Altered Expression of GABAergic Markers in the Forebrain of Young and Adult Engrailed-2 Knockout Mice

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Morphometric Analysis
2.3. In Situ Hybridization
2.4. Quantitative RT-PCR
2.5. Immunohistochemistry
2.6. Cell Counts
2.7. Statistical Analyses
3. Results
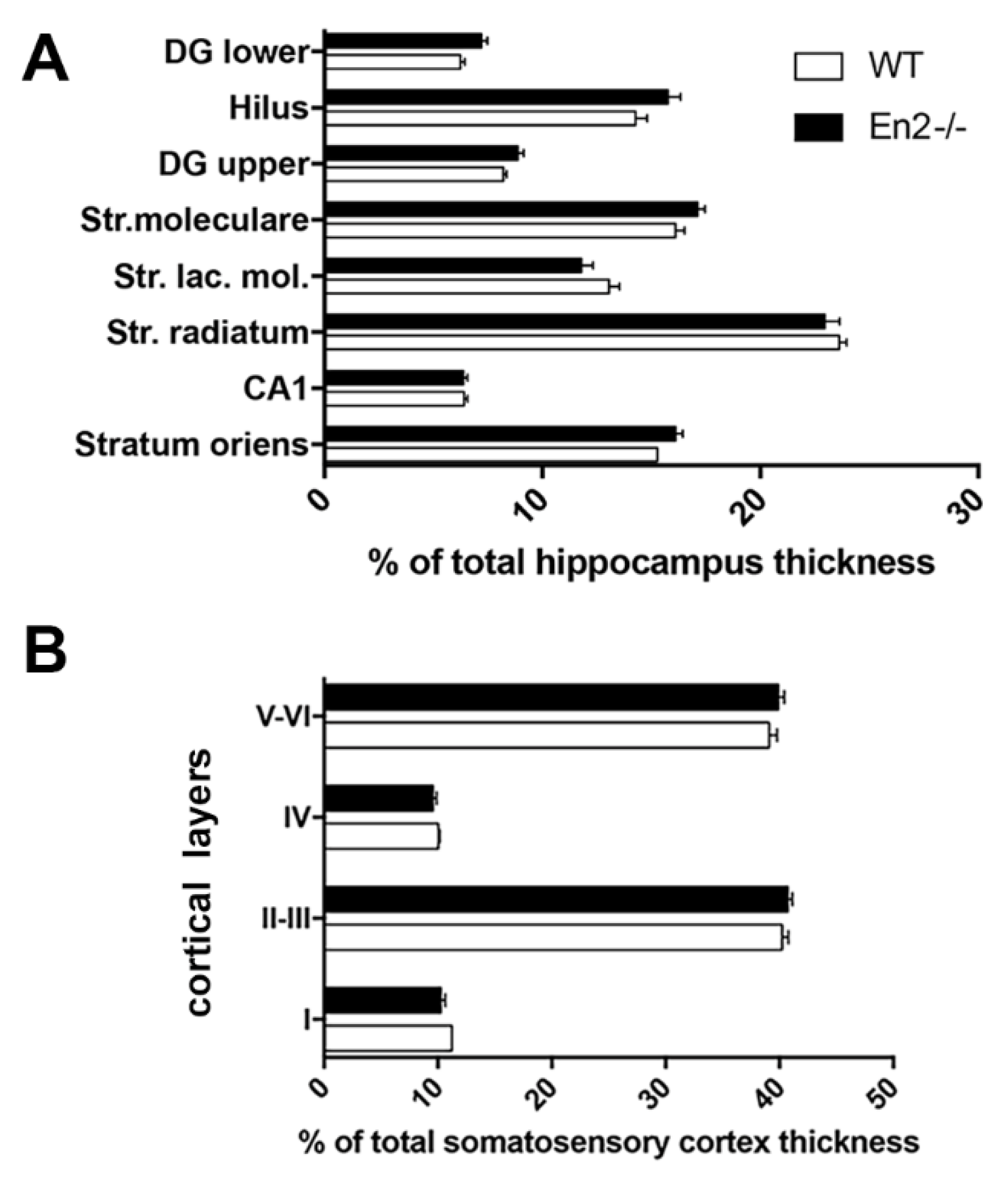
3.1. Normal Layering of Hippocampus and Somatosensory Cortex in Young En2 Mutant Mice
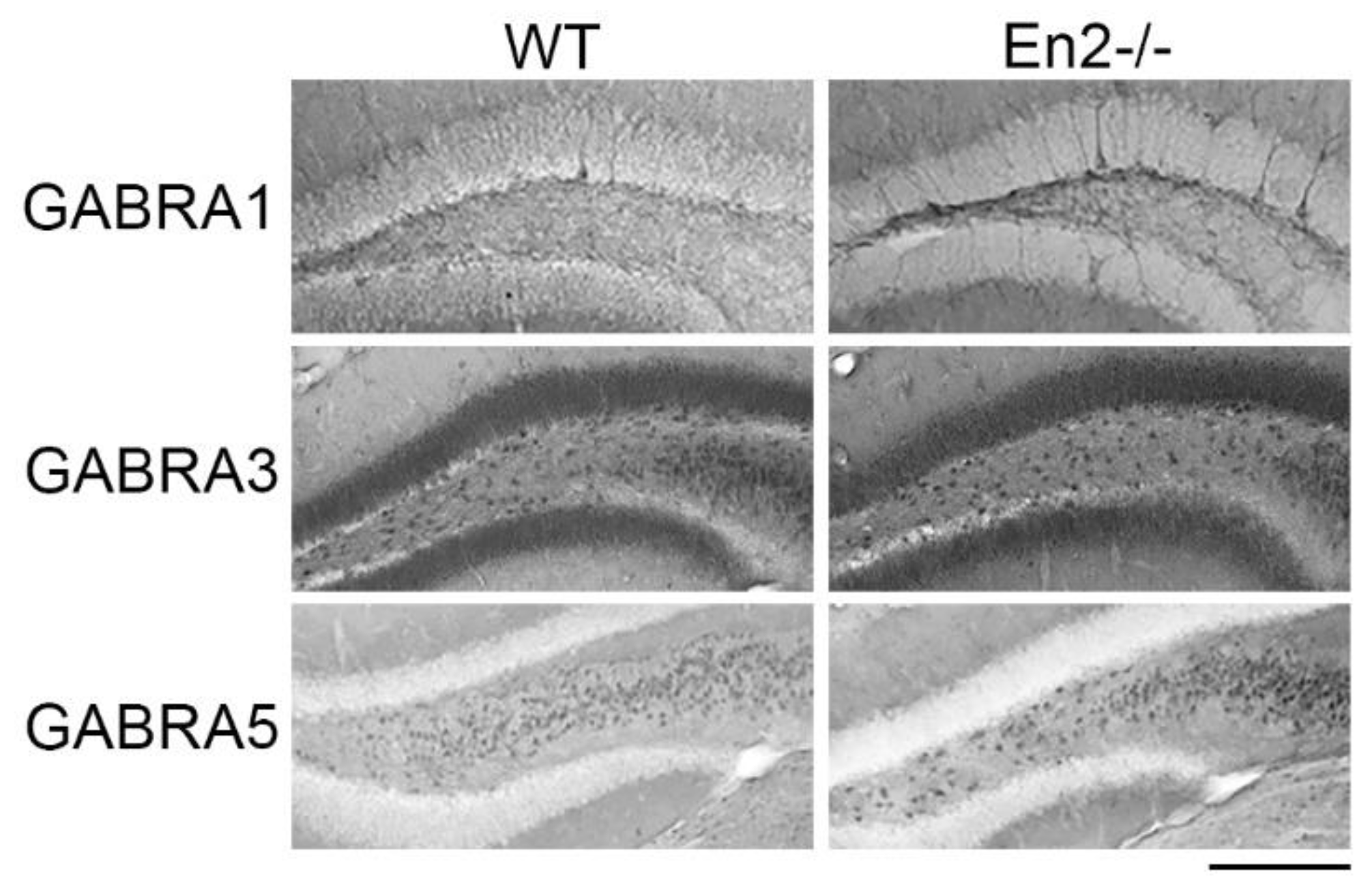
3.2. Altered Expression of GABA Receptor Subunits in Young and Adult En2 Mutant Mice
4. Discussion
4.1. Brief Summary of Results
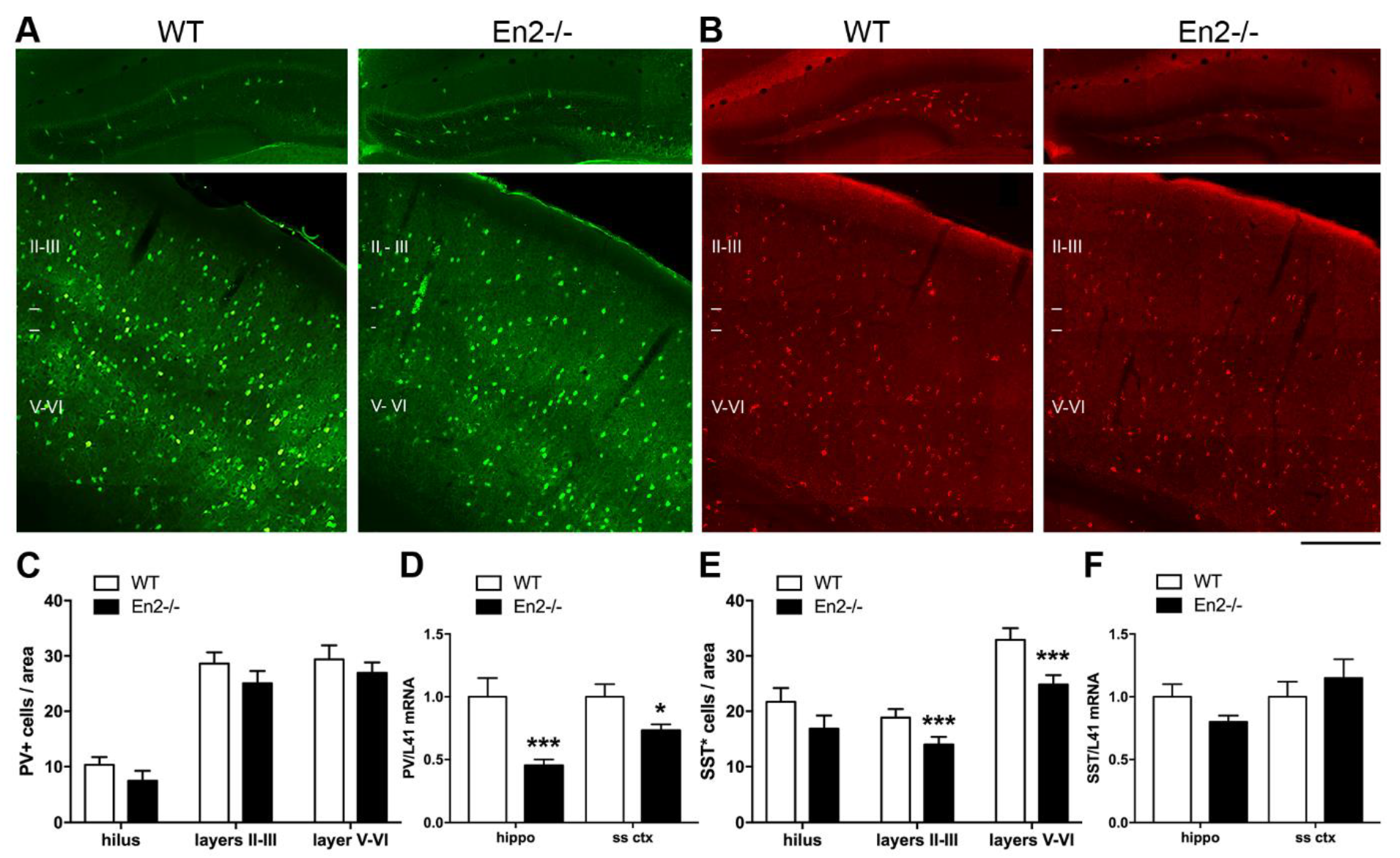
4.2. Altered Expression of GABAergic Interneuron Markers in Young En2 Mutant Mice
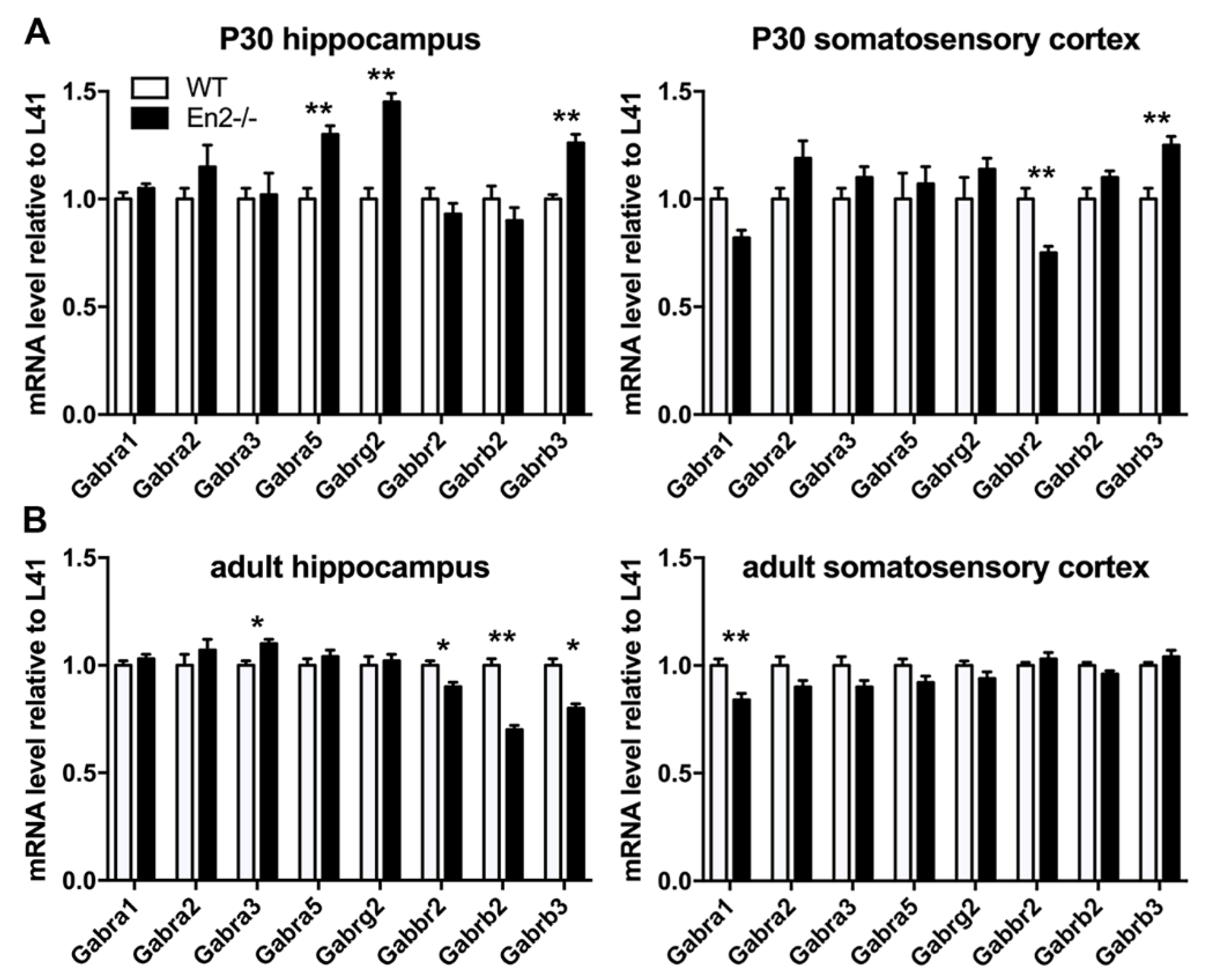
4.3. Altered Expression of GABA Receptor Subunits in Young and Adult En2 Mutant Mice
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rubenstein, J.L.R.; Merzenich, M.M. Model of autism: Increased ratio of excitation/inhibition in key neural systems. Genes Brain Behav. 2003, 2, 255–267. [Google Scholar] [CrossRef] [PubMed]
- Wonders, C.P.; Anderson, S.A. The origin and specification of cortical interneurons. Nat. Rev. Neurosci. 2006, 7, 687–696. [Google Scholar] [CrossRef] [PubMed]
- Ascoli, G.A.; Alonso-Nanclares, L.; Anderson, S.A.; Barrionuevo, G.; Benavides-Piccione, R.; Burkhalter, A.; Buzsáki, G.; Cauli, B.; Defelipe, J.; Fairén, A.; et al. Petilla terminology: Nomenclature of features of GABAergic interneurons of the cerebral cortex. Nat. Rev. Neurosci. 2008, 9, 557–568. [Google Scholar]
- Kepecs, A.; Fishell, G. Interneuron cell types are fit to function. Nature 2014, 505, 318–326. [Google Scholar] [CrossRef] [Green Version]
- Tremblay, R.; Lee, S.; Rudy, B. GABAergic interneurons in the neocortex: From cellular properties to circuits. Neuron 2016, 91, 260–292. [Google Scholar] [CrossRef] [Green Version]
- Pelkey, K.A.; Chittajallu, R.; Craig, M.T.; Tricoire, L.; Wester, J.C.; McBain, C.J. Hippocampal GABAergic inhibitory interneurons. Physiol. Rev. 2017, 97, 1619–1747. [Google Scholar] [CrossRef]
- Huang, Z.J.; Paul, A. The diversity of GABAergic neurons and neural communication elements. Nat. Rev. Neurosci. 2019, 20, 563–572. [Google Scholar] [CrossRef]
