Characterizing Y-STRs in the Evaluation of Population Differentiation Using the Mean of Allele Frequency Difference between Populations

Abstract
:1. Introduction
2. Materials and Methods
2.1. Data-Set Collection
2.2. Calculation of the Mean of Allele Frequency Difference
2.3. The Evaluation of Population Differentiation
3. Results and Discussion
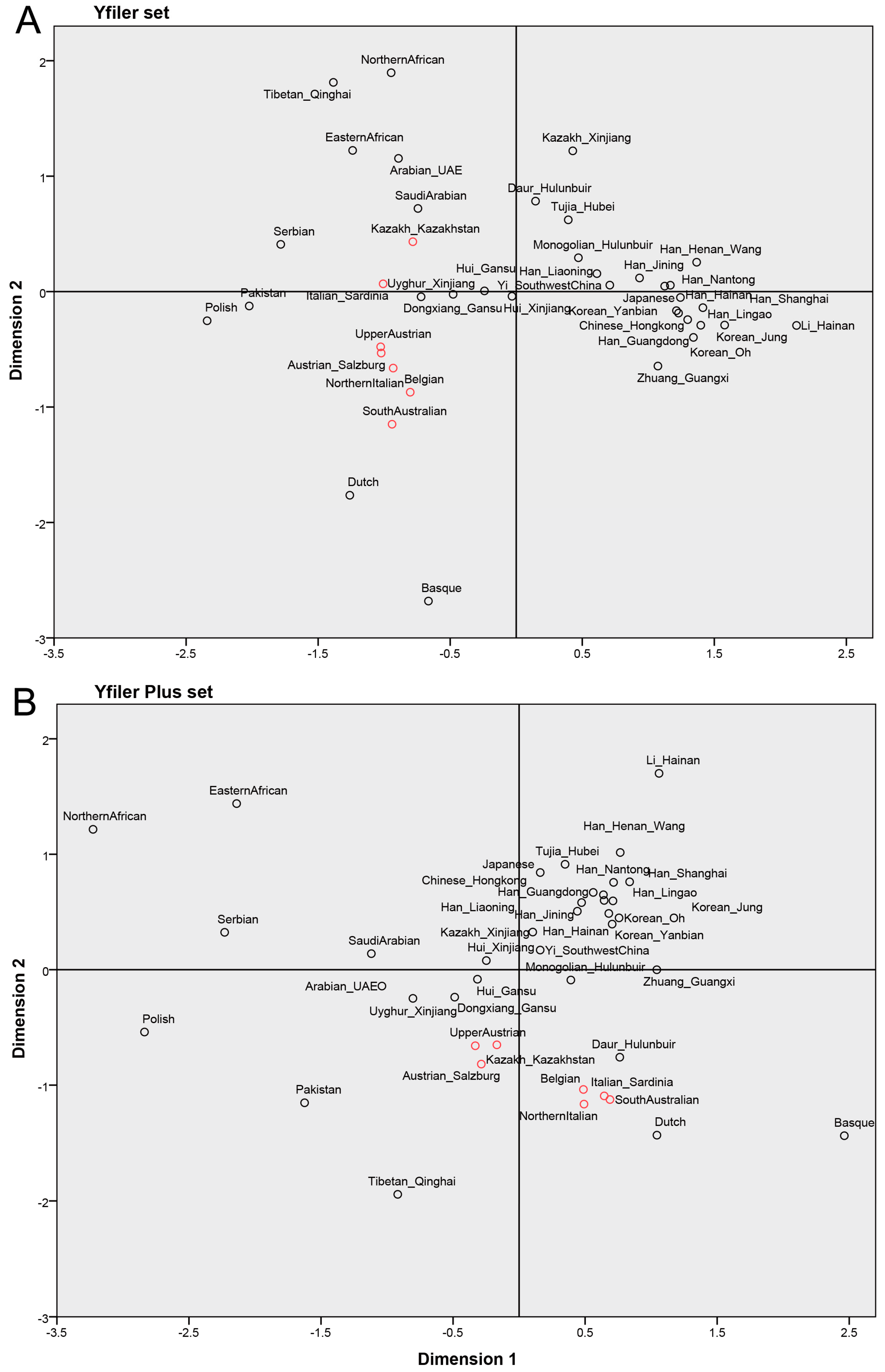
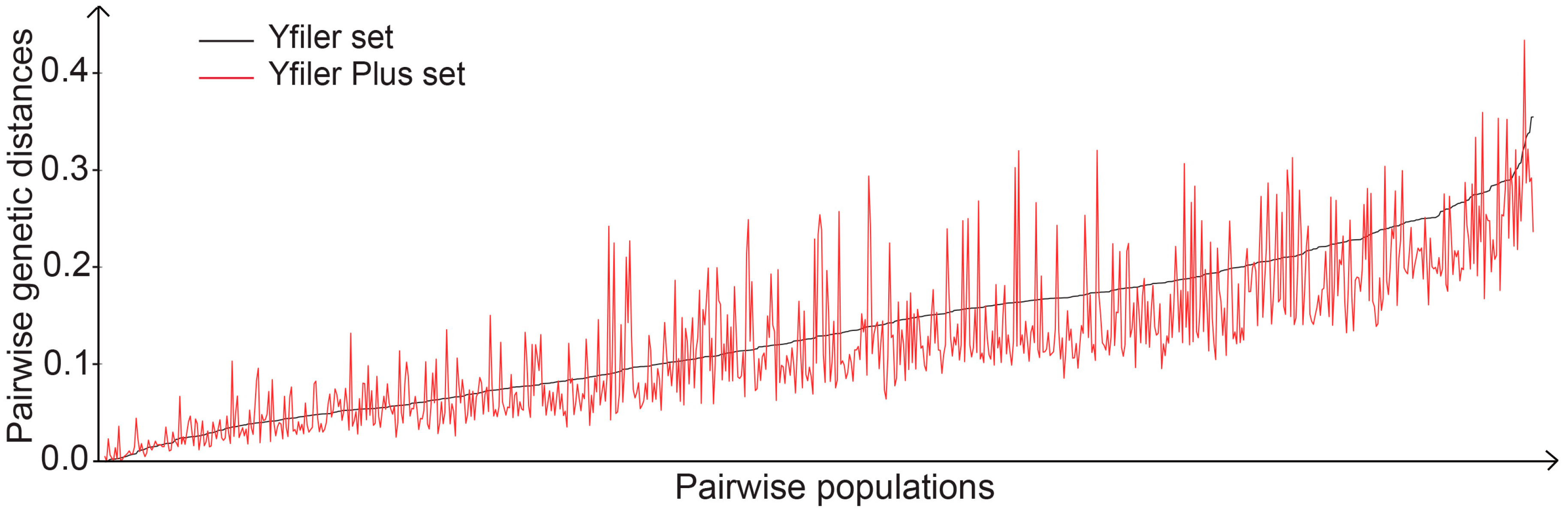
3.1. Evaluation of Population Differentiation Using a Yfiler Set and a Yfiler Plus Set
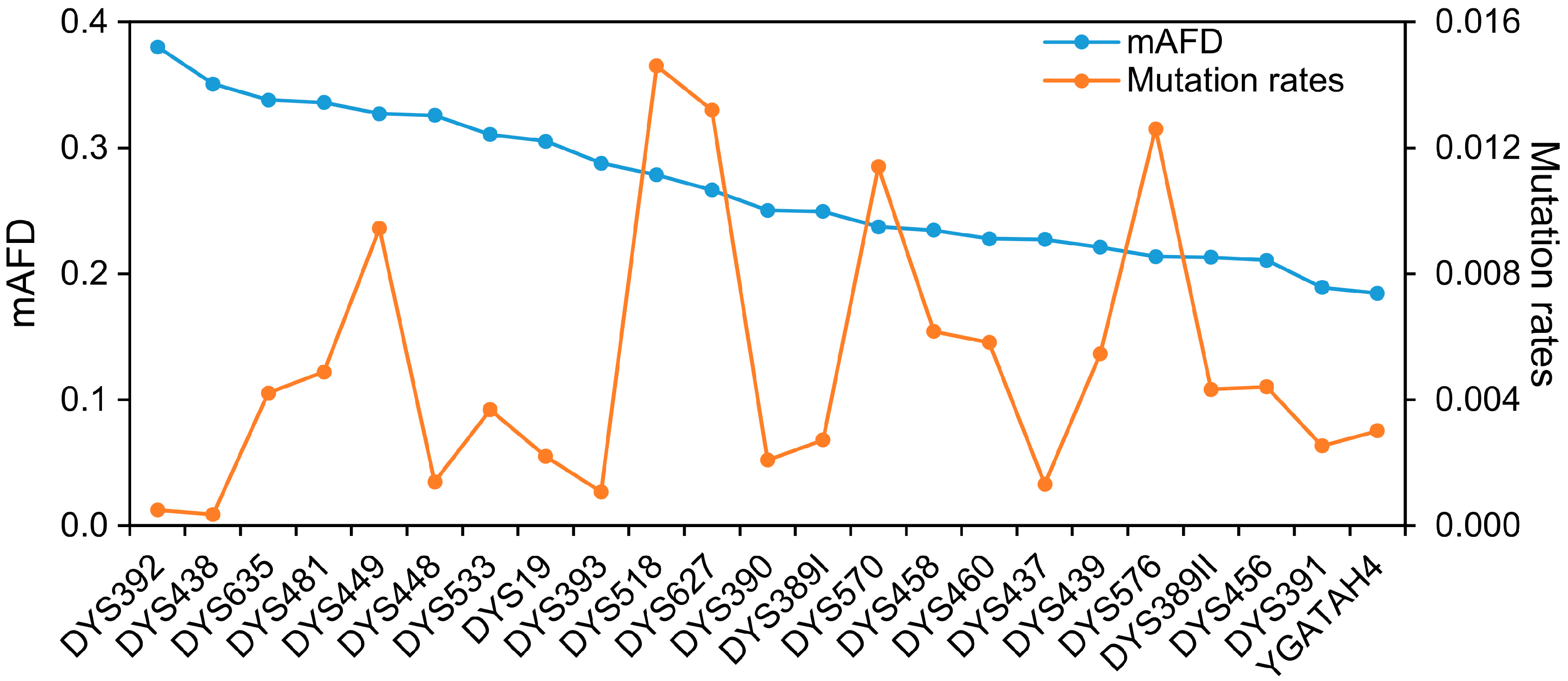
3.2. Assessment of the mAFD Values of Y-STRs
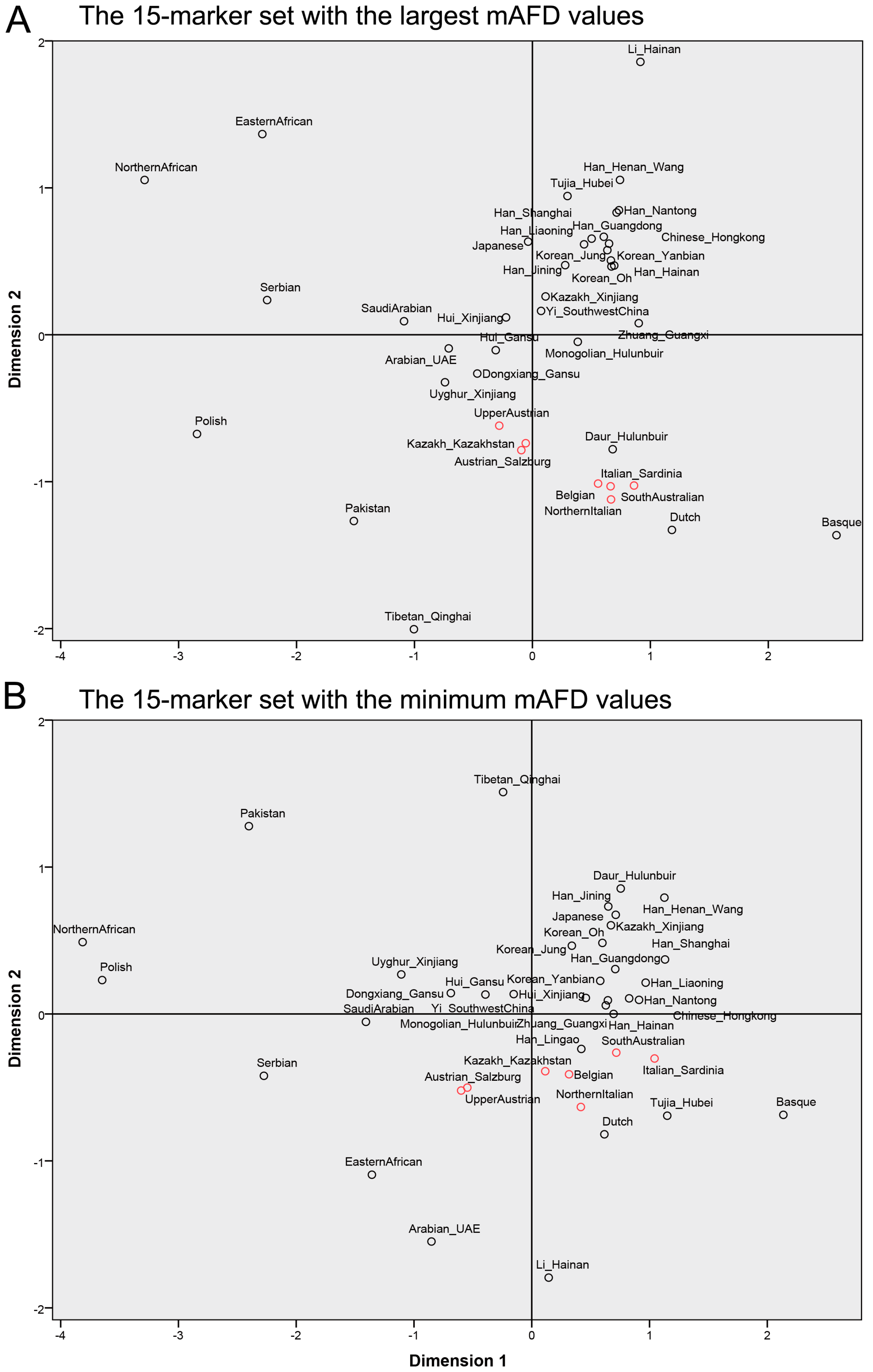
3.3. Analysis of Y-STR Marker Sets in the Population Differentiation
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Sibille, I.; Duverneuil, C.; Lorin de la Grandmaison, G.; Guerrouache, K.; Teissière, F.; Durigon, M.; De Mazancourt, P. Y-STR DNA amplification as biological evidence in sexually assaulted female victims with no cytological detection of spermatozoa. Forensic Sci. Int. 2002, 125, 212–216. [Google Scholar] [CrossRef]
- Prinz, M.; Boll, K.; Baum, H.; Shaler, B. Multiplexing of Y chromosome specific STRs and performance for mixed samples. Forensic Sci. Int. 1997, 85, 209–218. [Google Scholar] [CrossRef]
