score Summary | ||||
|---|---|---|---|---|---|---|---|
| D-loop | rRNA | tRNA | Protein | Total | |||
| D/T | 9 | 2 | 1 | 26 | 39 | 5 | 1 in 0–50, 4 in 51–100 |
| D/S | 10 | 4 | 2 | 29 | 46 | 4 | 2 in 0–50, 2 in 51–100 |
| T/S | 11 | 4 | 1 | 25 | 41 | 5 | 1 in 0–50, 4 in 51–100 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kong, M.; Xiang, H.; Wang, J.; Liu, J.; Zhang, X.; Zhao, X. Mitochondrial DNA Haplotypes Influence Energy Metabolism across Chicken Transmitochondrial Cybrids. Genes 2020, 11, 100. https://doi.org/10.3390/genes11010100
Kong M, Xiang H, Wang J, Liu J, Zhang X, Zhao X. Mitochondrial DNA Haplotypes Influence Energy Metabolism across Chicken Transmitochondrial Cybrids. Genes. 2020; 11(1):100. https://doi.org/10.3390/genes11010100
Chicago/Turabian StyleKong, Minghua, Hai Xiang, Jikun Wang, Jian Liu, Xiben Zhang, and Xingbo Zhao. 2020. "Mitochondrial DNA Haplotypes Influence Energy Metabolism across Chicken Transmitochondrial Cybrids" Genes 11, no. 1: 100. https://doi.org/10.3390/genes11010100
APA StyleKong, M., Xiang, H., Wang, J., Liu, J., Zhang, X., & Zhao, X. (2020). Mitochondrial DNA Haplotypes Influence Energy Metabolism across Chicken Transmitochondrial Cybrids. Genes, 11(1), 100. https://doi.org/10.3390/genes11010100