- Rossignol, E. Genetics and function of neocortical GABAergic interneurons in neurodevelopmental disorders. Neural. Plast. 2011, 2011, 649325. [Google Scholar] [CrossRef] [Green Version]
- Marín, O. Interneuron dysfunction in psychiatric disorders. Nat. Rev. Neurosci. 2012, 13, 107–120. [Google Scholar] [CrossRef]
- Bozzi, Y.; Provenzano, G.; Casarosa, S. Neurobiological bases of autism-epilepsy comorbidity: A focus on excitation/inhibition imbalance. Eur. J. Neurosci. 2018, 47, 534–548. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, Y.A.; Kemper, T.L.; Bauman, M.L.; Blatt, G.J. Parvalbumin-, calbindin-, and calretinin-immunoreactive hippocampal interneuron density in autism. Acta Neurol. Scand. 2010, 121, 99–108. [Google Scholar] [CrossRef]
- Blatt, G.J.; Fatemi, S.H. Alterations in GABAergic biomarkers in the autism brain: Research findings and clinical implications. Anat. Rec. 2011, 294, 1646–1652. [Google Scholar] [CrossRef] [Green Version]
- Benayed, R.; Choi, J.; Matteson, P.G.; Gharani, N.; Kamdar, S.; Brzustowicz, L.M.; Millonig, J.H. Autism-associated haplotype affects the regulation of the homeobox gene, ENGRAILED 2. Biol. Psychiatry 2009, 66, 911–917. [Google Scholar] [CrossRef] [Green Version]
- Joyner, A.L.; Herrup, K.; Auerbach, B.A.; Davis, C.A.; Rossant, J. Subtle cerebellar phenotype in mice homozygous for a targeted deletion of the En-2 homeobox. Science 1991, 251, 1239–1243. [Google Scholar] [CrossRef]
- DiCicco-Bloom, E.; Lord, C.; Zwaigenbaum, L.; Courchesne, E.; Dager, S.R.; Schmitz, C.; Schultz, R.T.; Crawley, J.; Young, L.J. The developmental neurobiology of autism spectrum disorder. J. Neurosci. 2006, 26, 6897–6906. [Google Scholar] [CrossRef] [Green Version]
- Cheh, M.A.; Millonig, J.H.; Roselli, L.M.; Ming, X.; Jacobsen, E.; Kamdar, S.; Wagner, G.C. En2 knockout mice display neurobehavioral and neurochemical alterations relevant to autism spectrum disorder. Brain Res. 2006, 1116, 166–176. [Google Scholar] [CrossRef]
- Brielmaier, J.; Matteson, P.G.; Silverman, J.L.; Senerth, J.M.; Kelly, S.; Genestine, M.; Millonig, J.H.; Dicicco-Bloom, E.; Crawley, J.N. Autism-relevant social abnormalities and cognitive deficits in engrailed-2 knockout mice. PLoS ONE 2012, 7, e40914. [Google Scholar] [CrossRef] [Green Version]