- Kayser, M. Forensic use of Y-chromosome DNA: A general overview. Hum. Genet. 2017, 136, 621–635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jobling, M.A.; Pandya, A.; Tyler-Smith, C. The Y chromosome in forensic analysis and paternity testing. Int. J. Leg. Med. 1997, 110, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Kayser, M.; Caglià, A.; Corach, D.; Fretwell, N.; Gehrig, C.; Graziosi, G.; Heidorn, F.; Herrmann, S.; Herzog, B.; Hidding, M.; et al. Evaluation of Y-chromosomal STRs: A multicenter study. Int. J. Leg. Med. 1997, 110, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Ballantyne, K.N.; Goedbloed, M.; Fang, R.; Schaap, O.; Lao, O.; Wollstein, A.; Choi, Y.; Van Duijn, K.; Vermeulen, M.; Brauer, S.; et al. Mutability of Y-chromosomal microsatellites: Rates, characteristics, molecular bases, and forensic implications. Am. J. Hum. Genet. 2010, 87, 341–353. [Google Scholar] [CrossRef] [PubMed]
- Gao, T.; Yun, L.; Gu, Y.; He, W.; Wang, Z.; Hou, Y. Phylogenetic analysis and forensic characteristics of 12 populations using 23 Y-STR loci. Forensic Sci. Int. Genet. 2015, 19, 130–133. [Google Scholar] [CrossRef]
