A COL7A1 Variant in a Litter of Neonatal Basset Hounds with Dystrophic Epidermolysis Bullosa

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Animal Selection
2.3. Necropsy, Histopathology, and Electron Microscopy
2.4. DNA Extraction
2.5. Whole Genome Sequencing
2.6. Variant Calling
2.7. Sanger Sequencing and Genotyping
3. Results
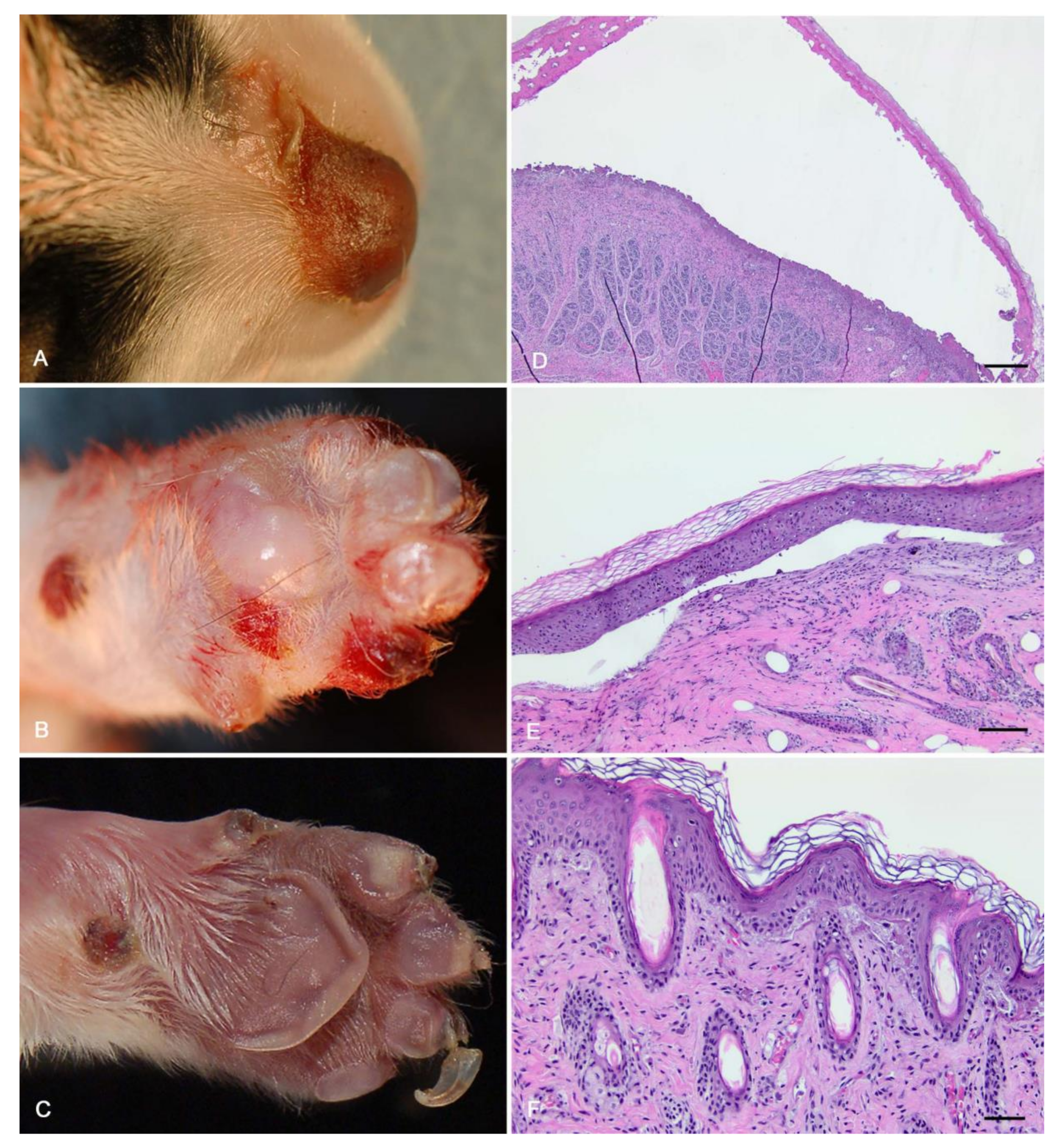
3.1. Clinical History and Necropsy Findings
3.2. Histologic Findings
3.3. Electron Microscopy
3.4. Genetic Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Has, C.; Bauer, J.; Bodemer, C.; Bolling, M.; Bruckner-Tuderman, L.; Diem, A.; Fine, J.-D.; Heagerty, A.; Hovnanian, A.; Marinkovich, M.; et al. Consensus reclassification of inherited epidermolysis bullosa and other disorders with skin fragility. Br. J. Dermatol. 2020, 183, 416–627. [Google Scholar] [CrossRef] [Green Version]
- Fine, J.-D.; Bruckner-Tuderman, L.; Eady, R.A.J.; Bauer, E.A.; Bauer, J.W.; Has, C.; Heagerty, A.; Hintner, H.; Hovnanian, A.; Jonkman, M.F.; et al. Inherited epidermolysis bullosa: Updated recommendations on diagnosis and classification. J. Am. Acad. Dermatol. 2014, 70, 1103–1126. [Google Scholar] [CrossRef]
- Vahidnezhad, H.; Youssefian, L.; Saeidian, A.H.; Touati, A.; Pajouhanfar, S.; Baghdadi, T.; Shadmehri, A.A.; Giunta, C.; Kraenzlin, M.; Syx, D.; et al. Mutations in PLOD3, encoding lysyl hydroxylase 3, cause a complex connective tissue disorder including recessive dystrophic epidermolysis bullosa-like blistering phenotype with abnormal anchoring fibrils and type VII collagen deficiency. Matrix Biol. 2019, 81, 91–106. [Google Scholar] [CrossRef] [Green Version]
- Has, C.; Liu, L.; Bolling, M.; Charlesworth, A.; El Hachem, M.; Escámez, M.; Fuentes, I.; Büchel, S.; Hiremagalore, R.; Pohla-Gubo, G.; et al. Clinical practice guidelines for laboratory diagnosis of epidermolysis bullosa. Br. J. Dermatol. 2019, 182, 574–592. [Google Scholar] [CrossRef]
- DEB Register—The International Database of Dystrophic Epidermolysis Bullosa Patients and COL7A1 Mutations. Available online: https://www.deb-central.org/ (accessed on 2 October 2020).
- Akker, P.C.V.D.; Jonkman, M.F.; Rengaw, T.; Bruckner-Tuderman, L.; Has, C.; Bauer, J.W.; Klausegger, A.; Zambruno, G.; Castiglia, D.; Mellerio, J.E.; et al. The international dystrophic epidermolysis bullosa patient registry: An online database of dystrophic epidermolysis bullosa patients and their COL7A1 mutations. Hum. Mutat. 2011, 32, 1100–1107. [Google Scholar] [CrossRef]
- Wertheim-Tysarowska, K.; Sobczyńska-Tomaszewska, A.; Kowalewski, C.; Skroński, M.; Święćkowski, G.; Kutkowska-Kaźmierczak, A.; Woźniak, K.; Bal, J. The COL7A1 mutation database. Hum. Mutat. 2011, 33, 327–331. [Google Scholar] [CrossRef]