- Provenzano, G.; Pangrazzi, L.; Poli, A.; Pernigo, M.; Sgadò, P.; Genovesi, S.; Zunino, G.; Berardi, N.; Casarosa, S.; Bozzi, Y. Hippocampal dysregulation of neurofibromin-dependent pathways is associated with impaired spatial learning in engrailed 2 knock-out mice. J. Neurosci. 2014, 34, 13281–13288. [Google Scholar] [CrossRef] [Green Version]
- Chelini, G.; Zerbi, V.; Cimino, L.; Grigoli, A.; Markicevic, M.; Libera, F.; Robbiati, S.; Gadler, M.; Bronzoni, S.; Miorelli, S.; et al. Aberrant somatosensory processing and connectivity in mice lacking Engrailed-2. J. Neurosci. 2019, 39, 1525–1538. [Google Scholar] [CrossRef] [Green Version]
- Tripathi, P.P.; Sgadò, P.; Scali, M.; Viaggi, C.; Casarosa, S.; Simon, H.H.; Vaglini, F.; Corsini, G.U.; Bozzi, Y. Increased susceptibility to kainic acid-induced seizures in Engrailed-2 knockout mice. Neuroscience 2009, 159, 842–849. [Google Scholar] [CrossRef]
- Sgadò, P.; Provenzano, G.; Dassi, E.; Adami, V.; Zunino, G.; Genovesi, S.; Casarosa, S.; Bozzi, Y. Transcriptome profiling in engrailed-2 mutant mice reveals common molecular pathways associated with autism spectrum disorders. Mol. Autism. 2013, 4, 51. [Google Scholar] [CrossRef] [Green Version]
- Sgadò, P.; Genovesi, S.; Kalinovsky, A.; Zunino, G.; Macchi, F.; Allegra, M.; Murenu, E.; Provenzano, G.; Tripathi, P.P.; Casarosa, S.; et al. Loss of GABAergic neurons in the hippocampus and cerebral cortex of Engrailed-2 null mutant mice: Implications for autism spectrum disorders. Exp. Neurol. 2013, 247, 496–505. [Google Scholar] [CrossRef] [Green Version]
- Allegra, M.; Genovesi, S.; Maggia, M.; Cenni, M.C.; Zunino, G.; Sgadò, P.; Caleo, M.; Bozzi, Y. Altered GABAergic markers, increased binocularity and reduced plasticity in the visual cortex of Engrailed-2 knockout mice. Front. Cell. Neurosci. 2014, 8, 163. [Google Scholar] [CrossRef] [Green Version]
- Provenzano, G.; Corradi, Z.; Monsorno, K.; Fedrizzi, T.; Ricceri, L.; Scattoni, M.L.; Bozzi, Y. Comparative gene expression analysis of two mouse models of autism: Transcriptome profiling of the BTBR and En2 (-/-) Hippocampus. Front. Neurosci. 2016, 10, 396. [Google Scholar] [CrossRef]