- Wang, Z.; Du, W.; He, G.; Liu, J.; Hou, Y. Forensic characteristics and phylogenetic analysis of Hubei Han population in central China using 17 Y-STR loci. Forensic Sci. Int. Genet. 2017, 29, e4–e8. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.; Sarkar, A.; Nandineni, M.R. A comprehensive portrait of Y-STR diversity of Indian populations and comparison with 129 worldwide populations. Sci. Rep. 2018, 8, 2–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, G.L.; Chen, P.Y.; Zou, X.; Chen, X.; Song, F.; Yan, J.; Hou, Y.P. Genetic polymorphism investigation of the Chinese Yi minority using PowerPlex® Y23 STR amplification system. Int. J. Leg. Med. 2017, 131, 663–666. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Mao, J.; Xia, Y.; Bai, X.; Zhu, W.; Peng, D.; Liang, W. Genetic polymorphisms of 17 Y-chromosomal STRs in the Chengdu Han population of China. Int. J. Leg. Med. 2017, 131, 967–968. [Google Scholar] [CrossRef] [PubMed]
- Siegert, S.; Roewer, L.; Nothnagel, M. Shannon’s equivocation for forensic Y-STR marker selection. Forensic Sci. Int. Genet. 2015, 16, 216–225. [Google Scholar] [CrossRef] [PubMed]
- Berner, D. Allele frequency difference AFD—An intuitive alternative to FST for quantifying genetic population differentiation. Genes 2019, 10, 308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Excoffier, L.; Lischer, H.E.L. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef] [PubMed]