- Varki, R.; Sadowski, S.; Uitto, J.; Pfendner, E. Epidermolysis bullosa. II. Type VII collagen mutations and phenotype-genotype correlations in the dystrophic subtypes. J. Med. Genet. 2006, 44, 181–192. [Google Scholar] [CrossRef] [Green Version]
- Mauldin, E.A.; Wang, P.; Olivry, T.; Henthorn, P.S.; Casal, M.L. Epidermolysis bullosa simplex in sibling Eurasier dogs is caused by aPLECnon-sense variant. Veter. Dermatol. 2017, 28, 10-e3. [Google Scholar] [CrossRef] [Green Version]
- Olivry, T.; Linder, K.E.; Wang, P.; Bizikova, P.; Bernstein, J.A.; Dunston, S.M.; Paps, J.S.; Casal, M.L. Deficient Plakophilin-1 Expression Due to a Mutation in PKP1 Causes Ectodermal Dysplasia-Skin Fragility Syndrome in Chesapeake Bay Retriever Dogs. PLoS ONE 2012, 7, e32072. [Google Scholar] [CrossRef] [Green Version]
- Nagata, M.; Iwasaki, T.; Masuda, H.; Shimizu, H. Non-lethal junctional epidermolysis bullosa in a dog. Br. J. Dermatol. 1997, 137, 445–449. [Google Scholar] [CrossRef]
- Dunstan, R.W.; Sills, R.C.; Wilkinson, J.; Paller, A.S.; Hashimoto, K.H. A disease resembling junctional epidermolysis bullosa in a toy poodle. Am. J. Dermatopathol. 1988, 10, 442–447. [Google Scholar] [PubMed]
- Olivry, T.; Poujade-Delverdier, A.; Dunston, S.M.; Fine, J.-D.; Ortonne, J.-P. Absent expression of collagen XVII (BPAG2, BP180) in canine familial localized junctional epidermolysis bullosa. Veter. Dermatol. 1997, 8, 203–212. [Google Scholar] [CrossRef]
- Niskanen, J.E.; Dillard, K.; Arumilli, M.; Salmela, E.; Anttila, M.; Lohi, H.; Hytönen, M.K. Nonsense variant in COL7A1 causes recessive dystrophic epidermolysis bullosa in Central Asian Shepherd dogs. PLoS ONE 2017, 12, e0177527. [Google Scholar] [CrossRef]
- Palazzi, X.; Marchal, T.; Chabanne, L.; Spadafora, A.; Meneguzzi, G.; Magnol, J.-P. Inherited Dystrophic Epidermolysis Bullosa in Inbred Dogs: A Spontaneous Animal Model for Somatic Gene Therapy. J. Investig. Dermatol. 2000, 115, 135–137. [Google Scholar] [CrossRef] [Green Version]
- Baldeschi, C.; Gache, Y.; Rattenholl, A.; Bouille, P.; Danos, O.; Ortonne, J.-P.; Bruckner-Tuderman, L.; Meneguzzi, G. Genetic correction of canine dystrophic epidermolysis bullosa mediated by retroviral vectors. Hum. Mol. Genet. 2003, 12, 1897–1905. [Google Scholar] [CrossRef] [Green Version]
- Nagata, M.; Shimizu, H.; Masunaga, T.; Nishikawa, T.; Nanko, H.; Kariya, K.; Washizu, T.; Ishida, T. Dystrophic form of inherited epidermolysis bullosa in a dog (Akita Inu). Br. J. Dermatol. 1995, 133, 1000–1003. [Google Scholar] [CrossRef]
- Kiener, S.; Laprais, A.; Mauldin, E.A.; Jagannathan, V.; Olivry, T.; Leeb, T. LAMB3 Missense Variant in Australian Shepherd Dogs with Junctional Epidermolysis Bullosa. Genes 2020, 11, 1055. [Google Scholar] [CrossRef]