- Cellot, G.; Cherubini, E. GABAergic signaling as therapeutic target for autism spectrum disorders. Front Pediatr 2014, 2, 70. [Google Scholar] [CrossRef] [Green Version]
- Braat, S.; Kooy, R.F. The GABAA Receptor as a therapeutic target for neurodevelopmental disorders. Neuron 2015, 86, 1119–1130. [Google Scholar] [CrossRef] [Green Version]
- Fritschy, J.M.; Brünig, I. Formation and plasticity of GABAergic synapses: Physiological mechanisms and pathophysiological implications. Pharmacol. Ther. 2003, 98, 299–323. [Google Scholar] [CrossRef]
- Olsen, R.W.; Sieghart, W. International Union of Pharmacology, LXX. Subtypes of gamma-aminobutyric acid(A) receptors: Classification on the basis of subunit composition, pharmacology, and function. Update. Pharmacol. Rev. 2008, 60, 243–260. [Google Scholar] [CrossRef] [Green Version]
- Gassmann, M.; Bettler, B. Regulation of neuronal GABA(B) receptor functions by subunit composition. Nat. Rev. Neurosci. 2012, 13, 380–394. [Google Scholar] [CrossRef]
- Provenzano, G.; Sgadò, P.; Genovesi, S.; Zunino, G.; Casarosa, S.; Bozzi, Y. Hippocampal dysregulation of FMRP/mGluR5 signaling in engrailed-2 knockout mice: A model of autism spectrum disorders. Neuroreport 2015, 26, 1101–1105. [Google Scholar] [CrossRef]
- Boschian, C.; Messina, A.; Bozza, A.; Castellini, M.E.; Provenzano, G.; Bozzi, Y.; Casarosa, S. Impaired neuronal differentiation of neural stem cells lacking the Engrailed-2 gene. Neuroscience 2018, 386, 137–149. [Google Scholar] [CrossRef] [PubMed]
- Filice, F.; Vörckel, K.J.; Sungur, A.Ö.; Wöhr, M.; Schwaller, B. Reduction in parvalbumin expression not loss of the parvalbumin-expressing GABA interneuron subpopulation in genetic parvalbumin and shank mouse models of autism. Mol. Brain 2016, 9, 10. [Google Scholar] [CrossRef] [Green Version]
- Lauber, E.; Filice, F.; Schwaller, B. Prenatal valproate exposure differentially affects parvalbumin-expressing neurons and related circuits in the cortex and striatum of mice. Front. Mol. Neurosci. 2016, 9, 150. [Google Scholar] [CrossRef] [Green Version]