- Purps, J.; Siegert, S.; Willuweit, S.; Nagy, M.; Alves, C.; Salazar, R.; Angustia, S.M.T.; Santos, L.H.; Anslinger, K.; Bayer, B.; et al. A global analysis of Y-chromosomal haplotype diversity for 23 STR loci. Forensic Sci. Int. Genet. 2014, 12, 12–23. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Shao, C.; Li, L.; Zhang, Y.; Liu, B.; Yang, Q.; Tang, Q.; Li, S.; Xie, J. Genetic analysis of 29 Y-STR loci in the Chinese Han population from Shanghai. Forensic Sci. Int. Genet. 2018, 32, e1–e4. [Google Scholar] [CrossRef] [PubMed]
- Nei, M.; Tajima, F.; Tateno, Y. Accuracy of Estimated Phylogenetic Trees from Molecular Data. J. Mol. Evol. 1983, 19, 153–170. [Google Scholar] [CrossRef] [PubMed]
- Nei, M. Genetic Distance; Academic Press: Cambridge, MA, USA, 2001; pp. 828–832. [Google Scholar]
- Willuweit, S.; Roewer, L. Y chromosome haplotype reference database (YHRD): Update. Forensic Sci. Int. 2007, 1, 83–87. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Loci | mAFD | Mutation Rates # | Binomial 95%CI | |
|---|---|---|---|---|
| 1 | DYS392 | 0.3802 | 4.96 × 10−4 | 2.14 × 10−4–9.77 × 10−4 |
| 2 | DYS438 | 0.3507 | 3.51 × 10−4 | 0.96 × 10−4–8.99 × 10−4 |
| 3 | DYS635 | 0.3379 | 4.21 × 10−3 | 2.97 × 10−3–5.80 × 10−3 |