- Jagannathan, V.; Drögemüller, C.; Leeb, T. Dog Biomedical Variant Database Consortium (DBVDC). A comprehensive biomedical variant catalogue based on whole genome sequences of 582 dogs and eight wolves. Anim. Genet. 2019, 50, 695–704. [Google Scholar] [CrossRef] [Green Version]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.B.; Daly, M.J.; et al. The Genome Analysis Toolkit: A MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cingolani, P.; Platts, A.; Wang, L.; Coon, M.; Nguyen, T.; Wang, L.; Land, S.J.; Lu, X.; Ruden, D.M. A program for annotating and predicting the effects of single nucleotide polymorphisms, SnpEff: SNPs in the genome of Drosophila melanogaster strain w1118; iso-2; iso-3. Fly 2012, 6, 80–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGrath, J.; Ashton, G.H.; Mellerio, J.E.; McMillan, J.R.; Eady, R.A.; Salas-Alanis, J.C.; Swensson, O. Moderation of Phenotypic Severity in Dystrophic and Junctional Forms of Epidermolysis Bullosa Through In-Frame Skipping of Exons Containing Non-Sense or Frameshift Mutations. J. Investig. Dermatol. 1999, 113, 314–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, H.J.; Uitto, J. Type VII Collagen: The Anchoring Fibril Protein at Fault in Dystrophic Epidermolysis Bullosa. Dermatol. Clin. 2010, 28, 93–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eady, R.A.J.; Dopping-Hepenstal, P.J. Transmission Electron Microscopy for the Diagnosis of Epidermolysis Bullosa. Dermatol. Clin. 2010, 28, 211–222. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Filtering Step | Homozygous Variants | Heterozygous Variants |
|---|---|---|
| All variants | 3,198,983 | 2,768,869 |
| Private variants | 18,820 | 80,057 |
| Protein-changing private variants | 49 | 174 |
| Protein-changing private variants in 37 candidate genes | 3 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garcia, T.M.; Kiener, S.; Jagannathan, V.; Russell, D.S.; Leeb, T. A COL7A1 Variant in a Litter of Neonatal Basset Hounds with Dystrophic Epidermolysis Bullosa. Genes 2020, 11, 1458. https://doi.org/10.3390/genes11121458
Garcia TM, Kiener S, Jagannathan V, Russell DS, Leeb T. A COL7A1 Variant in a Litter of Neonatal Basset Hounds with Dystrophic Epidermolysis Bullosa. Genes. 2020; 11(12):1458. https://doi.org/10.3390/genes11121458
Chicago/Turabian StyleGarcia, Teresa Maria, Sarah Kiener, Vidhya Jagannathan, Duncan S. Russell, and Tosso Leeb. 2020. "A COL7A1 Variant in a Litter of Neonatal Basset Hounds with Dystrophic Epidermolysis Bullosa" Genes 11, no. 12: 1458. https://doi.org/10.3390/genes11121458
APA StyleGarcia, T. M., Kiener, S., Jagannathan, V., Russell, D. S., & Leeb, T. (2020). A COL7A1 Variant in a Litter of Neonatal Basset Hounds with Dystrophic Epidermolysis Bullosa. Genes, 11(12), 1458. https://doi.org/10.3390/genes11121458