- Lauber, E.; Filice, F.; Schwaller, B. Dysregulation of parvalbumin expression in the cntnap2-/- mouse model of autism spectrum disorder. Front. Mol. Neurosci. 2018, 11, 262. [Google Scholar] [CrossRef] [Green Version]
- Lauber, E.; Filice, F.; Schwaller, B. Parvalbumin neurons as a hub in autism spectrum disorders. J. Neurosci. Res. 2018, 96, 360–361. [Google Scholar] [CrossRef] [Green Version]
- Fatemi, S.H.; Reutiman, T.J.; Folsom, T.D.; Thuras, P.D. GABA(A) receptor downregulation in brains of subjects with autism. J. Autism. Dev. Disord. 2009, 39, 223–230. [Google Scholar] [CrossRef] [Green Version]
- Fatemi, S.H.; Reutiman, T.J.; Folsom, T.D.; Rooney, R.J.; Patel, D.H.; Thuras, P.D. mRNA and protein levels for GABAA alpha4, alpha5, beta1 and GABABR1 receptors are altered in brains from subjects with autism. J. Autism. Dev. Disord. 2010, 40, 743–750. [Google Scholar] [CrossRef] [Green Version]
- Fatemi, S.H.; Folsom, T.D.; Kneeland, R.E.; Liesch, S.B. Metabotropic glutamate receptor 5 upregulation in children with autism is associated with underexpression of both Fragile X mental retardation protein and GABAA receptor beta 3 in adults with autism. Anat. Rec. 2011, 294, 1635–1645. [Google Scholar] [CrossRef] [Green Version]
- Oblak, A.; Gibbs, T.T.; Blatt, G.J. Decreased GABAA receptors and benzodiazepine binding sites in the anterior cingulate cortex in autism. Autism. Res. 2009, 2, 205–219. [Google Scholar] [CrossRef] [Green Version]
- Oblak, A.; Gibbs, T.T.; Blatt, G.J. Reduced GABAA receptors and benzodiazepine binding sites in the posterior cingulate cortex and fusiform gyrus in autism. Brain Res. 2011, 1380, 218–228. [Google Scholar] [CrossRef] [Green Version]
- McCauley, J.L.; Olson, L.M.; Delahanty, R.; Amin, T.; Nurmi, E.L.; Organ, E.L.; Jacobs, M.M.; Folstein, S.E.; Haines, J.L.; Sutcliffe, J.S. A linkage disequilibrium map of the 1-Mb 15q12 GABA(A) receptor subunit cluster and association to autism. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2004, 131B, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Delahanty, R.J.; Kang, J.Q.; Brune, C.W.; Kistner, E.O.; Courchesne, E.; Cox, N.J.; Cook, E.H., Jr.; Macdonald, R.L.; Sutcliffe, J.S. Maternal transmission of a rare GABRB3 signal peptide variant is associated with autism. Mol. Psychiatry 2011, 16, 86–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendez, M.A.; Horder, J.; Myers, J.; Coghlan, S.; Stokes, P.; Erritzoe, D.; Howes, O.; Lingford-Hughes, A.; Murphy, D.; Nutt, D. The brain GABA-benzodiazepine receptor alpha-5 subtype in autism spectrum disorder: A pilot [(11)C]Ro15-4513 positron emission tomography study. Neuropharmacology 2013, 68, 195–201. [Google Scholar] [CrossRef] [Green Version]
- Horder, J.; Andersson, M.; Mendez, M.A.; Singh, N.; Tangen, Ä.; Lundberg, J.; Gee, A.; Halldin, C.; Veronese, M.; Bölte, S.; et al. GABA(A) receptor availability is not altered in adults with autism spectrum disorder or in mouse models. Sci. Transl. Med. 2018, 10, eaam8434. [Google Scholar] [CrossRef] [Green Version]
- DeLorey, T.M.; Sahbaie, P.; Hashemi, E.; Homanics, G.E.; Clark, J.D. Gabrb3 gene deficient mice exhibit impaired social and exploratory behaviors, deficits in non-selective attention and hypoplasia of cerebellar vermal lobules: A potential model of autism spectrum disorder. Behav. Brain Res. 2008, 187, 207–220. [Google Scholar] [CrossRef] [Green Version]
- Orefice, L.L.; Zimmerman, A.L.; Chirila, A.M.; Sleboda, S.J.; Head, J.P.; Ginty, D.D. Peripheral mechanosensory neuron dysfunction underlies tactile and behavioral deficits in mouse models of ASDs. Cell 2016, 166, 299–313. [Google Scholar] [CrossRef] [Green Version]
- Ellegood, J.; Anagnostou, E.; Babineau, B.A.; Crawley, J.N.; Lin, L.; Genestine, M.; DiCicco-Bloom, E.; Lai, J.K.; Foster, J.A.; Peñagarikano, O.; et al. Clustering autism: Using neuroanatomical differences in 26 mouse models to gain insight into the heterogeneity. Mol. Psychiatry 2015, 20, 118–125. [Google Scholar] [CrossRef]
- Zerbi, V.; Ielacqua, G.D.; Markicevic, M.; Haberl, M.G.; Ellisman, M.H.; A-Bhaskaran, A.; Frick, A.; Rudin, M.; Wenderoth, N. Dysfunctional autism risk genes cause circuit-specific connectivity deficits with distinct developmental trajectories. Cereb. Cortex 2018, 28, 2495–2506. [Google Scholar] [CrossRef] [Green Version]
- Adusei, D.C.; Pacey, L.K.; Chen, D.; Hampson, D.R. Early developmental alterations in gabaergic protein expression in Fragile X knockout mice. Neuropharmacology 2010, 59, 167–171. [Google Scholar] [CrossRef] [Green Version]