| 4 | DYS481 | 0.3359 | 4.88 × 10−3 | 2.52 × 10−3–8.51 × 10−3 |
| 5 | DYS449 | 0.3271 | 9.44 × 10−3 | 5.78 × 10−3–1.45 × 10−2 |
| 6 | DYS448 | 0.3257 | 1.39 × 10−3 | 6.92 × 10−4–2.48 × 10−3 |
| 7 | DYS533 | 0.3104 | 3.68 × 10−3 | 1.68 × 10−3–6.97 × 10−3 |
| 8 | DYS19 | 0.3053 | 2.20 × 10−3 | 1.55 × 10−3–3.04 × 10−3 |
| 9 | DYS393 | 0.2878 | 1.07 × 10−3 | 6.11 × 10−4–1.74 × 10−3 |
| 10 | DYS518 | 0.2786 | 1.46 × 10−2 | 9.86 × 10−3–2.08 × 10−2 |
| 11 | DYS627 | 0.2666 | 1.32 × 10−2 | 8.95 × 10−3–1.88 × 10−2 |
| 12 | DYS390 | 0.2504 | 2.08 × 10−3 | 1.44 × 10−3–2.91 × 10−3 |
| 13 | DYS389I | 0.2494 | 2.72 × 10−3 | 1.96 × 10−3–3.69 × 10−3 |
| 14 | DYS570 | 0.2372 | 1.14 × 10−2 | 7.67 × 10−3–1.62 × 10−2 |
| 15 | DYS458 | 0.2345 | 6.17 × 10−3 | 4.57 × 10−3–8.15 × 10−3 |
| 16 | DYS460 | 0.2278 | 5.82 × 10−3 | 2.80 × 10−3–1.69 × 10−2 |
| 17 | DYS437 | 0.2273 | 1.32 × 10−3 | 7.39 × 10−4–2.18 × 10−3 |
| 18 | DYS439 | 0.2211 | 5.46 × 10−3 | 4.19 × 10−3–6.99 × 10−3 |
| 19 | DYS576 | 0.2136 | 1.26 × 10−2 | 8.86 × 10−3–1.73 × 10−2 |
| 20 | DYS389II | 0.2131 | 4.33 × 10−3 | 3.34 × 10−3–5.51 × 10−3 |
| 21 | DYS456 | 0.2106 | 4.41 × 10−3 | 3.07 × 10−3–6.12 × 10−3 |
| 22 | DYS391 | 0.1891 | 2.53 × 10−3 | 1.82 × 10−3–3.43 × 10−3 |