- D’Hulst, C.; De Geest, N.; Reeve, S.P.; Van Dam, D.; De Deyn, P.P.; Hassan, B.A.; Kooy, R.F. Decreased expression of the GABAA receptor in fragile X syndrome. Brain Res. 2006, 1121, 238–245. [Google Scholar] [CrossRef]
- Chau, D.K.; Choi, A.Y.; Yang, W.; Leung, W.N.; Chan, C.W. Downregulation of glutamatergic and GABAergic proteins in valproic acid associated social impairment during adolescence in mice. Behav. Brain Res. 2017, 316, 255–260. [Google Scholar] [CrossRef]
- Van der Aa, N.; Kooy, R.F. GABAergic abnormalities in the fragile X syndrome. Eur. J. Paediatr. Neurol. 2020, 24, 100–104. [Google Scholar] [CrossRef]
- Leonzino, M.; Busnelli, M.; Antonucci, F.; Verderio, C.; Mazzanti, M.; Chini, B. The timing of the excitatory-to-inhibitory GABA switch is regulated by the oxytocin receptor via KCC2. Cell Rep. 2016, 15, 96–103. [Google Scholar] [CrossRef] [Green Version]
- Modi, B.; Pimpinella, D.; Pazienti, A.; Zacchi, P.; Cherubini, E.; Griguoli, M. Possible implication of the CA2 hippocampal circuit in social cognition deficits observed in the neuroligin 3 knock-out mouse, a non-syndromic animal model of autism. Front. Psychiatry 2019, 10, 513. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Genbank # | Forward Primer (5′–3′) | Reverse Primer (5′–3′) |
|---|---|---|---|
| L41 | NM_001031808.2 | GGTTCTCCCTTTCTCCCTTG | GCACCCCGACTCTTAGTGAA |
| PV | NM_013645 | TGCTCATCCAAGTTGCAGG | GCCACTTTTGTCTTTGTCCAG |
| SOM | NM_009215 | AGGACGAGATGAGGCTGG | CAGGAGTTAAGGAAGAGATATGGG |
| Gabra1 | NM_010250 | CTCTCCCACACTTTTCTCCC | CCGACAGTGTGCTCAGAATG |
| Gabra2 | NM_008066.4 | AGATTCAAAGCCACTGGAGG | CCAGCACCAACCTGACTG |
| Gabra3 | NM_001357816.1 | CAGACTGAGATAGGGACTAGGAG | AGACAGCAACTTGAAGAGACC |
| Gabra5 | NM_176942.4 | CCCTATCCCAACACCTGAAC | AATGTTCAAAGGGTTCTGCC |
| Gabrg2 | NM_008073.4 | CACCGGGCATGAATGTG | GGATGGTACACGCAGAGATG |
| Gabrb2 | NM_001347314 | TCAGAGGATGACTTTGCTA | GCACACAATAATGTTTACTAT |
| Gabrb3 | NM_008071.3 | GAGGTCTTCACAAGCTCAAAATC | AGGCAGGGTAATATTTCACTCAG |
| Gabbr2 | NM_031802.1 | ACATGCAAAGACCCCATAGAG | TCGTGAGAGTAAGACCGTCG |