| 23 | YGATAH4 | 0.1845 | 3.01 × 10−3 | 1.98 × 10−3–4.38 × 10−3 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, Y.; Yao, Y.; Liu, B.; Yang, Q.; Zhou, Z.; Shao, C.; Li, S.; Tang, Q.; Xie, J. Characterizing Y-STRs in the Evaluation of Population Differentiation Using the Mean of Allele Frequency Difference between Populations. Genes 2020, 11, 566. https://doi.org/10.3390/genes11050566
Zhou Y, Yao Y, Liu B, Yang Q, Zhou Z, Shao C, Li S, Tang Q, Xie J. Characterizing Y-STRs in the Evaluation of Population Differentiation Using the Mean of Allele Frequency Difference between Populations. Genes. 2020; 11(5):566. https://doi.org/10.3390/genes11050566
Chicago/Turabian StyleZhou, Yuxiang, Yining Yao, Baonian Liu, Qinrui Yang, Zhihan Zhou, Chengchen Shao, Shilin Li, Qiqun Tang, and Jianhui Xie. 2020. "Characterizing Y-STRs in the Evaluation of Population Differentiation Using the Mean of Allele Frequency Difference between Populations" Genes 11, no. 5: 566. https://doi.org/10.3390/genes11050566
APA StyleZhou, Y., Yao, Y., Liu, B., Yang, Q., Zhou, Z., Shao, C., Li, S., Tang, Q., & Xie, J. (2020). Characterizing Y-STRs in the Evaluation of Population Differentiation Using the Mean of Allele Frequency Difference between Populations. Genes, 11(5), 566. https://doi.org/10.3390/genes11050566