| Interneuron Marker | Hippocampus | Somatosensory Cortex | |||
|---|---|---|---|---|---|
| mRNA | Cell Counts (hilus) | mRNA | Cell Counts (layers II–III) | Cell Counts (layers V–VI) | |
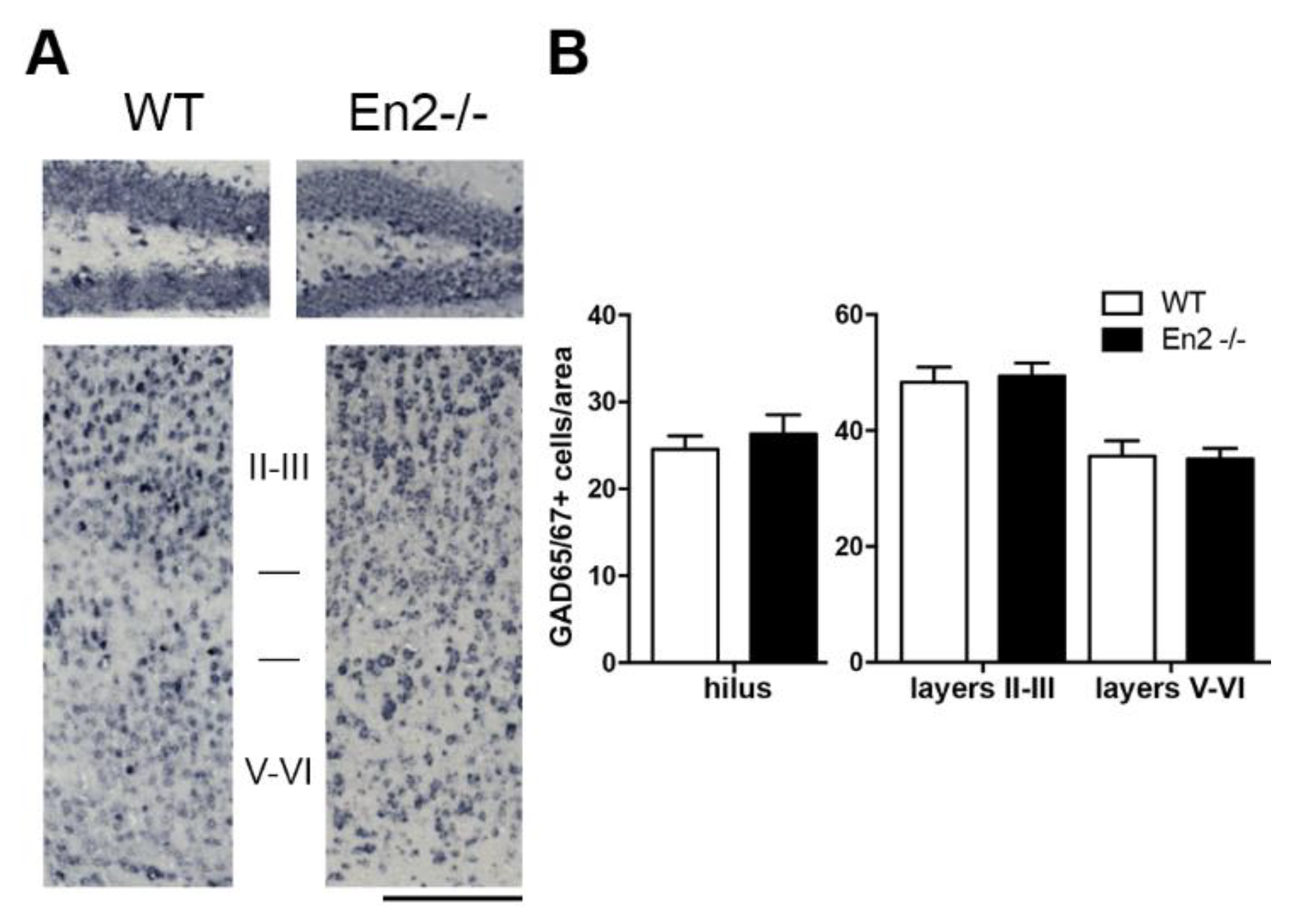
| GAD | not tested | no difference | not tested | no difference | no difference |
| PV | −55% | no difference | −27% | no difference | no difference |
| SST | no difference | no difference | no difference | −25% | −25% |
| Receptor Subunit | P30 | Adult | ||
|---|---|---|---|---|
| Hippocampus | Somatosensory Cortex | Hippocampus | Somatosensory Cortex) | |
| Gabra1 | no difference | no difference | no difference | −17% |
| Gabra2 | no difference | no difference | no difference | no difference |
| Gabra3 | no difference | no difference | +10% | no difference |
| Gabra5 | +30% | no difference | no difference | no difference |
| Gabrb2 | no difference | no difference | −33% | no difference |
| Gabrb3 | +30% | +30% | −20% | no difference |
| Gabrg2 | +43% | no difference | no difference | no difference |
| Gabbr2 | no difference | −26% | −10% | no difference |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Provenzano, G.; Gilardoni, A.; Maggia, M.; Pernigo, M.; Sgadò, P.; Casarosa, S.; Bozzi, Y. Altered Expression of GABAergic Markers in the Forebrain of Young and Adult Engrailed-2 Knockout Mice. Genes 2020, 11, 384. https://doi.org/10.3390/genes11040384
Provenzano G, Gilardoni A, Maggia M, Pernigo M, Sgadò P, Casarosa S, Bozzi Y. Altered Expression of GABAergic Markers in the Forebrain of Young and Adult Engrailed-2 Knockout Mice. Genes. 2020; 11(4):384. https://doi.org/10.3390/genes11040384
Chicago/Turabian StyleProvenzano, Giovanni, Angela Gilardoni, Marika Maggia, Mattia Pernigo, Paola Sgadò, Simona Casarosa, and Yuri Bozzi. 2020. "Altered Expression of GABAergic Markers in the Forebrain of Young and Adult Engrailed-2 Knockout Mice" Genes 11, no. 4: 384. https://doi.org/10.3390/genes11040384
APA StyleProvenzano, G., Gilardoni, A., Maggia, M., Pernigo, M., Sgadò, P., Casarosa, S., & Bozzi, Y. (2020). Altered Expression of GABAergic Markers in the Forebrain of Young and Adult Engrailed-2 Knockout Mice. Genes, 11(4), 384. https://doi.org/10.3390/genes11040384